
Abstract
In this study, the anti-inflammatory effects of mangosteen extract (MGE) on dextran sulfate sodium (DSS)–induced colitis in mice and nuclear factor (NF)-κB pathway modulation were investigated. Acute colitis was induced by administering 3% DSS in drinking water for 7 days, and three groups of Institute of Cancer Research mice were treated with 30 and 120 mg/kg MGE or 5-aminosalicylic acid for 7 days; an additional two groups of mice served as healthy and disease controls. The results indicated that MGE significantly prevented weight loss, reduced disease activity index scores, and preserved colon length compared with the findings in the untreated colitis group. MGE downregulated the NF-κB pathway by inhibiting the phosphorylation of IκB and IKK in a dose-dependent manner. These findings suggest that MGE alleviates ulcerative colitis by modulating the NF-κB pathway.
Introduction
I
Nuclear factor-κB (NF-κB) is a key regulator of the inducible expression of numerous genes involved in immune and inflammatory responses in the gut. 5 Moreover, the NF-κB pathway has been defined primarily in response to cytokines that have important roles in the pathogenesis of chronic inflammatory diseases such as gastric ulcer and UC. 6 –8 NF-κB is markedly activated in IBD patients, and through the ability of the transcription factor to promote the expression of various pro-inflammatory genes, NF-κB strongly influences the course of mucosal inflammation. 9 Although sulfasalazine and the 5-aminosalicylic acid (5-ASA) drugs are available as therapeutic agents for IBD and are also known to regulate DNA binding of NF-κB, 10 adverse effects of these drugs such as exanthema, fever, nausea, and vomiting have limited their use for IBD treatment. 11
Garcinia mangostana L., commonly known as mangosteen, is a tropical evergreen tree with leathery glabrous leaves. 12 People in tropical countries have used the pericarp of mangosteen as a traditional medicine to treat abdominal pain, diarrhea, dysentery, infected wounds, suppuration, and chronic ulcer. 13 Mangosteen contains xanthones, tannins, and multiple vitamins as biologically active compounds, 14 and numerous studies have been conducted to evaluate its health benefits such as antiangiogenesis and anti-allergic and anti-obesity effects. 15 –17 Although mangosteen has shown beneficial effects and seen growing applications, its exact mechanisms in treating IBD have not been discovered. Thus the purpose of this study was to determine the role of mangosteen extract (MGE) in IBD using dextran sulfate sodium (DSS)–induced murine colitis; DSS-induced mice is an established animal model for IBD, and the disease progression in mice is similar to that in humans in aspects of inflammation, clinical signs (diarrhea, occult blood, and gross rectal bleeding), histopathological changes of the colon, including prominent sites, and shortening of the colon.
Materials and Methods
Extraction and isolation of test material
Peels of G. mangostana were obtained from a local store in Phuket, Thailand, in June 2012. These plant samples were identified by Dr. Jin Hyub Paik, International Biological Material Research Centre, Korea Research Institute of Bioscience and Biotechnology, Daejeon, Korea. Raw material used in the present study (CYWDU-CP002) was deposited at the College of Pharmacy at Dongguk University, Seoul, Republic of Korea, and was extracted with ethanol followed by evaporation and lyophilization. We conducted quantitative analysis of α- and γ-mangostin using a previously reported method, and 25.7% of α-mangostin and 3.8% of γ-mangostin were present in the MGE. 18
Reagents
DSS and 5-ASA were purchased from Sigma-Aldrich Corp. (St Louis, MO, USA). Polyethylene glycol 400 was procured from Showa Chemical Company (Tokyo, Japan). Phosphorylated IKKα (p-IKKα), IKKα, phosphorylated IκBα (p-IκBα), IκBα, NF-κB (p65), and β-actin antibodies were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
DSS-induced colitis
The protocols for the animal studies were approved by the Institute of Laboratory Animal Resources of Dongguk University, Seoul (IACUC-2016-020-1). Male Institute of Cancer Research mice (6 weeks old, weighing 20–30 g) were purchased from Charles River Company Korea (Orient, Seoul, South Korea). Upon arrival, the animals were randomized and housed three per cage under strictly controlled environmental conditions (20–25°C and 48–52% relative humidity) with a 12-h light/12-h dark cycle. To induce experimental colitis, the mice were divided into five groups as follows: CON (control; n = 8), DSS (n = 8), DM30 (n = 8), DM120 (n = 8), and D5A100 (n = 8). In the DM30 and DM120 groups, 30 and 120 mg/kg, respectively, of MGE (dissolved in distilled water containing 0.5% carboxymethylcellulose [CMC]) were orally administered for 3 days before initiating the DSS treatment. In addition, the D5A100 group was orally administered 100 mg/kg of 5-ASA (dissolved in 0.5% CMC distilled water) in the same period. The distilled water with 0.5% CMC was administered to the CON and DSS groups for 3 days. From the 4th to 10th days, 4% DSS (dissolved in filtered purified water; Millipore Corp., Bedford, MA, USA) was administered for 7 days to the DSS, DM30, DM120, and D5A100 groups. In the same period, the filtered-purified water was provided to the CON group. We observed no differences in water consumption between groups (11.4 ± 2.92 mL/[day·mouse]) throughout the experimental period.
Disease activity index scores in DSS-induced colitis
Mice were examined each day to check for evidence of colitis development by monitoring the body weight, stool consistency, and gross rectal bleeding from day 0 to 10 based on previous reports. 19,20 The overall disease activity index (DAI) scores were the combined scores of body weight changes compared with day 1, diarrhea score, and bleeding score during DSS treatment, all of which were assessed by a clinical scoring system on a scale of 0–4. In brief, scoring was as follows: 0, no weight loss, no occult blood in the stools, and normal stool consistency; 1, weight loss of 1–5%, no occult blood, and normal stool consistency; 2, 5–10% weight loss, positive for fecal occult blood, and loose stools; 3, 10–20% weight loss, positive for fecal occult blood, and loose stools; and 4, greater than 20% weight loss, gross rectal bleeding, and diarrhea.
Dissection
On day 10, the mice were anesthetized, and blood samples were taken for plasma biochemistry analysis. The mice were then euthanized, the abdomens were opened with longitudinal incisions, and the gastrointestinal tracts were exposed. Tissues were then washed in 0.9% NaCl solution, and colon lengths were recorded. Approximately 0.3–0.5 cm of the most proximal (close to the cecum) and the most distal (rectal) colon ends were cut and snap frozen in liquid nitrogen so we could measure myeloperoxidase (MPO) activity using hematoxylin and eosin (H&E) staining.
Histology
For histological analysis, the middle portions of the colons were fixed with 10% neutralized formalin overnight at 4°C and further processed for paraffin embedding; the paraffin-embedded samples were then cut into 5 μm cross sections for H&E staining. We acquired the images using an Olympus CX41 microscope equipped with a ToupCam digital camera.
Determining colonic MPO activity in the esophagus and colons
We measured MPO activity in the colon following procedures that were described previously with some modifications. 21 The esophagus and colon samples were rinsed with cold phosphate-buffered saline, which were finely minced in 0.5% hexadecyltrimethylammonium bromide dissolved in 50 mM potassium phosphate buffer (pH 6.0) and then homogenized with an IKA-T10 basic homogenizer (Wilmington, NC, USA) twice for 20 sec each. The samples were kept on ice at all times. The solutions were centrifuged at 800 g for 20 min at 4°C, and the supernatant (50 μL) was added to 50 μL of 0.5% hexadecyltrimethylammonium bromide dissolved in 50 mM potassium phosphate buffer (pH 6.0) mixed with 100 μL of 0.68 mg/mL o-Dianisidine dihydrochloride and 0.1% hydrogen peroxide (Fisher Scientific, Hanover Park, IL, USA) for 90 min. We measured the changes in absorbance at 450 nm using a spectrophotometer (Molecular Devices, Sunnyvale, CA, USA). The results were expressed as units of MPO activity per mg protein, where one unit of MPO activity was defined as activity that degraded one micromole of hydrogen peroxide per minute at room temperature.
Immunoblot analysis
We assessed protein expression by Western blotting according to standard procedures, and we determined protein concentrations in the colons using the BIO-RAD protein assay reagent. We prepared a 1:1 mix of 20 μg of protein, from the colons, with 2 × sample buffer (20% glycerol, 4% sodium dodecyl sulfate [SDS], 10% 2-mercaptoethanol, 0.05% bromophenol blue, and 1.25 M Tris-buffer of pH 6.8), loaded it on to an 8% or 15% SDS-PAGE gel, and ran it at 150 V for 90 min. The tissue proteins were transferred to an immunoblot polyvinylidene difluoride (PVDF) membrane with a Bio-Rad semidry transfer system. The PVDF membrane was then incubated with primary antibody (dilutions 1:500–1:1000) with 0.1% Tween 20 in Tris-buffered saline overnight. The blots were washed thrice with Tris-buffered saline containing 0.1% Tween 20 and incubated for 1 h with horseradish peroxidase-conjugated secondary anti-IgG antibody (dilution 1:2000–1:20,000). The blots were washed again thrice with 0.1% Tween 20 in Tris-buffered saline, and immunoreactive bands were developed using the chemiluminescent substrate (Amersham Biosciences, Piscataway, NJ, USA).
Statistical analysis
We deemed P < .05 to be statistically significant using Duncan's multiple range test in IBM Statistical Package of Social Sciences (SPSS) statistics, a posteriori analysis of variance (version 23.0; IBM, New York, NY, USA). All data are expressed as mean ± standard deviation.
Results
Effects of MGE on DAI scores, including body weight loss, diarrhea, and bleeding in stool
We used the total DAI scores, which incorporated body weight loss, diarrhea, and bleeding in stools, to assess the therapeutic effects of MGE in the DSS-induced colitis mouse model (Fig. 1). DSS administration was associated with significant weight loss and the appearance of pasty to liquid bloody stool, which was reflected in the gradual increase in the DAI scores, which reached a maximum of 5.25 ± 1.91 at day 10. The differences in body weight loss and the scores for diarrhea and bleeding in stool between the CON and DSS groups first appeared on day 2, but the significant differences between the two groups were observed on day 7–10. In the DM30 and DM120 groups, the significant restoration of lost body weight and the scores of diarrhea and bleeding in stool appeared on days 8, 9, and 10, which reduced the DAI scores.
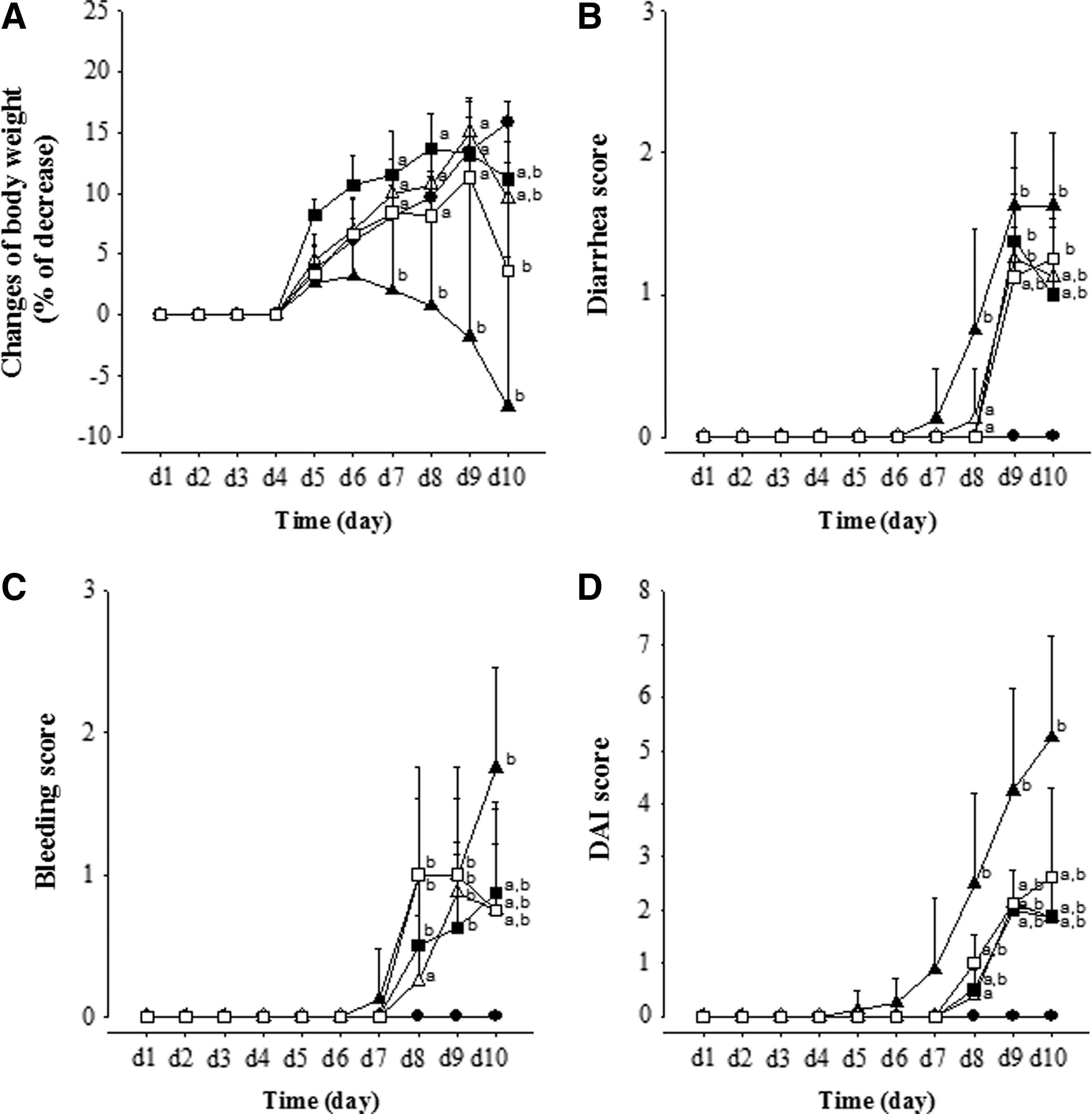
Effects of MGE on the DAI during DSS treatment period. The overall DAI score
Effects of MGE on colon length and macroscopy
The colon lengths, a marker of sufficient induction of colitis by DSS, are shown in Figure 2A. The colons on day 10 in the DSS group were significantly shorter (by 32.4%) than in the CON group, indicating that DSS-induced colitis was sufficiently established for this study. There were no significant changes in the DM30, DM120, and D5A100 groups compared with the DSS group (Fig. 2A). In addition, on macroscopy, we detected different shapes, lengths, ulcerative lesions, and stool contents depending on the DSS-induced colitis conditions with and without therapeutic compounds (Fig. 2B). In this study, the colon condition was significantly restored in DM30 and DM120 groups, compared with DSS group.
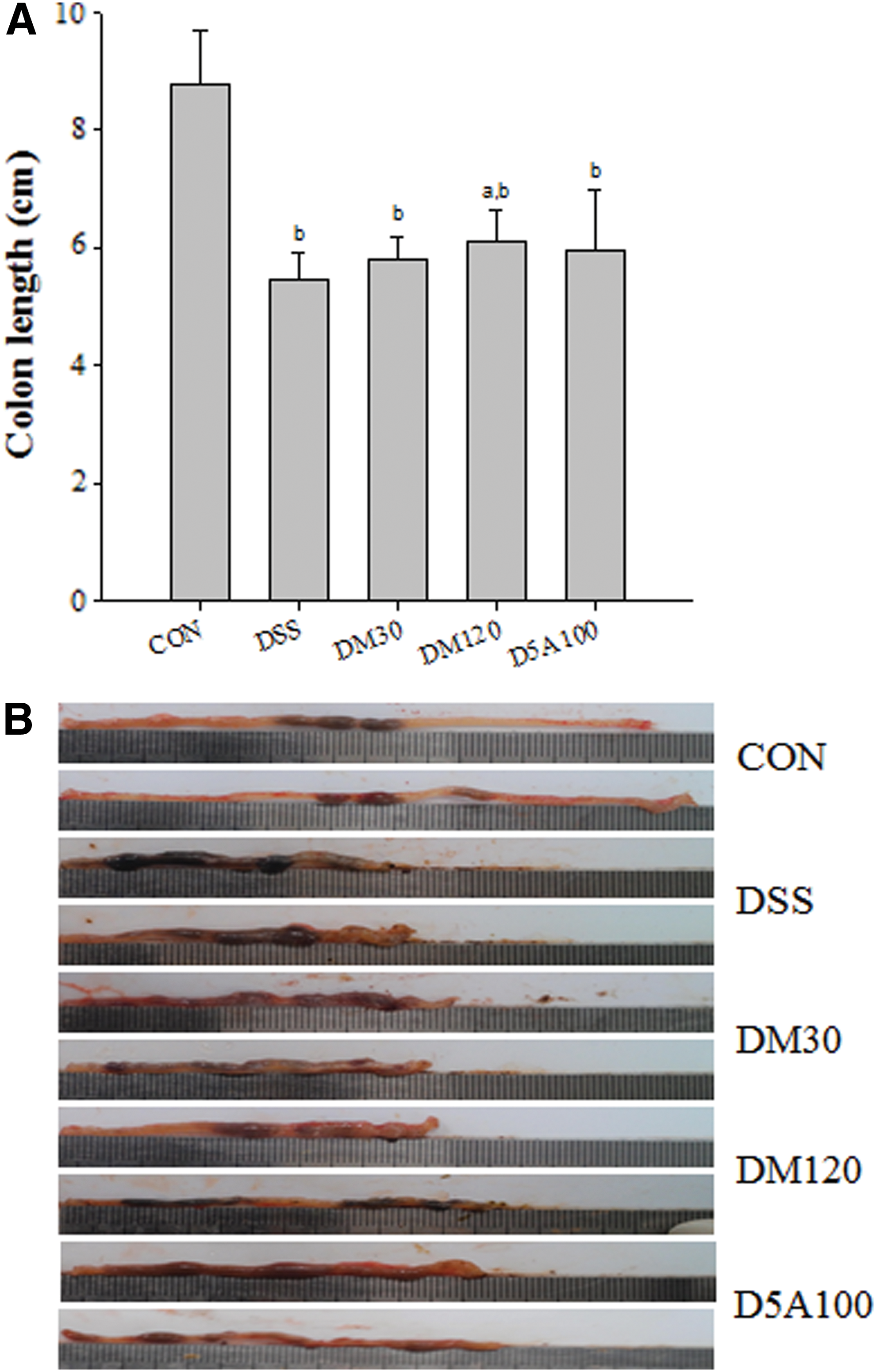
Colon lengths
Histopathological analysis
We conducted histological analysis of colons from each group using H&E staining (Fig. 3). On day 7, all mice were sacrificed, and we monitored the colitis. The control mice had been given water only, and the colons in the water-consuming control group showed no signs of colitis. In contrast, the colons from the group that consumed DSS showed inflammatory infiltrates (Fig. 3B, asterisk), and destroyed epithelial architecture (Fig. 3B, arrow). In the DM30, DM120, and D5A100 groups, the common features of colitis such as the epithelial architecture damage were attenuated.

Effects of DM and D5A on DSS-induced histological injury by H&E staining of colonic tissue sections ( × 100). Eight-week-old Institute of Cancer Research mice were given 5% DSS in drinking water for 7 days with indicated compounds. Control mice were given water only. On day 7, mice were sacrificed, and the histological changes were analyzed. Low- (left panel: 40 × ) and high-magnification (right panel: 100 × ) view of H&E-stained colonic sections. Scale bar represents 150 μm (left panel) and 60 μm (right panel).
In addition, inflammatory cell infiltration and submucosal edema in the DM30 and D5A100 groups were normal. Although inflammatory cell infiltration still appeared in the DM120-treated group, epithelial architecture was protected. The DSS-induced colitis was largely prevented by DM and D5A.
MPO activities in esophagus and colons
MPO activity correlated closely with clinical, macroscopic, and histological grading of inflammation in the experimental groups. MPO activity in the esophagus and colon in the DSS group was significantly higher (by 59.2% and 605%, respectively) than it was in the CON group (Fig. 4). In contrast, MGE treatment at 30 and 120 mg/kg decreased the elevated MPO activity in the esophagus by 56.8% and 40.9%, respectively, compared with the DSS group. The corresponding values in the colons from the DM30 and DM120 groups were decreases of 50.5% and 46.9%, respectively, compared with the DSS group. These results strongly suggest the inhibition of leukocyte recruitment as a mechanism for the protective effects of MGE.

Effects of MGE on MPO activity at day 10 in esophagus
Effects of MGE on the NF-κB pathway activation in colon tissue
We used Western blot analysis to evaluate the effects of MGE on activating the NF-κB pathway in the colon. DSS treatment stimulated NF-κB cascades, including upregulating IKKα and IκBα phosphorylation. To elucidate how NF-κB was affected by MGE, we measured the protein levels of the upstream signaling molecules, such as IKKα and IκBα. We analyzed total colon tissue lysates by Western blot using antibodies to p-IKKα, IKKα, p-IκBα, and IκBα. Figure 5 shows the total IKKα in all groups. NF-κB activation can proceed without the intervention of IκBs. 22 DSS stimulation markedly increased p-IKKα, but MGE treatment downregulated it. As expected from this result, the IκBα protein levels decreased with DSS treatment, and MGE treatment did not restore the IκBα, although it did decrease p-IκBα (Fig. 5). In this study, the NF-κB pathway was activated in the DSS group (vs. CON group), but its NF-κB cascade levels were significantly inactivated in the DM120 and D5A100 group, compared with the DSS group.
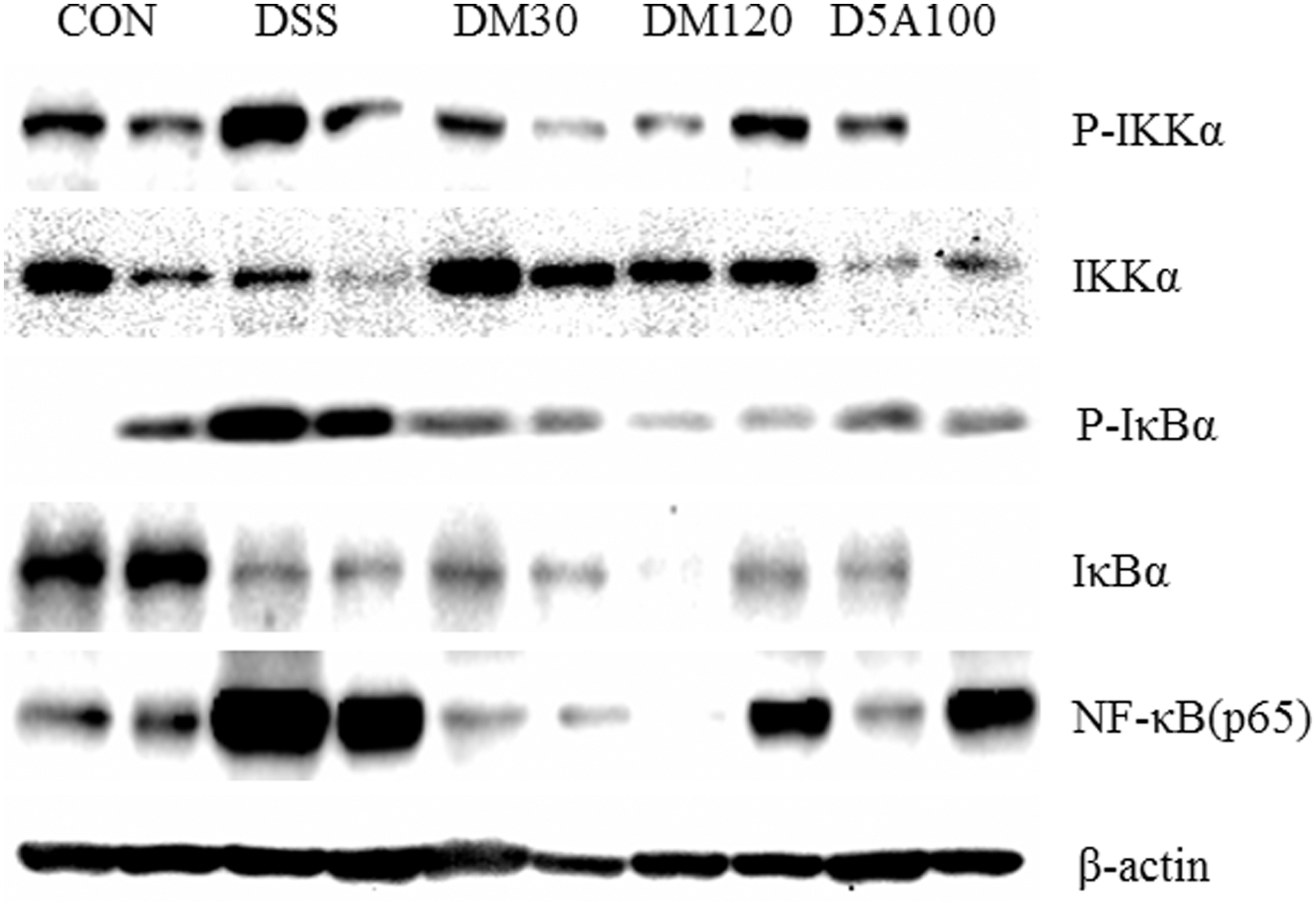
Effects of MGE on IκBα phosphorylation and degradation and phosphorylation of IKK and NF-κB (p65) in DSS-induced colitis. To evaluate the levels of p-IKK, IKK, p-IκBα, IκBα, and NF-κB, the mice colons were treated with DSS for 7 days, after which the colon tissue lysates were analyzed by immunoblot analysis. Immunoblot signals were quantified using Molecular Analyst/PC densitometry software (Version 4.6; Bio-Rad, Hercules, CA, USA). NF, nuclear factor.
Discussion
This study is the first report to clinically and histologically demonstrate that MGE ameliorates DSS-induced colitis in mice, as assessed by DAI, as well as histological inflammatory scores, and that the beneficial effect of MGE treatment might be linked at least, in part, to its inhibition of NF-κB and downregulation of inflammatory mediators such as MPO. These findings suggest that MGE may be a useful therapeutic approach to treating IBD.
Genetics and dietary patterns have been identified as the principal factors for inducing impaired intestinal tract barriers. 23,24 UC typically begins in the rectum and may extend continuously to involve the entire colorectum, with the symptoms of diarrhea, abdominal pain, bloody stools, and weight loss. 25 The DSS model in mice is characterized by both acute and chronic UC and may result from hyperplasia in the epithelial cells and the intestinal mucosal barrier. 26 In the present study, mice with DSS-induced colitis (the DSS group) displayed abnormal physiological conditions such as body weight loss, diarrhea, bloody stools, and shortened colons. To investigate the underlying pathway of MGE in ameliorating the DSS-induced colitis in the mice, we administered 30 and 120 mg/kg of MGE based on our preliminary study (Fig. 1).
Colon shortening is another indicator of the severity of colorectal inflammation. After DSS treatment, the DSS group showed significantly shorter colorectal lengths compared with the vehicle group and the D5A group (Fig. 2). Macroscopically, we observed typical ulcerative lesions in the colons of the DSS group, but not in the control or high-dose DM120 and D5A100 groups (Fig. 2B). The potential protective effects of MGE, as well as 5-ASA, were further confirmed by examining H&E-stained colon slices (Fig. 3). Tissue sections from the control group indicated intact surface epithelia, crypts, muscularis mucosae, and submucosa (Fig. 3). The DSS-induced group exhibited serious inflammatory cell infiltration, large loss of goblet cells and crypts, and extensive submucosal edema, reflected in high DAI scores on histological analysis. However, the colons from the mice in the DM120 and 5-ASA groups, which showed relatively intact histological structures, obtained drastically low DAI scores. Administering a low dose of MGE also moderately diminished the DAI score (Fig. 1D). The MPO expression, as an inflammatory marker, increased significantly during DSS administration. However, MGE- and 5-ASA-treated mice had statistically less MPO activity than did the DSS-induced mice (Fig. 4).
While in the normal state, NF-κB is located in the cytosol complex with the inhibitory protein IκBα, 27 through the intermediacy of integral membrane receptors, a variety of extracellular signals can activate the enzyme IKK. 28 The phosphorylation and subsequent degradation of IκB are important events for NF-κB activation. 29 Therefore, NF-κB may be considered an ideal target for IBD treatment. In this experiment, we observed the phosphorylation of IKK in the DSS group, and the combination treatment with MGE and DSS appeared to decrease IKK phosphorylation compared with the levels in the DSS group. IKK, in turn, phosphorylates IκBα, which results in ubiquitination, dissociation of IκBα from NF-κB, and eventual degradation of IκBα by the proteasome. 30 In addition, IκBα and NF-κB (p65) in the NF-κB pathway were significantly downregulated by MGE and 5-ASA compared with the DSS group (Fig. 5). NF-κB has been commonly observed in inflammatory responses, and in the present study, the elevated NF-κB (p65) in DSS-induced colitis was lowered by 5-ASA or MGE treatment.
Our study explored the therapeutic benefits of mangosteen for UC, and treatment with MGE significantly attenuated DSS-induced experimental colitis in mice. The mechanism of MGE involved inhibiting colorectal NF-κB (p65) activation in mice. Our results collectively suggest that MGE might be an effective candidate for treating UC.
Footnotes
Acknowledgments
This work was supported by a grant from the National Research Foundation of Korea funded by the government of Korea (NRF-2016R1C1B2010849; Y.H.C.) and grants from the GRRC program of Gyeonggi Province ([GRRC-DONGGUK2016-B01], development of new health supplements/therapeutics for neurodegenerative diseases and [GRRCDONGGUK2016-B03], development of functional food to alleviate metabolic syndromes and circulatory disorders).
Author Disclosure Statement
No competing financial interests exist.