
Abstract
Stress contributes to physiological changes such as weight loss and hormonal imbalances. The aim of the present study was to investigate antistress effects of high hydrostatic pressure extract of ginger (HPG) in immobilization-stressed rats. Male Sprague-Dawley rats (n = 24) were divided into three groups as follows: control (C), immobilization stress (2 h daily, for 2 weeks) (S), and immobilization stress (2 h daily, for 2 weeks) plus oral administration of HPG (150 mg/kg body weight/day) (S+G). Immobilization stress reduced the body weight gain and thymus weight by 50.2% and 31.3%, respectively, compared to the control group. The levels of serum aspartate transaminase, alanine transaminase, and corticosterone were significantly higher in the stress group, compared to the control group. Moreover, immobilization stress elevated the mRNA levels of tyrosine hydroxylase (Th), dopamine beta-hydroxylase (Dbh), and cytochrome P450 side-chain cleavage (P450scc), which are related to catecholamine and corticosterone synthesis in the adrenal gland. HPG administration also increased the body weight gain and thymus weight by 12.7% and 16.6%, respectively, compared to the stress group. Furthermore, the mRNA levels of Th, Dbh, phenylethanolamine-N-methyltransferase, and P450scc were elevated by the HPG treatment when compared to the stress group. These results suggest that HPG would have antistress effects partially via the reversal of stress-induced physiological changes and suppression of mRNA expression of genes related to corticosterone and catecholamine synthetic enzymes.
Introduction
S
Ginger (Zingiber officinale Roscoe) has been reported as having therapeutic and preventive effects on inflammation, cancer, obesity, and hypertension for thousands of years. 8 –11 Moreover, ginger ethanol extract alleviates the physiological disturbances of rats exposed to swimming endurance stress or cold stress, 12 and ginger and ginkgo biloba extract reduces anxiety of rats subjected to plus-maze testing. 13 Ginger extracts contain a variety of bioactive constituents, whose major compounds are gingerols and shogaols. 14 Gingerols are the major bioactive component in fresh ginger, whereas shogaols are more plentiful in dried ginger. 14 However, the constitution of ginger extract can vary depending on the extraction conditions, and the maximum effect of ginger function can be changed based on the processing technique.
High hydrostatic pressure (HHP) is a nonthermal food processing technology that can improve shelf-life, function, and appearance of foods. 15 Since HHP uses low temperature (−20 to 60°C), there is little change in heat-sensitive and volatile food components when extracting, compared with other thermal extraction methods. 16 Another advantage of using HHP is that it increases extraction efficiency without destroying the physiologically active substance unlike hydrothermal extraction. 16 Under HHP condition, cell membranes can be disrupted, which leads to the easy elution of cellular compounds into the extracellular space. 17 Total phenolic contents and antioxidant activity of longan fruit pericarp extract prepared by high pressure are higher than conventional extract. 18 HHP extraction increases extraction efficiency of ginsenoside of ginseng compared to heat extraction. 19
Ethanolic extract of ginger increases blood cell counts and organ weights in rats under forced swimming endurance stress or cold stress. 12 However, beneficial effects of high hydrostatic pressure extract of ginger (HPG) on immobilization stress are not clear. Moreover, the underlying mechanism by which ginger extracts exert antistress actions at the molecular level has not yet been fully elucidated. Therefore, the present study was designed to investigate the beneficial effects of HPG on immobilization-stressed rats. The body weight gain and food intake were measured, and the organ weights and serum corticosterone concentrations were analyzed. In particular, to understand the mechanism of stress relieving of HPG, mRNA expression of genes involved in corticosterone and catecholamine biosynthesis was examined in the adrenal gland.
Materials and Methods
Preparation of materials
Ginger extract was kindly supplied by the Korea Food Research Institute (KFRI) (Seongnam, Gyeonggi, Korea). Ginger was purchased from the local market of Muan (Muan-gun, Jeollanam-do, Korea) and used for the preparation of HPG. Ginger suspension was poured into plastic bags with 2% complex enzyme (Termamyl, Celluclast, and Viscozyme) and inserted into a programmable high-pressure treatment apparatus (TFS-10 L; Innoway Co., Bucheon, Korea) set at a pressure of 100 MPa for 24 h at 50°C. After incubation, the extract was stored at 4°C for 24 h for enzyme inactivation and subsequently centrifuged at 11,000 g for 10 min. The supernatant was filtered using Whatman No. 4 filter paper and lyophilized into a powder form.
Determination of 6-gingerol, 6-shogaol, and total saponin
The levels of 6-gingerol and 6-shogaol in ginger extract were analyzed by high-performance liquid chromatography (HPLC) using a JASCO HPLC system (Tokyo, Japan). Standard 6-gingerol and 6-shogaol were purchased from Sigma-Aldrich (St. Louis, MO, USA). Briefly, ginger extract was dissolved in 70% ethanol in 20 mg/mL concentration. An Eclipse XDB column (C18; 4.6 × 250 mm, 10 μm pore size; Agilent Technologies, Palo Alto, CA, USA) was used for chromatographic analysis at a flow rate of 1 mL/min. The mobile phases consisted of solvent A (water) and solvent B (acetonitrile). The elution had the following gradient: 0–12 min, 65% B; 12–27 min, 80% B; and 25–30 min, 45% B. The HPLC chromatogram of the 6-gingerol and 6-shogaol is presented in Figure 1. The contents of 6-ginerol and 6-shogaol in ginger extract were 3.07 ± 0.09 mg/g and 0.65 ± 0.02 mg/g, respectively.
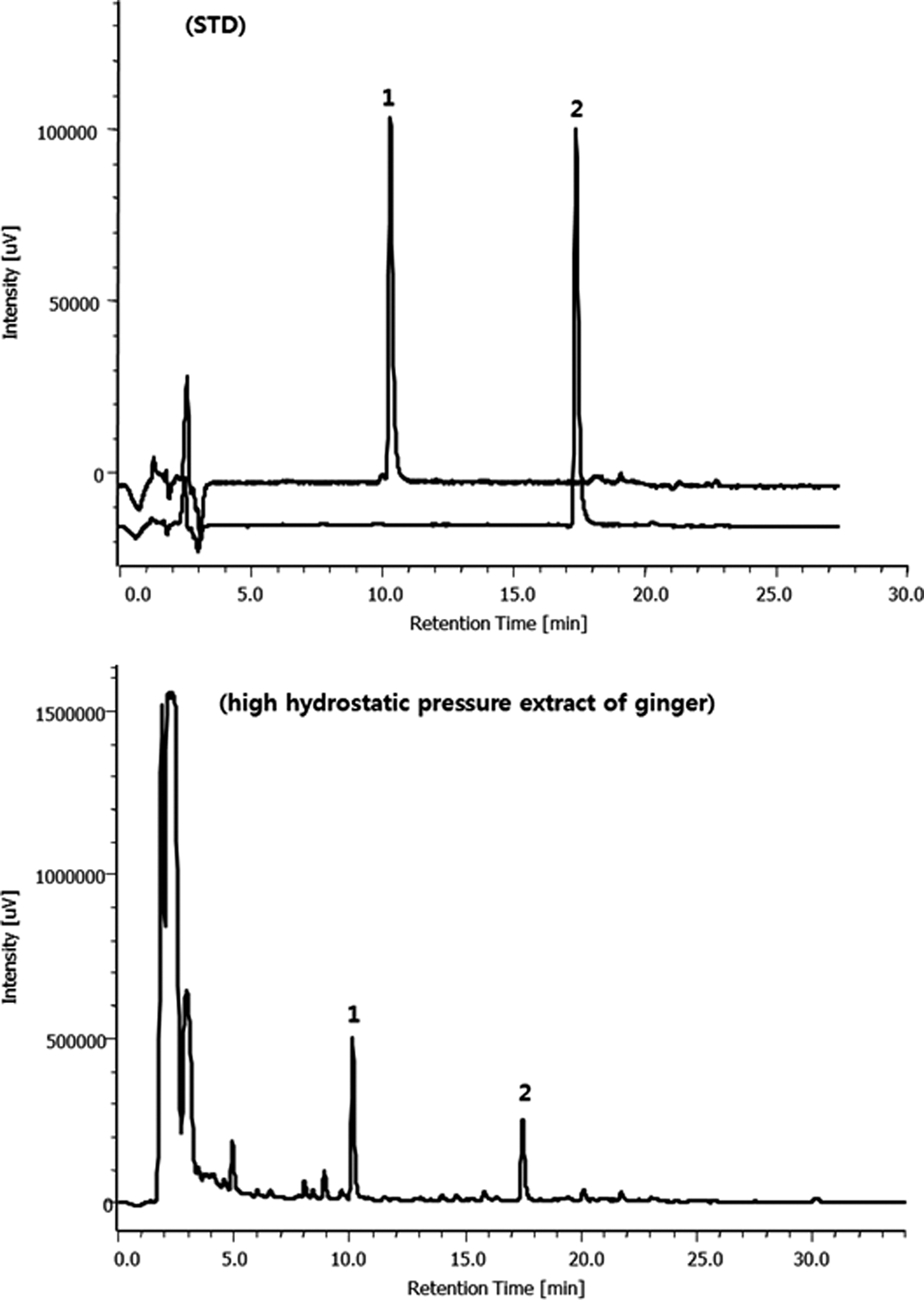
High-performance liquid chromatography chromatogram of 6-gingerol and 6-shogaol in high hydrostatic pressure extract of ginger. 1, 6-gingerol; 2, 6-shogaol.
Total saponin content of ginger extract was determined as follows: the ginger extract was freeze dried and dissolved in 10 mL of distilled water, and 100 μL of solution was mixed with 0.3 μL of 8% vanillin solution and added 4 mL of 75% sulfuric acid, and then shaken and placed in cold water. The tubes are maintained for 10 min at 60°C and then cooled. The average absorbance was measured at 545 nm. Total saponin was quantified by means of calibration of curves obtained from commercial standards of ginsenoside Re (Wako Chem. Co., Japan). The measured saponin content of the ginger extract was 32.89 ± 4.65 g/100 g.
Animals and treatment
Thirty male Sprague-Dawley rats, 6 weeks old (initial weights 150–160 g), were obtained from Doo Yeol Biotech (Seoul, Korea). Each rat was individually housed in a stainless steel wire mesh cage with controlled temperature (22°C ± 2°C), humidity (55% ± 5%), and lighting (12 h light/dark cycle). After 1 week of acclimatization with free access to water and normal chow diet (Harlan 2018S, Indianapolis, IN, USA), the animals were randomly divided into three groups (n = 8 rats per group): control (C), immobilization stress and 10 mL/kg of distilled water (p.o.) (S), and immobilization stress and 150 mg/kg of HPG (p.o.) (S+G). Distilled water and HPG were administered to rats 1 h before immobilization. For immobilization stress experiments, rats were immobilized in restraint cages for 2 h (09:00–11:00 am). Repeated stress was achieved by immobilizing the animals for two consecutive weeks. The Institutional Animal Care and Use Committee (IACUC) of Ewha Womans University approved all animal procedures (IACUC No.15–067). At the end of the experiment, rats were deprived of food for 12 h. After the last immobilization stress, rats were anesthetized using a mixture of 2,2,2-tribromoethanol (Avertin; Sigma, St. Louis, MO) and medetomidine (Domitor®; Orion Corporation, Espoo, Finland) by i.p. injection. Blood samples were collected by cardiac puncture and centrifuged at 1500 g for 20 min at 4°C, and the serum was stored at −70°C until analyzed. The adrenal glands, thymus, spleen, liver, and epididymal adipose tissue were dissected and stored at −70°C for analysis.
Serum analysis
The serum total cholesterol concentration was measured enzymatically using a commercial kit (Asan Pharmaceutical, Seoul, Korea). Aspartate transaminase (AST) and alanine transaminase (ALT) were analyzed using a commercial kit (M.B.L, Gunpo, Korea). Serum corticosterone concentrations were detected using competitive enzyme-linked immunosorbent assay (Enzo Life Sciences, NY, USA). All samples were diluted 1:50 with assay buffer provided in the kit. The sensitivity of the kit was 26.99 pg/mL.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from adrenal glands using TRIzol® reagent (GeneAll Biotechnology, Seoul, Korea). cDNA was synthesized from 4 μg of RNA with the Moloney Murine Leukemia Virus Reverse Transcriptase kit (Bioneer Co., Daejeon, Korea). After cDNA synthesis, real-time polymerase chain reaction (PCR) was performed using AccuPower 2X Greenstar qPCR MasterMix (-ROX Dye) (Bioneer Co.) on a Rotor-Gene 3000 (Corbett Research, Sydney, Australia). Primers used for real-time PCR are presented in Table 1. Gene expression was calculated according to the comparative 2−ΔΔCT method. 20 The ΔΔCt value for each sample was determined by calculating the difference between the Ct value of the target gene and the Ct value of the reference gene (glyceraldehyde 3-phosphate dehydrogenase [Gapdh]). Values are expressed as fold change of the control group.
PCR efficiency (E) was calculated according to the equation E = (10[−1/slope]–1) × 100.
Dbh, dopamine beta-hydroxylase; Gapdh, glyceraldehyde 3-phosphate dehydrogenase; P450scc, cytochrome P450 side-chain cleavage; PCR, polymerase chain reaction; Pnmt, phenylethanolamine-N-methyltransferase; Th, tyrosine hydroxylase.
Statistical analyses
All data are expressed as the mean ± standard error of the mean for eight rats in each group. Statistical analyses were conducted using SPSS software (SPSS, Inc., Chicago, IL, USA) version 19. Significant differences between groups (C and S or S and S+G) were analyzed using an unpaired Student's two-tailed t-test. The P value < .05 was taken to indicate a significant difference.
Results
Body weight gain and food intake
At the end of the experiment, immobilization stress significantly decreased the body weight gain by 50.2% (P < .01) (Fig. 2A) and food intake by 18.1% (P < .01) (Fig. 2B), compared to the control group. Administration of HPG to immobilization-stressed rats significantly increased the body weight gain by 12.7% (P < .05), whereas there were no significant differences in food intake.
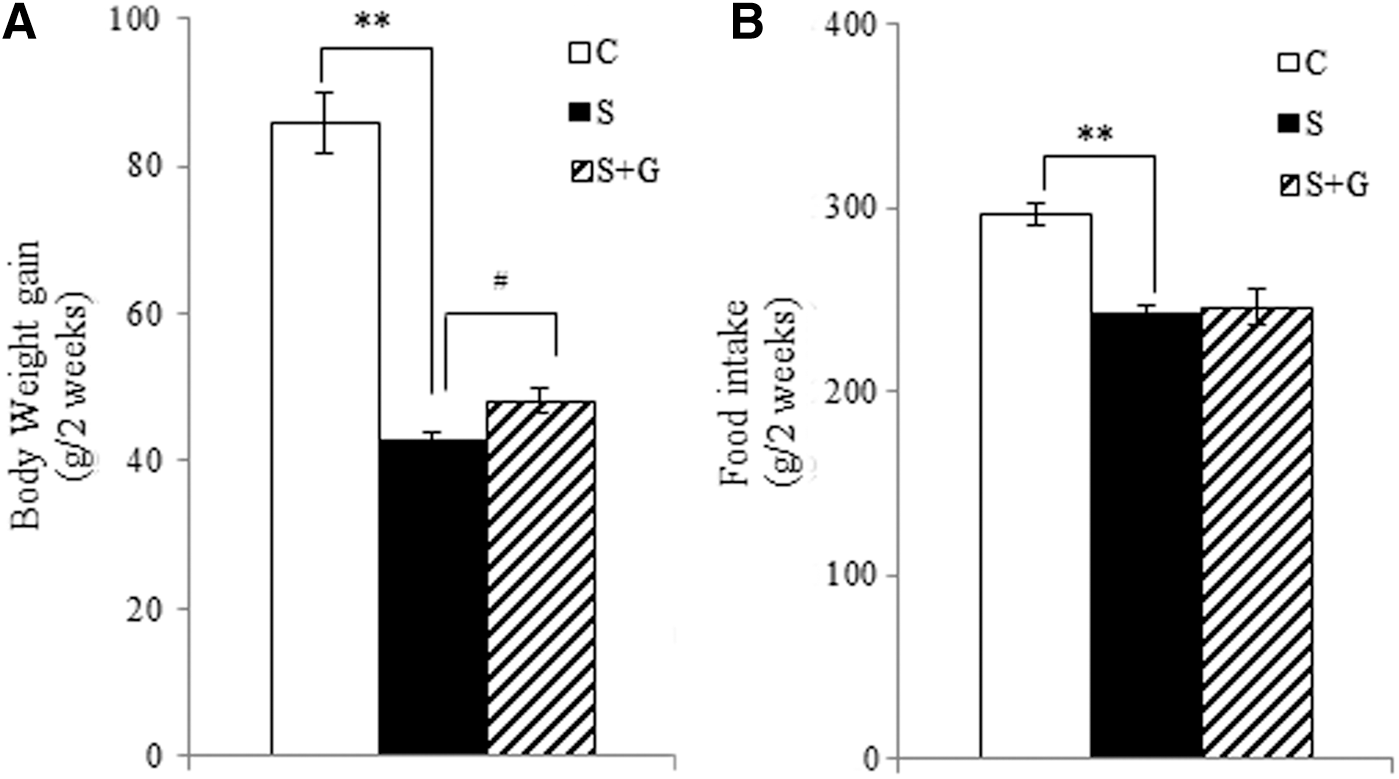
Effects of ginger extract on body weight gain
Tissue weights
Immobilization stress increased the adrenal gland weight (P < .01) and decreased the spleen (P < .05), thymus (P < .01), and epididymal adipose tissue weights (P < .01), compared to those of the control group. Administration of HPG significantly increased the thymus weight by 16.6%, compared to the immobilization stress group (P < .01). However, there were no significant differences in the adrenal gland, spleen, and epididymal adipose tissue weights between the S and S + G groups (Fig. 3a). Liver weight was not statistically different among the groups. Changes in tissue appearance are presented in Figure 3b.
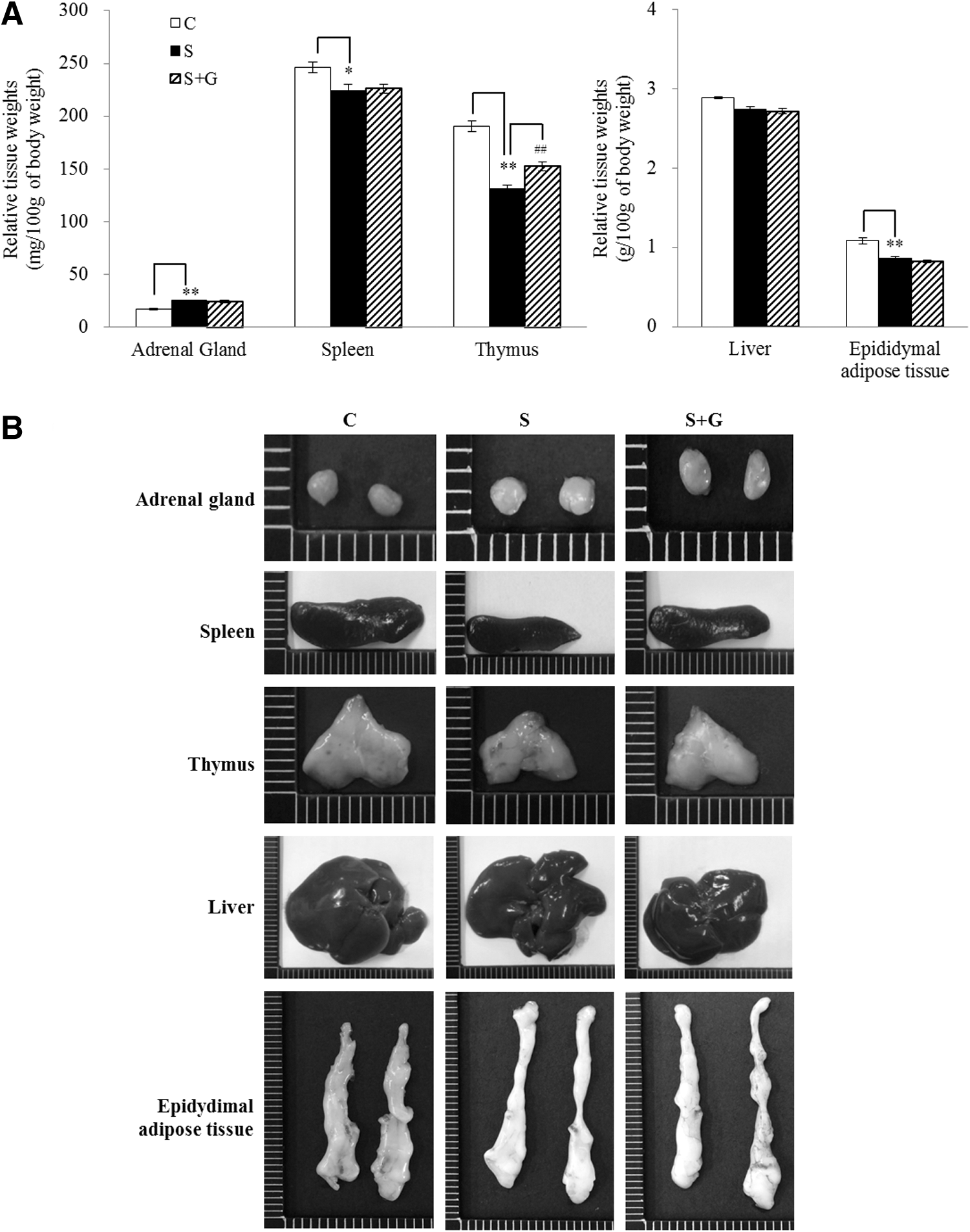
Effects of ginger extract on relative tissue weights
Serum biochemical parameters
Immobilization stress significantly increased the serum AST and ALT levels in comparison to the control group. Administration of HPG to the stressed rats decreased the AST (P < .01) and ALT (P < .05) levels, compared to the immobilization stress group. There were no significant differences in serum total cholesterol concentration among the groups (Table 2). The serum corticosterone concentration was significantly elevated in the S group by 18.2%, compared to the control group (P < .05), whereas the HPG reduced the serum corticosterone level by 22.6%, compared to the immobilization stress group (P < .05, Fig. 4).
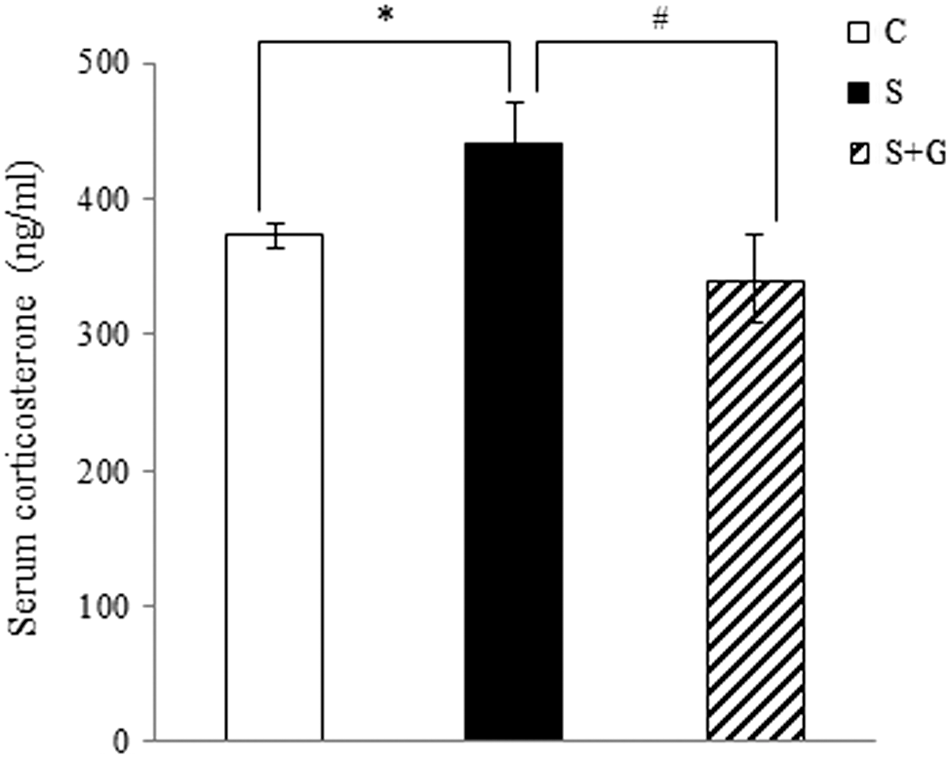
Effect of ginger extract on serum corticosterone concentrations of immobilization-stressed rats. The data are expressed as mean ± SEM (n = 8). Differences in mean values between the corresponding C and S groups were assessed by unpaired t-tests: *P < .05, differences in mean values between the corresponding S and S + G group were assessed by unpaired t-tests: # P < .05. C, control; S, immobilization stress; S+G, immobilization stress plus high hydrostatic pressure extract of ginger.
The data are expressed as mean ± standard error of the mean (n = 8). Differences in mean values between the corresponding C and S groups were assessed by unpaired t-tests: * P < .05 and ** P < .01, differences in mean values between the corresponding S and S + G group were assessed by unpaired t-tests: # P < .05 and ## P < .01.
ALT, alanine transaminase; AST, aspartate transaminase; C, control; S, immobilization stress; S+G, immobilization stress plus high hydrostatic pressure extract of ginger.
Gene expression in the adrenal glands
To elucidate the molecular mechanism by which the HPG decreased the effects of immobilization stress in rats, we examined mRNA expressions in the adrenal glands of rats. The mRNA levels of genes related to catecholamine synthesis such as tyrosine hydroxylase (Th), dopamine beta-hydroxylase (Dbh), phenylethanolamine-N-methyltransferase (Pnmt) were measured. In addition, the mRNA level of P450scc (cytochrome P450 side-chain cleavage or CYP11A1), a corticosterone synthetic enzyme, was also analyzed. In the immobilization stress group, the Th (P < .05) and Dbh (P < .01) mRNA levels were significantly increased by 3.7- and 4.2-fold, compared to the control group, respectively (Fig. 5). The Pnmt mRNA expression level in the S group was partially increased, but the change was not statistically significant. HPG significantly decreased the Th (P < .05), Dbh (P < .01), and Pnmt (P < .01) mRNA levels by 70.2%, 78.1%, and 70.1%, respectively, compared to the S group (Fig. 5). Furthermore, the mRNA level of P450scc in the S group was significantly increased by 3.04-fold, compared to the control group (P < .05, Fig. 5). However, HPG markedly downregulated the P450scc mRNA expression by 91.8%, compared to the S group (P < .05).
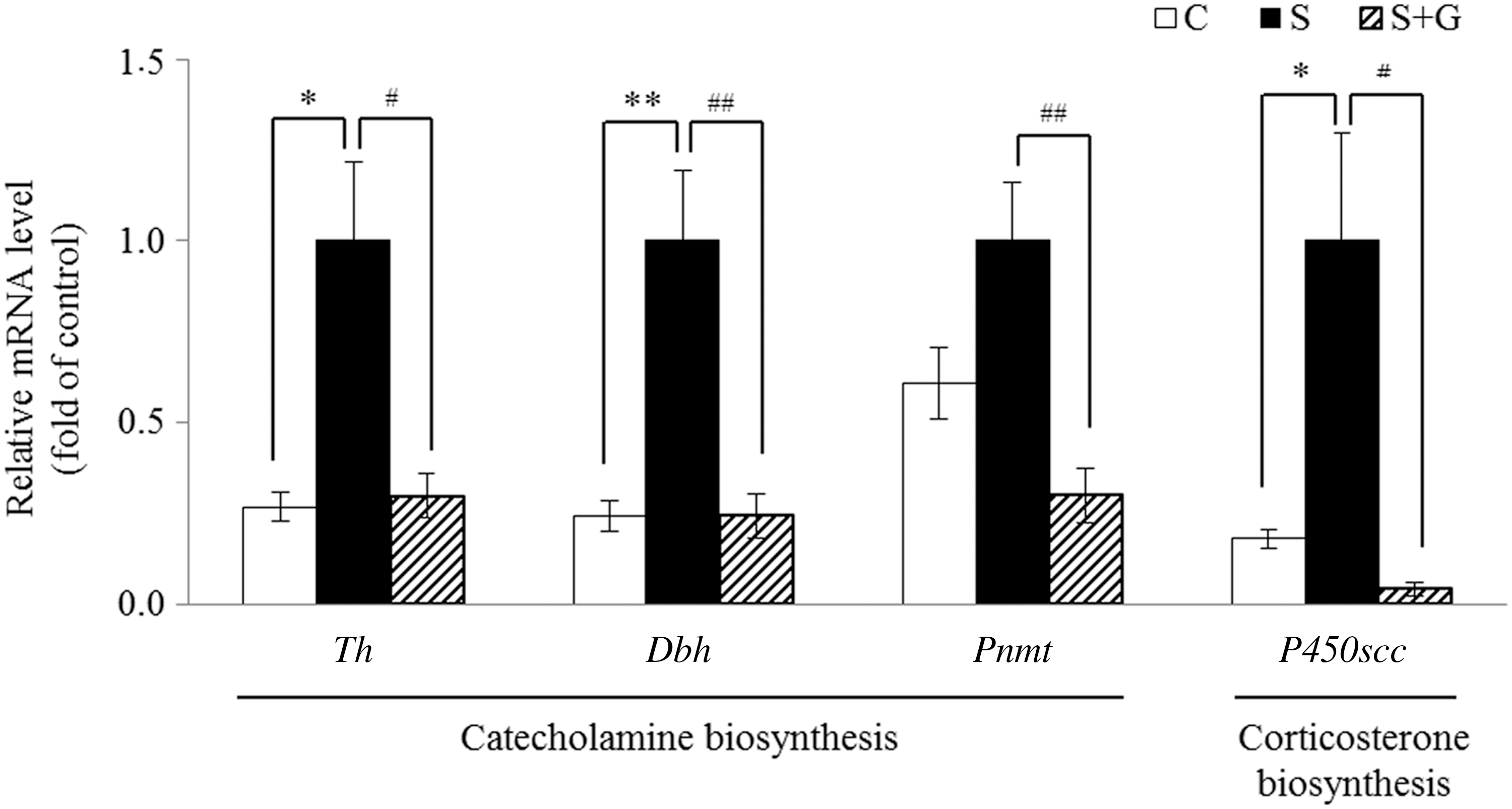
Effect of ginger extract on relative mRNA levels of genes related to catecholamine and corticosterone biosynthesis in adrenal glands of immobilization-stressed rats. The mRNA levels were analyzed by quantitative real-time polymerase chain reaction and normalized to glyceraldehyde 3-phosphate dehydrogenase. The data are expressed as mean ± SEM (n = 8). Differences in mean values between the corresponding C and S groups were assessed by unpaired t-tests: *P < .05 and **P < .01, differences in mean values between the corresponding S and S + G group were assessed by unpaired t-tests: # P < .05 and ## P < .01. C, control; Dbh, dopamine beta-hydroxylase; P450scc, cytochrome P450 side-chain cleavage; Pnmt, phenylethanolamine-N-methyltransferase; S, immobilization stress; S+G, immobilization stress plus high hydrostatic pressure extract of ginger; Th, tyrosine hydroxylase.
Discussion
Chronic stress influences homeostasis of metabolic and neural states. 21 Repeated exposure to stress decreases the body weight and food intake, induces adrenal hypertrophy, and adversely affects the function of spleen, thymus, and lymph nodes. 22 The positive effects of ethanolic ginger extract on the attenuation of swimming endurance or cold stress-induced perturbation have been reported. 12 Thus, the aim of this study was to examine the beneficial effects of HPG on immobilization-induced stress in rats. In addition, mRNA expression of genes related to catecholamine and corticosterone synthesis was analyzed to investigate the molecular mechanism involved in the antistress effect of ginger extract.
To investigate the effect of HPG on physiological changes in immobilization-stressed rats, body weight and food intake were measured. Repeated stress induces a chronic weight loss independent of energy status. 23 Weight loss in animals under chronic stress is due to increased metabolic rate as well as decreased dietary intake. It is well established that corticotropin-releasing hormone (CRH) is elevated in the brain in response to stress, and CRH blocks the neuropeptide Y (NPY) release that stimulates food intake and suppresses sympathetic nerve activity to brown adipose tissue. 24,25 Sympathetic nerve activation increases body surface temperature that could lead to increased energy expenditure and weight loss. Rats exposed to restraint stress showed body weight reduction and heat loss due either to vasodilation or increased heat production. 24 Furthermore, stress hormones such as catecholamine and glucocorticoid have been reported to facilitate catabolic reactions. Catecholamine stimulates lipolysis in adipose tissue in vitro. 26 Glucocorticoid inhibits glucose and amino acid uptake, and enhances lipolysis in adipose tissue. 27
In this study, the body weight gain and food intake in stressed rats were reduced compared to the controls. These observations correspond to animal studies in which chronic stress causes weight loss and anorexic effects in male rats. 28 Our study also showed that HPG increased the body weight gain compared to the stressed rats, but did not change the food intake. Similarly, it has been reported that Polygala tenuifolia is effective in ameliorating weight loss in mice exposed to repeated restraint stress for 14 days. 29 Also, ginseng extract was effective in ameliorating weight loss in immobilization-stressed rats. 30 Therefore, it can be postulated that HPG inhibits weight loss induced by immobilization stress, and that inhibition of sympathetic nerve activity and suppression of catabolic response seem to be partially associated with the effects of HPG.
Chronic stress enlarges the adrenal gland by releasing the corticosterone from the adrenal cortex continuously. 2 The spleen is the primary site for the storage of lymphocytes, and in a stressful states, many splenic lymphocytes are released into the blood, resulting in spleen atrophy. 3,31 The thymus plays a central role in the production and secretion of T cells, and chronic stress causes apoptosis and necrosis of immature T cells and B cells, thereby reducing the thymus weight. 32,33 Studies have reported that functional food materials could improve stress-induced immune dysfunction. In mice, chlorella vulgaris culture supernatant reduces stress-induced apoptosis in thymocytes, especially the CD4+CD8+ population. 34 Green Brazilian propolis counteracts the alterations found in the spleen of stressed mice. 35 In this study, immobilization stress reduced the spleen and thymus weights, whereas HPG significantly relieved stress-induced thymus atrophy. Thus, it is anticipated that HPG may have the ability to restore the alterations in lymphoid organs due to stress. And new research could be performed to reveal the effects of HPG on immune regulation, particularly T cell differentiation/ function and changes in T cell subtypes, during stress.
AST and ALT are enzymes released into the blood when liver cells are damaged. 36 The reference ranges for AST and ALT of rats are 50–150 and 10–40 IU/L, respectively. 37 However, it has been reported that higher levels of AST and ALT are positively associated with increased mortality from liver disease, even within the normal range. 38 Furthermore, rats administered orally with insecticidal plant extracts showed an increase in leukocyte infiltration and hemorrhage in the liver, even though the levels of AST and ALT were in the normal range. 37 Elevated catecholamine levels in the stressed state result in natural killer T cell development, increased production of interleukin-6 and tumor necrosis factor-α, and hepatic blood flow disorders, leading to liver injury. 39 A previous study has shown that injection of catecholamine-based chemical, norepinephrine, increased AST and ALT levels in serum of dogs. 40 In this study, the levels of AST and ALT in serum were within the normal range. However, the serum AST and ALT levels in immobilization-stressed rats were markedly increased, but the HPG significantly reduced the stress-induced elevation of AST and ALT levels. It has been reported that ginger decreases serum AST and ALT levels in CCI4-induced hepatotoxic rats. 41 In addition, ethanol and aqueous extract of ginger ameliorate liquor- and mancozeb fungicide-induced liver injury, respectively. 42,43 Therefore, it is postulated that HPG would be beneficial in stress condition partially by protecting against stress-induced liver damage.
When the body is stressed, corticosterone is released in a large amount from the adrenal gland by activation of the hypothalamic–pituitary–adrenal axis, which is one of the primary defense mechanisms for external stressors. In our experiment, the serum corticosterone level in immobilization-stressed rats was increased compared to the control group, and HPG significantly reduced the corticosterone production in stressed rats. This may be because the HPG modifies the pathways that would respond to immobilization stress. This assumption can be supported by a study that the serum corticosterone level was decreased by an ethanolic ginger extract in immobilization-stressed rats. 12 Moreover, this implies that consumption of ginger may be able to favorably regulate the secretion of stress hormones.
To determine whether immobilization stress affects adrenal gene expression in rats, mRNA levels of genes related to catecholamine and corticosterone synthesis were examined. Catecholamine is produced in respond to stress, and includes epinephrine, norepinephrine, and dopamine. TH is an enzyme catalyzing tyrosine to dihydroxyphenylalanine (DOPA), which is the initial rate-limiting step in the biosynthesis of catecholamine. DOPA is converted to dopamine by DOPA decarboxylase, which in turn is converted to norepinephrine by the addition of a hydroxyl group to the dopamine by DBH. The norepinephrine is converted to epinephrine by PNMT. Under the stressed state, elevated norepinephrine infusion into the paraventricular nucleus dose dependently increases the circulating level of corticosterone. 44 The transfer of cholesterol to cytochrome P450 is the primary rate-limiting step in corticosterone synthesis in the adrenal gland. This reaction is catalyzed by mitochondrial membrane-bound enzyme P450scc. 45
Immobilization stress triggers catecholaminergic action by promoting Th, Dbh, and Pnmt mRNA expressions in adrenal glands of rats. 46 In addition, mice exposed to restraint stress also showed increased mRNA expression of Th, Dbh and Pnmt, while their protein levels were not affected. 47 The P450scc mRNA and protein levels also tended to be increase in goats exposed to stress by subacute ruminal acidosis. 48 Our study showed that the mRNA expression of Th, Dbh, and P450scc increased in the adrenal glands of immobilization-stressed rats. However, HPG decreased the mRNA levels of Th, Dbh, Pnmt, and P450scc, compared to the stressed rat group. Similarly, ginseng extract has antistress effects by normalizing elevated serum corticosterone concentration and the levels of Th and Dbh mRNA by stress. 22 Moreover, the aqueous ginger extract increases TH and DBH activity in chronic ethanol-fed rats, which suggests ginger extract influences the levels of monoamine neurotransmitters. 49 Thus, it is postulated that HPG would modulate the expression of genes involved in catecholamine and corticosterone synthesis, which is responsible at least, in part, for the antistress effect of the HPG.
The major physiologically active constituents of ginger are 6-gingerol and 6-shogaol. 6-gingerol and 6-Shogaol act as PPARδ-specific ligands, stimulating fat utilization in the muscle and liver of diet-induced obese mice. 10 Furthermore, 6-gingerol ameliorates inflammation by blocking NF-κB and PKC signaling pathways. 50 6-Gingerol inhibits the catalytic activity of cytochrome P450 enzyme isoforms such as CYP1A2 and CYP2C19 in human liver microsomes. 51 In this study, 6-gingerol and 6-shogaol were the major components of HPG, and thus, we assumed that these components may be associated with a reversal in gene expression related to catecholamine and corticosterone biogenesis, resulting in decreased serum corticosterone levels. However, as this is an assumption, the precise contribution of the 6-gingerol and 6-shogaol to the inhibition of corticosterone synthesis under stressful conditions remains to be determined in further studies.
In conclusion, HPG exhibits antistress effects possibly through mitigation of stress-induced physiological responses and downregulation of stress hormone-related gene expression. The HPG increased body weight gain and thymus weight in the immobilization-stressed rats. Whereas the serum corticosterone concentration and mRNA levels of genes, including Th, Dbh, Pnmt, and P450scc, were lowered by the HPG supplementation. Accordingly, these findings suggest that HPG can be used as a health functional food material for stress relief.
Footnotes
Acknowledgments
This study was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. 2016R1A2B4011021) and Brain Korea 21 PLUS project (No. 22A20130012143).
Author Disclosure Statement
No competing financial interests exist.