
Abstract
Hypertension, one of the famous “silent killers” that can attack people at any age, is a current hot topic among scientists due to multiple syndromic behavior and concomitant diseases. The new scientific-based Traditional Chinese Medicine (TCM) formulation approach was used in a previous study by combining five TCM herbs, including Gastrodia elata Bl., Uncaria rhynchophylla (Miq.) Miq. ex Havil., Pueraria thomsonii Benth., Panax notoginseng (Burk.) F.H. Chen, and Alisma orientalis (Sam.) Juzep in optimized ratio (named BPAid). The objective of the present study was to evaluate the mechanism pathways employed by BPAid for vasodilatory effect with the use of an in vitro isolated aortic rings assay. Interestingly, all the mechanisms investigated were involved in the BPAid's vasodilation activity in which the majority contributed through the nitric oxide/soluble guanylyl cyclase/cyclic guanosine monophosphate (NO/sGC/cGMP) pathways, followed by prostacyclin (PGI2), β 2-adrenergic, and M3-receptors pathways. Furthermore, the BPAid appeared to manage vascular tone by regulating action potential through potassium and both voltage-operated calcium channel and inositol triphosphate receptor (IP3R) pathways. The results obtained has confirmed the expected outcome that the benefits of TCM herbs in BPAid can meet the criteria of counteracting multiple signaling mechanism pathways involved in the etiology of hypertension. In addition to this study, the fingerprints and chemical properties of BPAid was identified by using tri-step Fourier transform infrared spectroscopy and compared with its derivatives. The results obtained suggested that the majority of the vasodilatory effects exerted by BPAid were attributed to the presence of saponins and aromatic ring-containing vasoactive compounds.
Introduction
H
For instance, there was an in vitro study evaluating the efficacy of a new scientific-based formula developed by combining the vasoactive components identified from different TCM herbs in optimized ratios through a series of orthogonal stimulus–response compatibility group studies. The results demonstrated that vasodilatory action achieved from the combined compounds was stronger than the single compounds.
8
Interestingly, this finding was similar to our previous in vitro study on the vasodilatory effects of five TCM herbs [Gastrodia elata Bl., Uncaria rhynchophylla (Miq.) Miq. ex Havil., Pueraria thomsonii Benth., Panax notoginseng (Burk.) F.H. Chen, and Alisma orientale (Sam.) Juzep] (Supplementary Table S1; Supplementary Data are available online at
However, the signaling mechanism pathway employed by the BPAid for its vasodilatory effect remains to be discovered; hence, the main purpose of the present study was to investigate the signaling mechanism pathways by using the isolated rat aortic rings model. Each pathway was investigated by using its respective selective/nonselective antagonists at recommended concentrations. 10,11 In addition, the fingerprints and chemical properties of BPAid were identified by using tri-step Fourier transform infrared (FTIR) spectroscopy and compared among its derivatives. Tri-step FTIR is composed of three fingerprint identification methods: conventional IR (1D-IR), second-derivative IR (SD-IR), and two-dimensional IR (2D-IR), and thus, the spectra obtained from the scannings were extremely reproducible. Moreover, this method is often used to interpret the fingerprint features of herbs since the resolution can be amplified gradually. Besides, the easy and rapid analytical system, and cost-effectiveness provide additional advantages for the tri-step FTIR spectroscopy. 12 –14
Materials and Methods
Preparation of herbs and chemicals
The five TCM herbs: G. elata Bl., U. rhynchophylla (Miq.) Miq. ex Havil., P. thomsonii Benth., P. notoginseng (Burk.) F.H. Chen, and A. orientale (Sam.) Juzep purchased from the local medical hall were authenticated by Dr. Mun Fei Yam (an expert in TCM, Fujian University of Traditional Chinese Medicine, China), and fingerprints were identified by both tri-step FTIR spectroscopy (Supplementary Fig. S1) and thin-layer chromatography (Supplementary Fig. S2). Chemicals purchased included: acetylcholine (Ach) and phenylephrine (PE) (Acros Organics, Belgium); 4-aminopyridine (4-AP) and potassium bromide (KBr) (Merck, Germany); methylene blue (MB) and ethylene glycol tetraacetic acid (EGTA) (Promedipharm Sdn. Bhd, Malaysia); tetraethylammonium (TEA), barium chloride (BaCl2), N
ω
-nitro-
Tri-step FTIR characterization of BPAid and its derivatives
Each extract, namely BPAid, F1, F1-1, F1-3, F1-4, and F1-5, was weighed and ∼1 g was dissolved in distilled water. The dissolved extracts were dried in an oven overnight at 50°C. The extracts were mixed with KBr and compressed as a tablet with ∼10 psi (GS15011, Atlas 15T Manual Hydraulic Press; Specac). KBr was used as a blank. The spectra (1D-IR) of BPAid and its derivatives were recorded at 0.2 cm/sec optical path speed with 16 scans at a resolution of 4 cm−1 within the range of 400–4000 cm−1 wavelength by using a Spectrum 400 FTIR spectrometer (v.6.3.5), which was equipped with DTGS detector (PerkinElmer, USA). The disturbances caused by water and carbon dioxide were eradicated during online scanning. The spectra obtained were considered valid only if more than 60% transmittance could be achieved. The extract pellet was reprepared if the transmittance was less than 60% by adding more KBr or the extract. 15 The SD-IR was implemented by using Savitzky–Golay polynomial fitting (13-point smoothing). The 2D-IR spectra were then determined by fixing the extract tablet into the sample holder with a programmable heated jacket controller (Model GS20730; Specac). Subsequently, the dynamic spectra were recorded at 10°C intervals from 50°C to 120°C. The 2D-IR spectra were obtained by treating the series of dynamic spectra with 2D-IR correlation analysis software developed by Tsinghua University (Beijing, China). 15,16 The spectrum of BPAid (EC50 = 0.028 ± 0.005 mg/mL; R max = 101.71% ± 3.64%) was then compared with the spectra of F1 (EC50 = 0.104 ± 0.014 mg/mL; R max = 97.80% ± 3.12%), F1-1 (EC50 = 0.250 ± 0.101 mg/mL; R max = 95.77% ± 2.75%), F1-3 (EC50 = 0.254 ± 0.067 mg/mL; R max = 93.25% ± 3.98%), F1-4 (EC50 = 0.171 ± 0.022 mg/mL; R max = 99.69% ± 2.12%), and F1-5 (EC50 = 0.377 ± 0.124 mg/mL; R max = 92.74% ± 3.38%), to identify the unique chemical constituents present in BPAid. 9
Animals
Eight- to 10-week-old male Sprague Dawley rats weighing between 180 and 250 g were used throughout the experiment. The experimental animals were acclimatized in the animal transit room at room temperature with 12-h light–12-h dark cycle and free access to water and food. The investigation was in accordance with the Guidelines in the Care and Use of Laboratory Animals by Universiti Sains Malaysia [USM/Animal Ethics Approval/2016/(103) (777)].
Aortic rings preparation
The Krebs–Henseleit (Krebs') solution (118.0 mM NaCl, 4.7 mM KCl, 25.0 mM NaHCO3, 2.5 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 11.0 mM glucose, pH 7.4) was prepared in a Petri dish and aerated with carbogen (95% O2 and 5% CO2). The male Sprague Dawley rat was sacrificed by overdose of CO2 inhalation. An incision was made on the upper abdomen, and the thoracic aorta was isolated immediately and placed in the Krebs solution. The adhering connective and adipose tissues were gently removed, and the aorta was cut into 3–4 mm ring segments. The aortic rings were mounted in the tissue bath containing 10 mL of Krebs solution, by using stainless L-shaped brace and needle hook. The tissue bath chamber was consecutively aerated with carbogen gas with the temperature maintained at ∼37°C. One hook was fixed on the L-shaped brace, whereas the other was connected to a force-electricity transducer (GRASS Force-Displacement Transducer FT03 C Isometric Measurements). The mounted aortic rings were left to equilibrate for 45 min and the Krebs solution was changed every 15 min. The resting tension was adjusted to 1.0 g. The validity of the aortic rings were assured by exposing them to a contractile agent, PE (1 μM), and a relaxing agent, Ach (1 μM), where at least 60% of responses were achieved. After that, the aortic rings were then rinsed three times with Krebs solution at15-min intervals, and the resting tension was readjusted to 1.0 g if necessary.
Vascular response to BPAid with PE precontraction
Once the endothelium-intact isolated aortic rings were validated, 1 μM of PE was applied to obtain a sustained contractile tone, and left for at least 30 min to achieve the plateau stage. Then, the BPAid was added into the tissue bath cumulatively from 0.0025 to 0.3175 mg/mL at 20-min intervals. 17 The contractile and relaxation responses were detected by the force-electricity transducer and applied by quad bridge amplifier (AD instrument, Australia), which would be converted into digital signals by a PowerLab 26T (AD Instrument). The concentration–response curve of BPAid was constructed and was assigned as the control.
Vascular response of endothelium-impaired aortic rings to BPAid
The endothelium dependency was first determined by mechanically removing the endothelium of the aortic rings using a stainless steel stick and gently rubbing on the lumen of the aorta. The complete removal of endothelium was assured by applying the Ach (1 μM) where no relaxation was detected. The PE was applied for precontraction and BPAid was added cumulatively from 0.0025 to 0.3175 mg/mL at 20-min intervals. The concentration–response curve was constructed and the EC50 and R max values obtained were compared with the control group.
Vascular response to BPAid with KCl precontraction
The precontraction of the endothelium-intact isolated aortic rings was induced by applying the 80 mM of KCl instead of PE, and left for at least 30 min until the KCl-induced contractile tone achieved the plateau stage. Then, the BPAid was added cumulatively from 0.0025 to 0.3175 mg/mL at 20-min intervals. The concentration–response curve was constructed and EC50 and R max values obtained were compared with the control group.
Determination of endothelium-derived relaxing factors and nitric oxide cascade antagonists on BPAid vasodilatory effect
Endothelium-intact isolated aortic rings were used to study endothelium-derived relaxing factors (EDRFs) and nitric oxide (NO) cascade signaling mechanism pathways. The antagonists, L-NAME (10 μM), indomethacin (10 μM), ODQ (1 μM), and MB (10 μM), were used to study endothelial NO synthase (eNOS), prostacyclin (PGI2), soluble guanylyl cyclase (sGC), and cyclic guanosine monophosphate (cGMP) mechanism pathways, respectively. The antagonist was preincubated with the aortic rings for 20 min before the PE precontraction. The BPAid was subsequently added cumulatively (0.0025–0.3175 mg/mL) at 20-min intervals once the PE-induced contractile tone had reached the plateau stage. The concentration–response curves for each mechanism studies were constructed, and the EC50 and R max values obtained were compared with the control group.
Investigation of BPAid's vasodilatory effect in the presence of G-protein-coupled receptor inhibitors
There are G-protein-coupled receptors (GPCRs), which play important roles in the vascular activity, such as muscarinic and β 2-adrenergic receptors. The atropine (1 μM) and propranolol (1 μM) were used to study muscarinic and β 2-adrenergic receptors, respectively. The antagonist was preincubated with the endothelium-intact isolated aortic rings in the tissue bath for 20 min before being primed with PE. Once the PE-induced contractile tone had reached a plateau, BPAid was applied cumulatively (0.0025–0.3175 mg/mL) at 20-min intervals. The concentration–response curves for each mechanism study were constructed, and the EC50 and R max values obtained were compared with the control group.
Studies of potassium channels in BPAid vasodilatory activity
Generally, there are four types of potassium channels frequently been studied: calcium-activated K+ channel (Kca), ATP-sensitive K+ channel (KATP), inward rectifier K+ channel (Kir), and voltage-activated K+ channel (Kv). Hence, they were studied by preincubating the endothelium-intact aortic rings with TEA (1 mM), glibenclamide (10 μM), BaCl2 (10 μM), and 4-AP (1 mM), respectively, for 20 min before the PE precontraction. Once the PE-induced contraction reached a plateau, BPAid was applied cumulatively (0.0025–0.3175 mg/mL) at 20-min intervals. The concentration–response curves for each potassium channel studied were constructed, and the EC50 and R max values obtained were compared with the control group.
Measurement of extracellular calcium influx
The extracellular calcium commonly enters into the cytosol by using the L-type calcium channel, which is also referred to as voltage-operated calcium channel (VOCC). Three sets of experiments were carried out in the VOCC mechanism study which included the negative control, positive control (nifedipine), and experimental sets (BPAid). The endothelium-intact isolated aortic rings were utilized throughout this study. The normal Krebs solution was replaced by the calcium-free high-potassium Krebs solution (91.04 mM NaCl, 50.0 mM KCl, 11.9 mM NaHCO3, 1.05 mM MgSO4, and 5.5 mM glucose, pH 7.4). Once the endothelium-intact isolated aortic rings were functionally validated, the aortic rings were rinsed with normal Krebs solution for 10 min. Subsequently, the normal Krebs solution was replaced by EGTA (0.2 mM) containing calcium-free high-potassium Krebs solution for 10 min to remove the calcium residue. The solution was then replaced by calcium-free high-potassium Krebs solution, and rinsed twice at10-min intervals. The resting potential was readjusted to 1.0 g if necessary. The control group was tested by adding the calcium (0.01–10 mM) cumulatively at3-min intervals. The concentration–response curve was constructed and C max value was recorded. This protocol was repeated for positive controls and experimental treatments. However, one additional step was added, at which the aortic rings were preincubated with nifedipine (0.1, 0.3, and 1 μM) in the positive control set, or BPAid (0.005, 0.02, and 0.08 mg/mL) in the experimental set for 20 min before the cumulative addition of CaCl2. The concentration–response curves of positive and experimental groups were constructed, and C max values were recorded and compared with the control group. 10,18,19
Measurement of intracellular calcium release
Typically, one of the major pathways for the intracellular calcium release into the cytosol is through the inositol triphosphate receptor (IP3R) located on the membrane of sacroplasmic reticulum (SR). Similar to the VOCC mechanism studies, three sets of experiments were conducted, which included negative control, positive control (2-APB), and experimental groups (BPAid). The endothelium-impaired isolated aortic rings were used throughout this study. Similar protocols as used in the VOCC mechanism study were implemented in this IP3R study. Briefly, once the endothelium-denuded aortic rings were functionally validated, normal Krebs solution was used to rinse the aortic rings for 10 min, then replaced with EGTA containing calcium-free high-potassium Krebs solution for 10 min, and subsequently rinsed twice by the calcium-free high-potassium Krebs solution at10-min intervals. Hereby, the control group where the PE (1 μM) was applied instead of the CaCl2 (as mentioned in VOCC mechanism study) was allowed for transient contraction for 20 min. The C max value was recorded and used as the control. This protocol was repeated for positive control and experimental sets. However, the additional step was added where the aortic rings were preincubated with 2-APB (100 μM) in positive control, or BPAid (0.005, 0.02, and 0.08 mg/mL) in experimental sets for 20 min before the addition of PE. The C max values were recorded and compared with control. 10,18
Statistical analysis
All results are expressed as mean ± SEM. Results from the controls and the antagonist-pretreated groups were compared by one-way ANOVA, and post hoc Dunnett's test by using SPSS version 22 software. All tests were two-tailed and significance was set at P < .05.
Results
Tri-step FTIR macro-fingerprint of the F1 and its derivatives
The fingerprints of the BPAid and its derivatives (F1, F1-1, F1-3, F1-4, and F1-5) were identified. Figure 1a shows the 1D-IR spectra of all extracts in the range of 400–4000 cm−1. Moreover, the 1D-IR spectra ranging from 750 to 1800 cm−1 of extracts were amplified by using the SD-IR spectroscopy to reveal the hidden characteristics of absorption peaks, which were invisible in the previous 1D-IR spectra, and most of the main absorption bands of chemical constituents were within this range as shown in Figure 1b. Subsequently, the absorption peaks of the extracts are summarized and tabulated in Table 1.
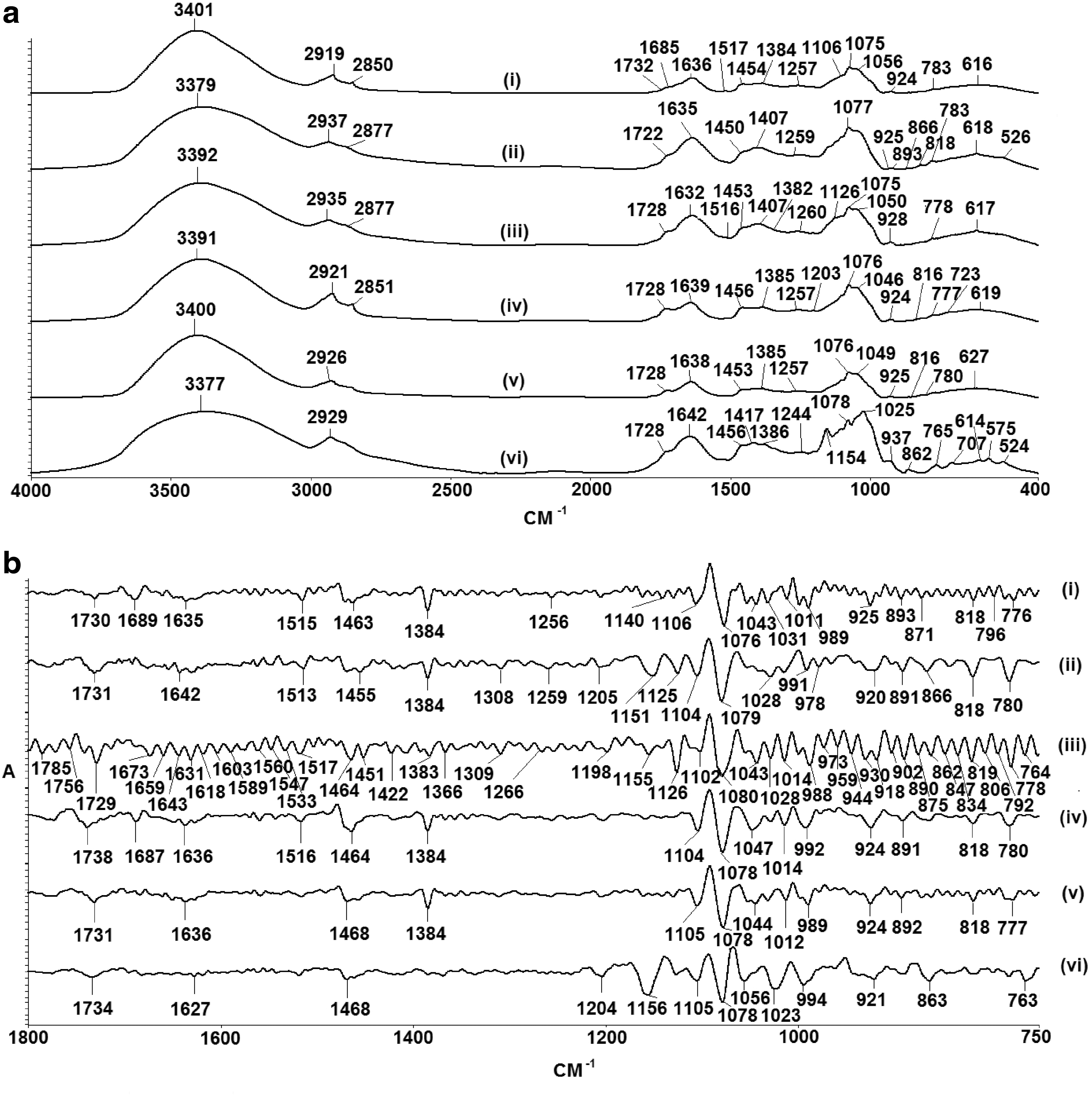
Representative 1D-IR
BPAid, 50% ethanolic extract of five herbs using F1 ratio at 50°C; F1, GE50, UR95, PT95, PN95, and AO50 combined in EC: 0, 25, 20, 20, and 10, respectively; F1-1, distilled water extract of five herbs using F1 ratio at 50°C; F1-3, 95% ethanolic extract of five herbs using F1 ratio at 50°C; F1-4, total 95% of ethanolic extraction of Uncaria rhynchophylla, Pueraria thomsonii, and Panax notoginseng (F1 ratio) combined with D (F1 ratio) at 50°C; F1-5, distilled water extract of five herbs following F1 ratio at 100°C; ν, stretching; ν s, symmetrical stretching; ν as, asymmetrical stretching; δ, bending.
Fundamentally, various organic components within different extracts would behave differently under different thermal perturbation, and these chemical properties could be discovered through the remarkably enhanced resolution of the spectrum obtained from the 2D-IR spectroscopy. 20 Thus, different temperatures were applied to BPAid and its derivatives individually during the 2D-IR spectroscopy scanning, and the dynamic spectra of each extract were obtained and separated into two ranges, which were within 850–1080 and 1080–1600 cm−1 as shown in Figures 2 –4. The range selected for each extract would be varied, which strictly depends on the pattern of the autopeaks that appeared on the dynamic spectra.
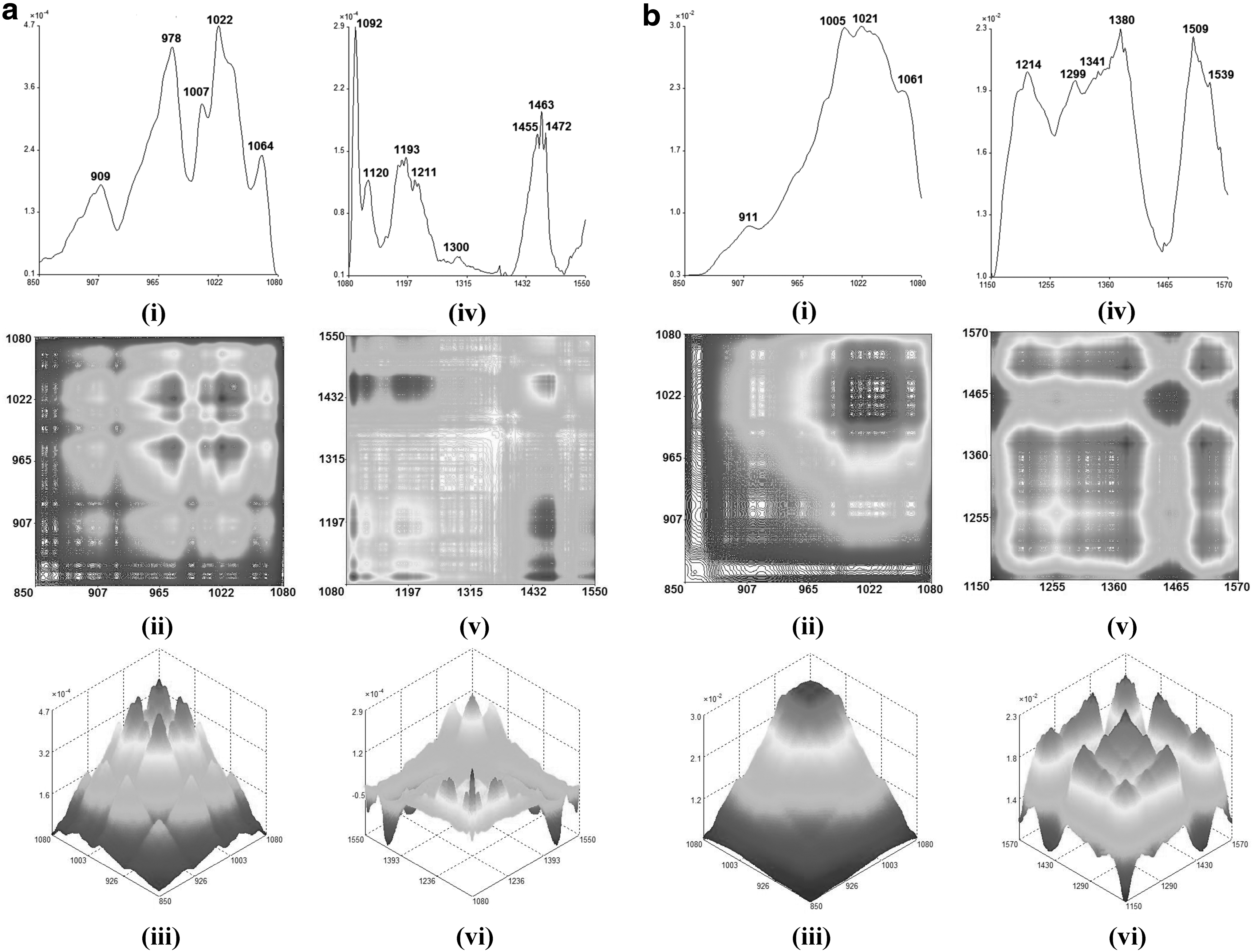
The autopeak curves (i), 2D-IR (ii), and 3D-IR (iii) in the range of 850–1080 cm−1 for F1
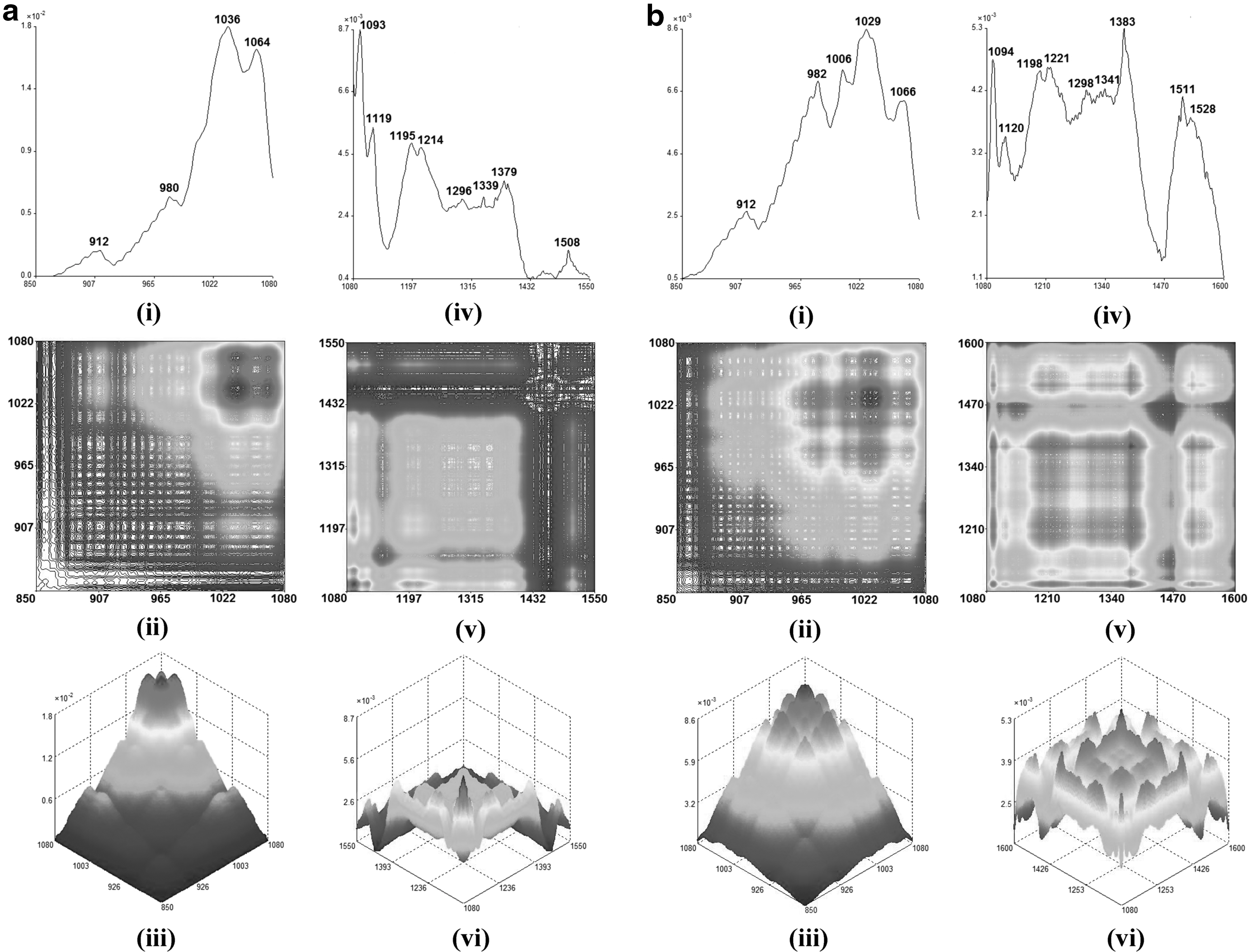
The autopeak curves (i), 2D-IR (ii), and 3D-IR (iii) in the range of 850–1080 cm−1 for BPAid

The autopeak curves (i), 2D-IR (ii), and 3D-IR (iii) in the range of 850–1080 cm−1 for F1-4
Vasodilatory effect of BPAid on PE precontracted vascular tension
A previous study showed that BPAid has exerted the highest vasodilation activity compared with other derivatives. Hence, the vasodilatory effect of BPAid was determined and the result is shown in Table 2. The R max and EC50 values of BPAid in exerting vasodilatory effect were obtained and assigned as the control. Apparently, the vasodilatory effect elicited by BPAid was concentration dependent. The signaling mechanism pathways employed by BPAid for vasodilatory effect was further determined by using the isolated rat aortic rings model.
Significance P < .001.
Significance P < .01
Significance P < .05.
EC50, half of effective concentration; EDRFs, endothelium-derived relaxing factors; GPCRs, G-protein-coupled receptors; L-NAME, N
ω
-nitro-
Primary screening of BPAid on endothelium and channel-linked mechanism pathway's dependency in vascular response
According to Figure 5, in the endothelium-impaired aortic rings, there was ∼33% of R max value reduction compared with the control, whereas the EC50 value is shown in Table 2. This indicated that further mechanism studies were required to elucidate the exact endothelium-dependent pathway employed for BPAid vasodilatory action. Moreover, approximately one quarter of maximal relaxation of BPAid was significantly suppressed on the 80 mM KCl-primed aortic rings compared with the PE-primed, whereas the EC50 value was significantly increased (Table 2). Concentration-dependent trends were observed for vasodilatory action in both endothelium-impaired and KCl precontraction experiments.

Effect on cumulative concentration (0.0025–0.3175 mg/mL) of BPAid on 1 μM PE precontracted endothelium-denuded aortic rings, and 80 mM KCL precontracted isolated endothelium-intact rat aortic rings. Both treatments significantly reduced BPAid vasodilation effects (*P < .05; **P < .01; ***P < .001; n = 8). PE, phenylephrine.
BPAid exerts vasodilatory activity through NO/sGC/cGMP signaling pathways
As shown in Figure 6, the presence of L-NAME and indomethacin in their respective experimental groups markedly attenuated the vasodilatory effect of BPAid compared with the control (Table 2). The results indicated that NO could play a greater role in BPAid endothelium-dependent vasodilatory activity than PGI2. Furthermore, down the NO signaling cascade, the selective antagonists ODQ and MB dramatically reduced the vasodilatory effect of BPAid in their respective tissue baths as shown in Figure 6. However, the EC50 value obtained when ODQ was present was markedly increased up to 13.710 ± 4.370 mg/mL (P < .001) compared with control, which indicated its pivotal role in eliciting BPAid-mediated vasodilatory effects. As shown in Figure 6, the concentration–response curves had shifted to the right in the presence of each individual antagonist.

Effects of 10 μM L-NAME, 1 μM ODQ, 10 μM MB, and 10 μM indomethacin on BPAid-mediated vasodilation effects on 1 μM PE precontracted isolated endothelium-intact aortic rings. All antagonists significantly inhibited BPAid's vasodilation effects (*P < .05; **P < .01; ***P < .001; n = 8). L-NAME, N
ω
-nitro-
Involvement of GPCRs in BPAid vasodilatory activity
As shown in Figure 7, the vasodilatory effect of BPAid in the presence of propranolol was significantly inhibited with increasing of EC50 values compared with control as shown in Table 2. There is another GPCR, which could be mediated in the BPAid vasodilatory effect, the M3 receptor. Its nonselective blocker, atropine, suppressed the vasodilatory effect of BPAid in the PE-primed aortic rings. The trend of inhibition resembled the inhibitory effect of propranolol, but to a lesser extent, as shown in Figure 7. The result indicated the participation of both GPCRs in the BPAid-mediated vasodilatory activity.
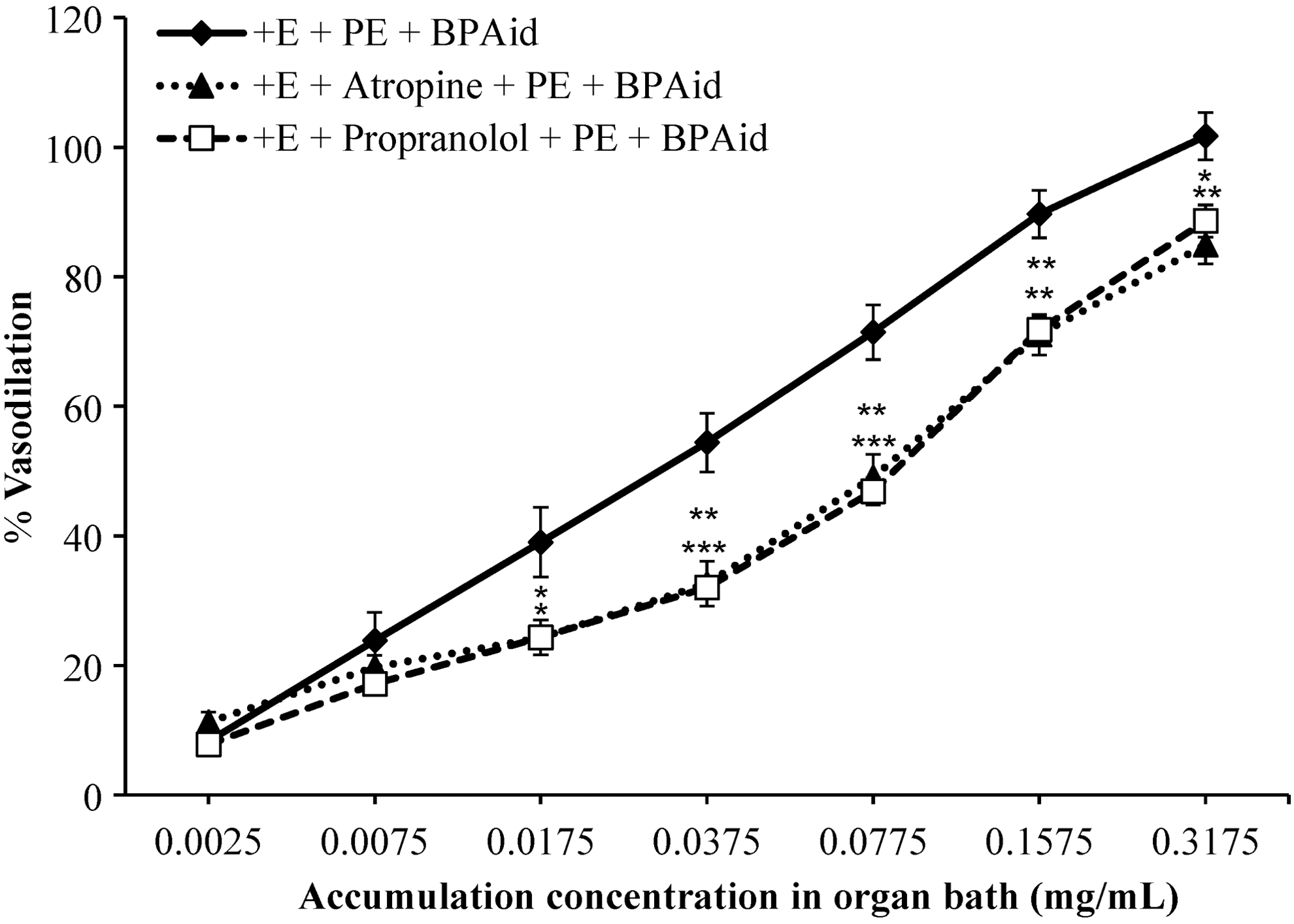
Effects of 1 μM atropine, and 1 μM propranolol on BPAid-mediated vasodilation effects on 1 μM PE precontracted isolated endothelium-intact aortic rings. Both antagonists significantly inhibited BPAid's vasodilation effects (*P < .05; **P < .01; n = 8).
Involvement of potassium channels in BPAid vasodilatory activity
Typically, there are four types of potassium channels frequently studied, which include Kca, KATP, Kir, and Kv channels. As shown in Figure 8, the vasodilatory effect of BPAid was markedly attenuated during the presence of 4-AP and BaCl2 in their respective tissue baths, thus, their EC50 values were significantly increased. Similarly, the presence of TEA and glibenclamide in their respective experimental groups had significantly suppressed the vasodilatory activity of BPAid, hence EC50 values were increased abundantly (Table 2). However, the trend of inhibition for both antagonists were similar, where their antagonizing effect started to decline at BPAid cumulative concentration of 0.1575 mg/mL, therefore, the R max values obtained were slightly reduced to 101.48% ± 5.27% and 94.88% ± 4.93% for TEA and glibenclamide, respectively. The significance of antagonizing effect could be arranged according to their EC50 values such as 4-AP>BaCl2>glibenclamide>TEA, as shown in Table 2.
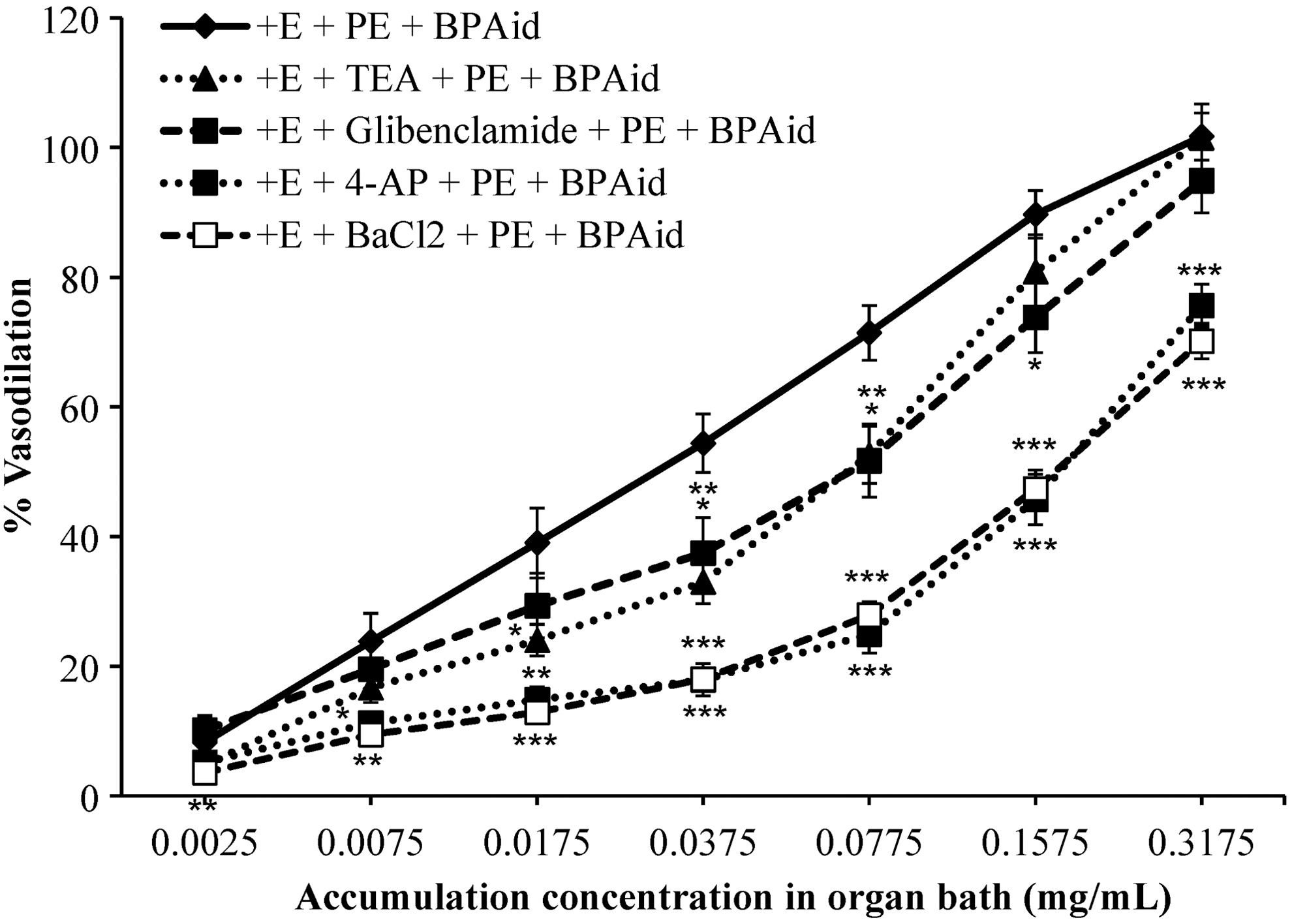
Effects of 1 μM TEA, 10 μM glibenclamide, 1 mM 4-AP, and 10 μM BaCl2 on BPAid's vasodilation effects on 1 μM PE precontracted isolated endothelium-intact aortic rings. All antagonists significantly inhibited BPAid's vasodilation effects (*P < .05; **P < .01; ***P < .001; n = 8). TEA, tetraethylammonium.
Involvement of extracellular and intracellular calcium channels in BPAid vasodilatory activity
There are two major routes for the entrance of calcium into the cytosol, which include extracellular influx through VOCC and intracellular release of calcium mainly through the IP3R located on the SR membrane. According to Table 3, the influx of calcium into the cytosol in the control group, the C max value obtained was 0.811 ± 0.069 g. Preincubation of the endothelium-intact aortic rings with a threefold increase in concentration of a selective blocker, nifedipine (0.1, 0.3, 1 μM), had dramatically suppressed at least 80% of the contractile tone, and almost abolished the contraction tension upon the preincubation with 1 μM of nifedipine compared with the control. Similar significance of the blocking effect was exerted by BPAid at all the three concentrations (0.005–0.08 mg/mL), but to a lesser extent compared with nifedipine as shown in Figure 9a. The blocking effect of BPAid on the vasoconstriction was enhanced gradually with increasing concentrations of preincubated BPAid. A maximum of 51% of contraction tone was abolished compared with the control when the endothelium-intact aortic rings were preincubated with 0.08 mg/mL of BPAid. The C max values achieved upon the preincubation of BPAid at concentrations of 0.005, 0.02, and 0.08 mg/mL are shown in Table 3. Furthermore, Figure 9b shows that the preincubation of 2-APB with the endothelium-denuded aortic rings had abolished ∼97% of the contraction tension compared with controls that originally had a C max value of 0.714 ± 0.027 g, as shown in Table 3. Low concentrations of BPAid (0.005 mg/mL) did not give any clear-cut inhibitory effect on the PE-induced contraction tone. However, the increasing concentration of BPAid applied was significantly reduced up to 50% of contraction tension compared with control. The antagonistic effect of BPAid on both VOCC and IP3R was exerted in a concentration-dependent manner.

Effect of different concentrations of BPAid (0.005, 0.02, 0.08 mg/mL) and nifedipine (0.1, 0.3, 1 μM) on CaCl2-induced contraction in endothelium-intact isolated aortic rings in Ca2+-free Krebs solution. Figure
Significance P < .001.
2-APB, 2-aminoethoxydiphenyl borate; BPAid, 50% ethanolic extract of five herbs using F1 ratio at 50°C; C max, maximal contraction; F1, extracts GE50, UR95, PT95, PL95, and AO50 combined in EC: 0, 25, 20, 20, and 10, respectively; IP3R, inositol triphosphate receptor; Nif, nifedipine; VOCC, voltage-operated calcium channel.
Discussion
Before the experiment, the fingerprints of F1 and its derivatives were analyzed by using tri-step FTIR spectroscopy, which includes 1D-IR, SD-IR, and 2D-IR identification methods. 12 Generally, the 1D-IR spectra of BPAid and its derivatives consist of similar peak position and shape, which indicated that all of them contained similar chemical components as shown in Figure 1a. However, there was slight difference on the spectra at ranges of ∼950–1200 cm−1, in terms of peak intensities and shape. This region belongs to saccharide absorption peaks. 12
Briefly, a hydroxyl absorption peak appeared at 1685 cm−1 in the spectrum of F1, which was not observed in other extract spectras, indicating that F1 contained more acid chemical components. Whereas, F1-3 has a strong characteristic peak of methylene at 2921 and 2851 cm−1, by coupling with the long-chain C-H bending absorption peak at 723 cm−1, suggesting that F1-3 contained certain amounts of unsaturated fatty acids. Interestingly, there were weak aromatic ring absorption peaks in the 1D-IR spectra of both F1 and BPAid, which were not found in other spectra. Hence, it was suggested that the aromatic rings could be one of the major chemical components attributed to the vasodilatory activity since both F1 and BPAid possessed highest vasodilatory effects. Whereas, F1-5 showed strong characteristic peaks of starch at 1642, 1417, 1244, 862, 765, 707, 575, and 524 cm−1, which means the main component of F1-5 is starch and it did not contribute to vasodilatory effect since it exerted the lowest R max value compared with other extracts as discussed in the previous study. 9
The resolution of the spectra was further magnified and analyzed in the range of 750–1800 cm−1 in the SD-IR spectroscopy identification as shown in Figure 1b, to discover the overlapped hidden absorption peaks which could appear as peak bundle and invisible in 1D-IR spectra. For instance, the 1728 cm−1 absorption peak that appeared in 1D-IR spectrum of BPAid has been further divided into 1785, 1756, and 1729 cm−1 in the SD-IR spectrum [Fig. 1b (iii)]. These three absorption peaks belong to the C = O stretching of ester groups, at which 1785 cm−1 is the C = O stretching of lactone, 12 whereas 1756 and 1729 cm−1 are the C = O stretching of general ester groups; however, the peak of 1729 cm−1 is the strongest, indicating that the content of esters are higher than lactones in BPAid. Additionally, there was characteristic peak of saponins found in SD-IR spectrum of F1, BPAid, and F1-4, which could be one of the major sources contributing to their vasodilatory effect. However, the SD-IR spectrum of BPAid has more absorption peaks compared with other extracts, which suggested that large amounts of vasoactive compounds were present in BPAid. Unlike the SD-IR spectrum of F1-5, all the absorption peaks shown in Figure 1b (vi) are the characteristic peaks of starch, such as the peaks located at 1468, 1204, 1156, 1105, 1078, 1056, 921, 863, and 763 cm−1. This phenomenon could be explained by the fact that most of the chemical components were destroyed during the preparation of F1-5 at a temperature of 100°C, and only huge amount of starch was left behind which ordinarily is not involved in vasodilatory effects.
Besides, autopeak and 2D-IR spectra are correlated and often interpreted in two different ranges, where the range selected depends on the pattern of the autopeaks. For instance, BPAid and its derivatives were analyzed from 850–1080 to 1080–1600 cm−1, whereas their synchronous 2D-IR autopeak spectra, contour plots, and mesh plots are shown in Figures 2 –4. According to the 2D-IR spectra obtained for all extracts, a high degree of similarity could be observed in the range of 1080–1550 cm−1, and the difference was found at ∼1460 and 1510 cm−1. The autopeak at 1460 cm−1 was assigned to the C-H bending or C = C stretching bond of aromatic ring skeleton. In 2D-IR spectra, only F1, F1-4, and F1-5 have autopeaks at ∼1460 cm−1, and it has formed negative correlation crosspeaks with other autopeaks. Theoretically, the structure of the aromatic ring skeleton is relatively stable and less susceptible to the influence of temperature, therefore, the autopeak that appeared at ∼1460 cm−1 in spectra of F1, F1-4, and F1-5 belongs to C-H bending bond, while it showed different thermal stability from other autopeaks. Meanwhile, the autopeak presented at around 1510 cm−1 also corresponds to the C = C stretching of the aromatic ring skeleton. The presence of ester and aromatic rings, containing chemical components, as well as the saponins in BPAid, were the main ingredients attributed to its highest vasodilatory effects.
The study on signaling mechanism pathways employed for BPAid-mediated vasodilatory activity was conducted by using isolated aortic rings. 10,11 Aorta is preferred rather than other parts of blood vessels to minimize the variation of smooth muscle cell orientation and avoid the damage to the tunica intima. 21 According to the results shown, the cumulative application of BPAid markedly relaxed the PE-induced endothelium-intact (control), endothelium-denuded, and the KCl-induced vasoconstricted aortic rings during the preliminary screening. These suggested that one-third of the vasodilatory effect was attributed to the EDRFs as well as indicated the involvement of action potential-regulating channel-linked receptors. 16,22 Thus, further mechanism studies were implemented.
Endothelium-intact isolated aortic rings were used to study the EDRFs mechanism pathways such as NO and PGI2, which had been well characterized as essential sources of vasodilatory activity. NO and PGI2 are produced from their respective precursors,
In this experiment, both ODQ and MB were used to study both sGC and cGMP mechanism pathways rather than using MB alone, to differentiate the cGMP-dependent and sGC-dependent vasodilatory effects. 28,29 The sGC signaling mechanism pathway was the major route used for the vasodilatory activity of BPAid down the NO cascade. In addition, the right shifting of the concentration–response curves was observed, which implied a competitive relationship among the antagonists and vasoactive components of BPAid since the blocking effect of the inhibitors could be overcome with the cumulative addition of BPAid.
In spite of the involvement of EDRFs, the endothelium-independent signaling pathways could have contributed to the BPAid vasodilatory activity. One of the important endothelium-independent vasodilatory factors is the β 2-adrenergic receptor, which is usually studied by using nonsteroidal antiinflammatory drugs (NSAID) types of antagonists such as propranolol. This receptor is present on VSMC rather than in endothelium and coupled with Gs α-protein. 23,30 The presence of the propranolol had mildly reduced the vasodilatory effect of BPAid; however, it also significantly increased the EC50 value compared with control. The result suggested that BPAid could stimulate the activation of adenylyl cyclase (AC) for the production of cyclic adenosine monophosphate (cAMP) through the activation of β 2-adrenoreceptor, hence eliciting the vasodilatory effect.
In addition to the similar activation of the vasodilatory signaling mechanism cascade, the increased production of PGI2 promoted by BPAid in the endothelium, tends to bind with the Gs α-protein-coupled IP receptor located on the VSMC, which subsequently activates the AC to allow for the upregulation of cAMP, hence promoting the activation of protein kinase A (PKA), which tends to inhibit the phosphorylation of MLC kinase (MLCK) and halts the vasoconstriction. 31,32 Nevertheless, there is another Gq α-protein-coupled receptor which is located in both endothelium and VSMC in vasculature, but functionally predominates in endothelium, which is M3-receptor. 33 The results obtained suggested the involvement of Gq α-protein-coupled M3-receptor during the vasodilatory activity of BPAid. It could enhance the breakdown of phosphatidylinositol 4,5-bisphosphate (PIP2) into the second messenger, inositol triphosphate (IP3), which binds with IP3 receptor (IP3R) located on the membrane of SR, hence allowing the intracellular influx of calcium into the cytosol that eventually results in upregulation of NO. 25,34 The presence of both antagonists, propranolol and atropine, in their respective tissue baths had exerted the similar inhibitory pattern as shown in the concentration–response curves, suggesting that their involvement in BPAid vasodilatory activity was almost equivalent. Both of the concentration–response curves were shifted to the right, suggesting the competitive relationship among antagonists and vasoactive components of BPAid with the binding sites.
Fundamentally, the vascular tone is strictly regulated by enzyme-linked, G-protein-coupled as well as channel-linked receptors. Potassium and calcium channels are the two major types of channel-linked receptors that control the action potential of blood vessels. Preliminary screening revealed that channel-linked receptors had partially contributed to the vasodilatory effect of BPAid, thus the calcium and potassium channel mechanisms were studied further. Basically, action potential will occur in VSMC rather than in endothelium, therefore, the presence of endothelium can be mostly ignored. 17 According to the results shown, all the potassium channels had been involved in the vasodilatory activity of BPAid by controlling the membrane potential. This could happen because there might be a variety of vasoactive components in the BPAid extract; hence, multiple signaling mechanism pathways could be employed. The results suggested that BPAid is capable of enhancing the hyperpolarizing current of the membrane potential by opening K+ channels. For instance, the activation of the Kca, KATP, and Kv channels to allow the efflux of potassium from the cytosol, thus creating hyperpolarizing current which impels the closure of calcium channels, elicited the vasodilatory effect. Since the Kir channel was one of the potassium channels mediating the vasodilatory activity of BPAid, henceforth at this point (after hyperpolarization stage) the BPAid would tend to hasten the recovery of the electrochemical gradient back to resting potential by activating the Kir channel, which favors the potassium influx from the extracellular rather than efflux. 35 In endothelium, the activation of K+ channels could act as the endothelium-derived hyperpolarizing factor (EDHF), which is claimed to be one of the major EDRFs. 36 Generally, the involvement of the potassium channels in the vasodilatory effect of BPAid could be arranged in descending order such as Kv>Kir>KATP>Kca. The involvement of Kv channel would suggest the involvement of VOCC because both of these channels are intercorrelated in vasculature. Therefore, the involvement of VOCC was further analyzed.
Along with the other vasodilation-mediated mechanism pathways, calcium is one of the essential second messengers in controlling the vascular tone especially in action potential regulation. Typically, there are two main routes for the entry of calcium into the cytosol, which includes extracellular calcium influx through the VOCC, and the intracellular calcium release from the SR store mainly through IP3R. 33,37 Before the commencement of the experiment, EGTA was used to eradicate the calcium residue in the buffer, and the selective antagonist for VOCC and IP3R, nifedipine, and 2-APB were used to assure the validity of the protocol, and were proven to be applicable since the calcium-induced and PE-induced vasoconstriction were almost abolished in the presence of their respective antagonists. According to the results, BPAid significantly suppressed the vasoconstriction in both cases, in a concentration-dependent manner. These results suggest the involvement of both extracellular influx and intracellular release of calcium during the vasodilatory activity of BPAid. The BPAid prevention of entry of calcium into the cytosol could indirectly avoid the upregulation of the formation of calcium–calmodulin complexes as well as the phosphorylation of MLCK, resulting in vasodilatory effects. 23,26 In addition, there was decreased contraction tension after the addition of CaCl2 at a concentration of 3 mM; this could be explained once the contraction had reached maximum, causing the VOCC to close and proceed to the repolarization stage where the potassium starts to efflux.
The signaling mechanism pathways employed for the BPAid vasodilatory activity included all the EDRFs, NO-cascade-mediated vasodilatory signaling messengers, both G-protein-coupled β 2-adrenoreceptor, and M3-receptors. At the same time, BPAid had acted as a potassium channel opener, which induced the hyperpolarizing current in the membrane potential during vascular tone regulation as well as to reduce the depolarizing current by blocking the entry of calcium through VOCC and IP3R into the cytosol.
In conclusion, the vasodilatory activity of BPAid was shown to act through the stimulation of NO/sGC/cGMP cascade, followed by activation of COX for production of PGI2, and subsequently through the G-protein-coupled β 2-adrenergic & M3-receptors mechanism pathways. On the other hand, BPAid used Kca, Kv, KATP, Kir, VOCC, and IP3R channel-linked receptors to regulate the membrane potential of vasculature either by opening the K+ channels in endothelium (EDHF)/VSMC, blockage of Ca2+ influx through VOCC, or inhibiting the intracellular release of Ca2+ through IP3R. The multiple signaling mechanism pathways utilized for the vasodilatory effect of BPAid could be explained due to the variety of aromatic ring-containing vasoactive components as well as saponins present in the BPAid since it is composed of five TCM herbs in an optimized ratio, which provides its holistic benefit. Further preclinical studies such as in vivo tests by oral administration on spontaneous hypertensive rats (SHR) model and toxicity test are necessary to determine the recommended dose of BPAid for the effective treatment of hypertension.
Footnotes
Acknowledgments
The mechanism study was conducted by Y.C.L. The tri-step FTIR spectroscopy identification and interpretation was performed by Y.S.C., C.S.T., and C.H.N. The overall process was guided by M.A. and M.F.Y. Thanks Kindly to the contributions of all co-authors.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.