
Abstract
The innate immune response is an important host primary defense system against pathogens. γ-Oryzanol is one of the nutritionally important phytoceutical components in rice bran oil. The goal of this study was to investigate the effect of γ-oryzanol-rich extract from black rice bran (γORE) on the activation of the innate immune system. In this study, we show that γORE increased the expression of CD14 and Toll-like receptor 4 and enhanced the phagocytic activity of RAW264.7 macrophages. Furthermore, γORE and its active ingredient γ-oryzanol promoted the secretion of innate cytokines, interleukin-8, and CCL2, which facilitate phagocytosis by RAW264.7 cells. These findings suggest that γ-oryzanol in the γORE enhances innate immune responses.
Introduction
T
Pathogen recognition is an essential function of the innate immune response. Toll-like receptors (TLRs) are a type of immune receptor that recognizes a wide range of pathogen-associated molecular patterns, which are expressed on infectious agents. 3 –5 To date, 11 functional TLRs (TLR1–11) have been identified in humans. 5 TLR1, TLR2, TLR4, TLR5, and TLR6 are expressed on the cell surface, while TLR3, TLR7, TLR8, and TLR9 are localized within intracellular vesicles. 6 Among them, TLR4 is usually expressed in macrophages and dendritic cells and is responsible for the detection of lipopolysaccharide (LPS), which is found in the outer membrane of most gram-negative bacteria. 3 Cluster of differentiation 14 (CD14) is a glycosylphosphatidylinositol-anchored membrane protein that acts as a coreceptor for LPS along with TLR4. 7 The activation of the TLR4/CD14 complex is important for inducing intracellular signaling pathways to prime innate immune responses. Thus, the control of TLR4 and CD14 gene expression is critical for the activation of innate immunity.
Rice is major staple foods in Asian countries, including India, Japan, Thailand, and Korea. Rice bran, the outer layer of rice kernel, is considered a waste product, and is often used as a livestock feed. Rice bran oil is well known to contain various bioactive compounds beneficial to health, including γ-oryzanol. 8 –10 It exhibits various pharmacological properties, including lowering of serum cholesterol concentrations and inhibition of platelet aggregation, 11 anti-inflammatory, 12,13 antitumor, 14 antidiabetic, 15 and antioxidant effects. 16 It has been demonstrated that black rice bran extracts display anti-inflammatory activity. 17 –19 Most studies have focused on the anti-inflammatory effects of black rice components on LPS-induced inflammatory responses. 19 However, the functional effect and the molecular mechanism underlying γ-oryzanol-induced innate immune response are not well characterized. γ-Oryzanol is a mixture of ferulic acid esters and phytosterols. 20 In the present study, we prepared a γ-oryzanol-rich extract from black rice bran (γORE) and investigated its effect on the expression of TLR4 and CD14 in macrophages and innate cytokines in monocytes.
Materials and Methods
Materials
LPS (from Escherichia coli 0111: B4, prepared by phenolic extraction and gel-filtration chromatography), bovine serum albumin, and rabbit serum were all purchased from Sigma-Aldrich (St. Louis, MO, USA). Purified γ-oryzanol was purchased from Wako (Pure Chemical Industries Ltd., Osaka, Japan). Recombinant human interleukin-8 (IL-8) was obtained from Life Technologies (Carlsbad, CA, USA).
Preparation of γORE
Black rice bran was obtained from the Korean National Institute of Crop Science (NICS) in 2012. The dried black rice bran (10 kg) was extracted three times with ethanol (9 L) for 3 days. The ethanolic extract (500 g) was obtained and subjected to filtration under reduced pressure. The filtrate was concentrated with a rotary evaporator and redissolved in acetone and allowed to settle in an oven at 60°C for 1 h to obtain clear solution. The solution was subjected to low-temperature separation (LTS) placing in a refrigerator at 2°C for 24 h to crystalize waxes and phospholipids. The upper phase was then collected by filtration and again subjected to LTS in a freezer at −5°C for 24 h to remove insoluble fraction. The solution separated by filtration was concentrated with a rotary evaporator, and the concentrate was dissolved in ethanol and acetone (7:3, v/v) and allowed to solidify for 48 h in a cryogenic freezer at −60°C. The solid phase, which was the γORE, was collected by filtration and washed by hexane. The cleaned γORE was analyzed using an Alliance e2695 HPLC system (Waters Co., Milford, MA, USA) equipped with a YMC PACK ODS-AM reversed-phase column (4.6 × 250 mm I.D., 5 μm; YMC Co., Ltd., Kyoto, Japan) and a 2998 photodiode array detector (a representative wavelength of 325 nm). The mobile phase was methanol:acetonitrile:dichloromethane:acetic acid (50:44:3:3, v/v/v/v) and the flow rate was 1.4 mL/min constantly.
Cell culture
RAW264.7 rat macrophages and the THP-1 human monocytic leukemia cell line were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in Dulbecco's modified Eagle's medium and Roswell Park Memorial Institute medium supplemented with 10% heat-inactivated fetal bovine serum (CellGro/Corning, Manassas, VA, USA), 2 mM
Cytotoxicity assay
The cytotoxicity of γORE was evaluated using a Cell Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Gaithersburg, MD, USA) according to the manufacturer's instructions. In brief, exponentially growing cells were treated with γORE (0, 10, 50, 100, or 200 μg/mL) for 24 h, followed by the addition of the CCK-8 solution for an additional 1 h. The absorbance at 450 nm was measured with an Emax Endpoint ELISA Microplate Reader (Molecular Devices, Sunnyvale, CA, USA).
Reverse transcription-polymerase chain reaction analysis
Total RNA isolation and first-strand cDNA synthesis were carried out as described previously.
21
The Reverse transcription-polymerase chain reaction (RT-PCR) primers were synthesized by Macrogen (Seoul, Republic of Korea): glyceraldehyde 3-phosphate dehydrogenase (GAPDH) forward, 5′-ACCCACTCCTCCACCTTTG-3′; GAPDH reverse, 5′-CCCAGCAAGAGCACAAGAG-3′; CD14 forward, 5′-CGAGGACCTAAAGATAACCGGC-3′; CD14 reverse, 5′-GTTGCAGCTGAGATCGAGCAC-3′; TLR4 forward, 5′-ACAGAAGCTGGTGGCTGTG-3′; TLR4 reverse, 5′-TCTTTAAATGCACCTGGTTGG-3′; IL-8 forward, 5′-AAACAGATGAAGTGCTCCTTCCAGG-3′; IL-8 reverse, 5′-TCTTGCACAAATATTTGATGC-3′; CCL2 forward 5′-CTTCTGTGCCTGCTGCTCATAG-3′; and CCL2 reverse, 5′-CTGGACAAGCAAACCCAAAC-3′. The amplified products were subjected to 1% agarose gel electrophoresis. The band intensities were quantified using the ImageJ software (National Institutes of Health, Bethesda, MA, USA;
Phagocytosis analysis
Phagocytic capability was measured using fluorescently labeled carboxylate-modified polystyrene latex beads (2.0 μm in size) opsonized in 10% fetal bovine serum, according to the manufacturer's instructions (Sigma-Aldrich). A previous study demonstrated that treatment with 0.1 μg/mL concentration of LPS stimulates phagocytic activity of RAW264.7 macrophages, 22 and 10–100 ng/mL doses of IL-8 stimulate innate immune responses. 23 In this study, RAW264.7 macrophages were treated with vehicle, LPS (0.1 μg/mL), IL-8 (20 ng/mL), or γORE (100 μg/mL) for 24 h, after which opsonized latex beads (2 × 108 beads/mL) were added. After 3 h, the cells were washed to remove extracellular beads, and the uptake of latex beads was imaged using an EVOS FL Auto Cell Imaging System (Life Technologies/Thermo Fisher Scientific). The phagocytic index was calculated as the number of cells with at least two beads as a percentage of the total number of cells.
Protein array analysis
Cytokines secreted from THP-1 cells into the culture medium were measured using a Proteome Profiler Antibody Array (ARY005B; Bio-Techne, Minneapolis, MN, USA), according to manufacturer's instructions. In brief, THP-1 cells were treated with 100 μg/mL γORE for 24 h, and then, culture supernatants were collected. The arrays were incubated with ∼2 mL of culture supernatant for 4 h, followed by extensive washing to remove unbound proteins. The secondary antibody reagents supplied in the array kit were incubated on a rocking platform shaker for 2 h. Protein spots were visualized using a chemiluminescence detection reagent (Amersham Pharmacia Biotech, Piscataway, NJ, USA). The average intensity of each duplicated array spot was quantified using ImageJ.
Statistical analysis
Statistical significance was determined by one-way ANOVA followed by Sidak's multiple comparisons test using GraphPad Prism version 7.0 (GraphPad Software, Inc., La Jolla, CA, USA). Data are presented as mean ± standard deviation. A P value less than .05 was considered significant.
Results
Identification of γ-oryzanol in γORE
γORE was prepared as described in the Materials and Methods Section. To confirm the γ-oryzanol components in the γORE, high-performance liquid chromatography was used. By comparison with authentic standards, 10 γ-oryzanol components were identified as follows: peak 1, Δ7-stigmastenyl ferulate; peak 2, stigamsteryl ferulate; peak 3, cycloartenyl ferulate; peak 4, 24-methylenecycloartanyl ferulate; peak 5, Δ7-campestenyl ferulate; peak 6, campesteryl ferulate; peak 7, Δ7-sitostenyl ferulate; peak 8, sitosteryl ferulate; peak 9, campestanyl ferulate; and peak 10, sitostanyl ferulate (Fig. 1). The chemical structures of γ-oryzanol components identified in γORE are shown in Figure 2.
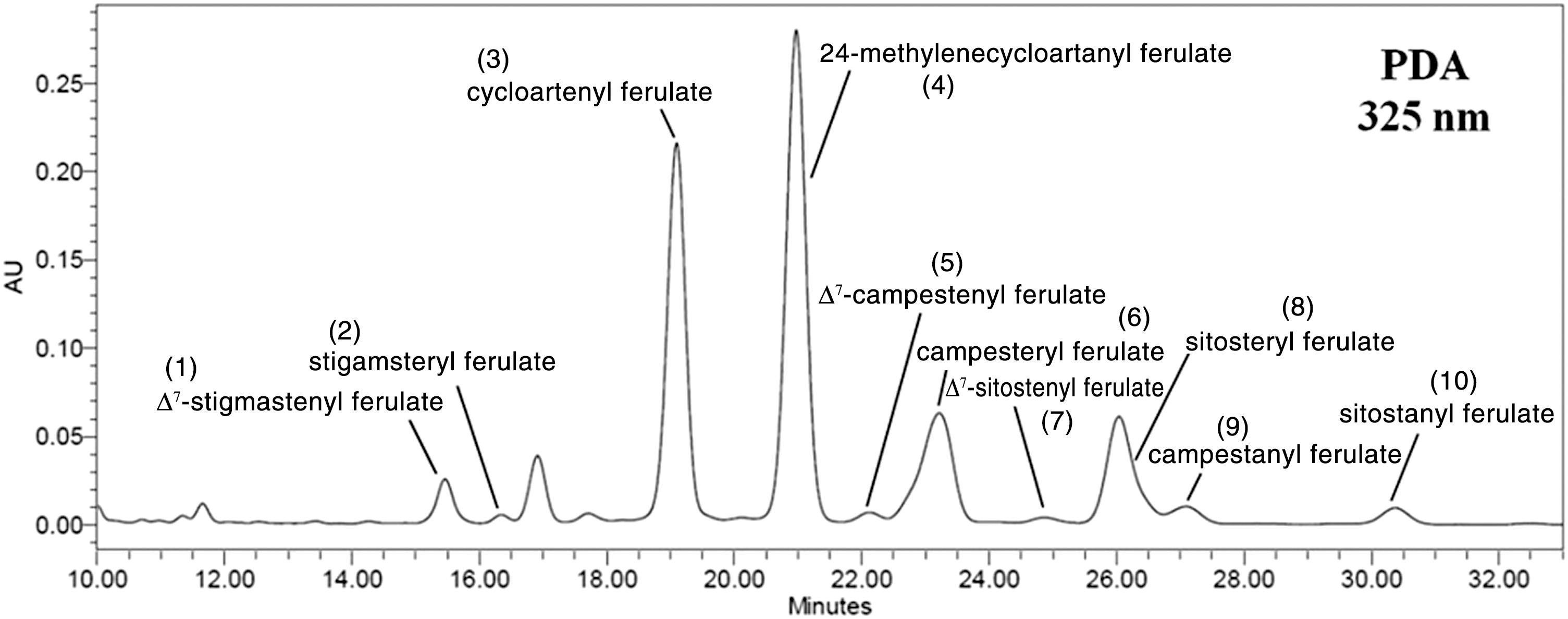
High-performance liquid chromatography chromatogram of γ-oryzanol-rich extract isolated from black rice bran (γORE). The peaks showing γ-oryzanol are indicated. PDA, photodiode array detector.
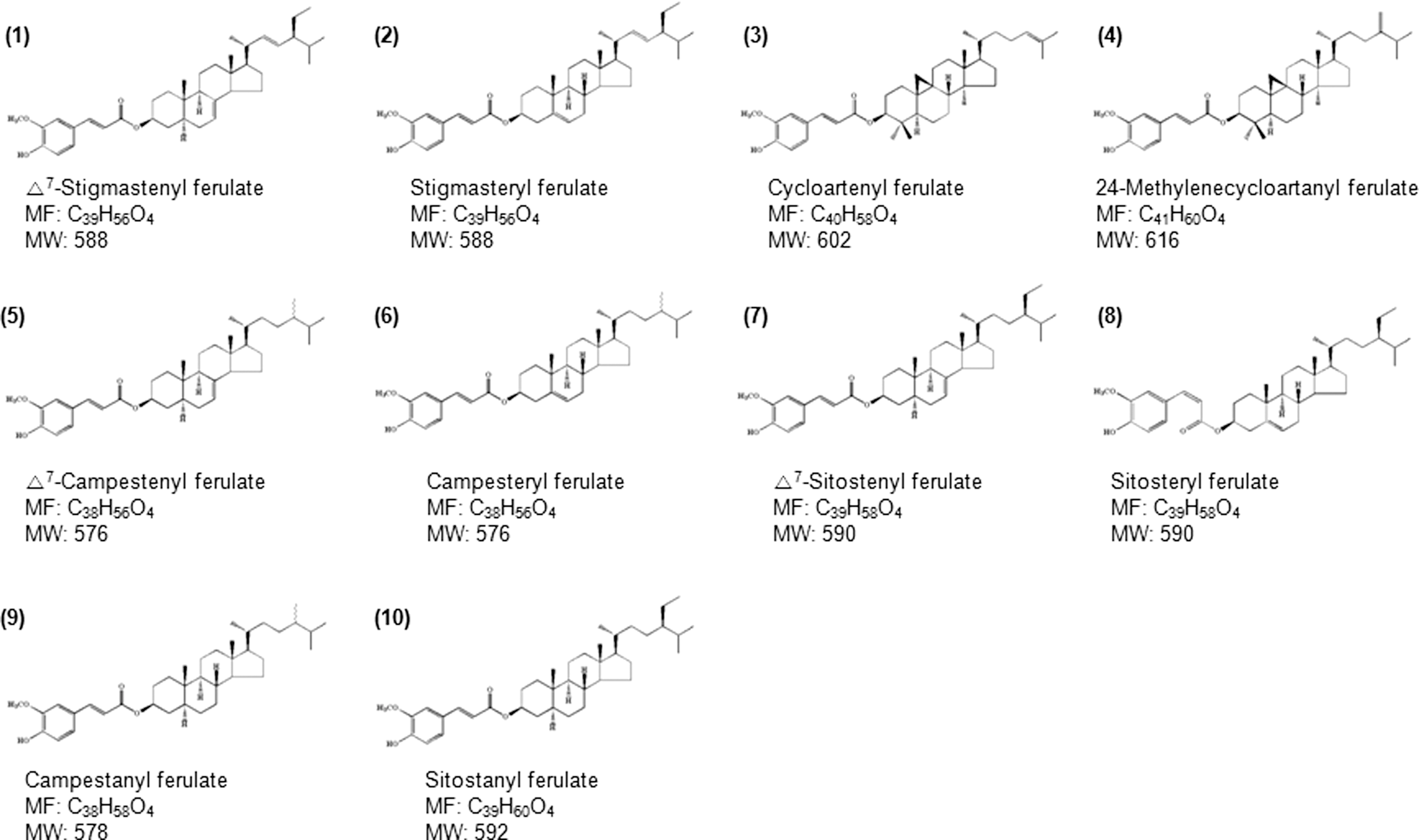
Chemical structure of γ-oryzanol compounds in the γORE. MF, molecular formula; MW, molecular weight.
Effect of γORE on phagocytic activity in RAW264.7 macrophages
It has been reported that γ-oryzanol has an anti-inflammatory effect on intestinal inflammation. 12 However, it remained unclear whether γ-oryzanol enhances the innate immune response. In general, the innate immune response is activated by the sequential action of LPS coreceptors, namely CD14, TLR4, and LPS-binding protein (LBP). TLR4 expression is important in pathogen recognition and in the activation of innate immune signaling pathways. 24 CD14 acts as a TLR4 coreceptor. 7 To determine whether the γORE stimulates the innate immune response, THP-1 monocytes were treated with γORE, and the mRNA expression of CD14 and TLR4 was examined using RT-PCR. LPS was used as a positive control. mRNA expression of both CD14 and TLR4 was barely detectable in nonstimulated conditions. Treatment with γORE substantially elevated the mRNA expression of both CD14 and TLR4 compared with that in vehicle-treated cells (Fig. 3A), suggesting that γORE may activate the innate immune response. No cytotoxic effect was observed after exposure to γORE at concentrations up to 200 μg/mL (Fig. 3B).
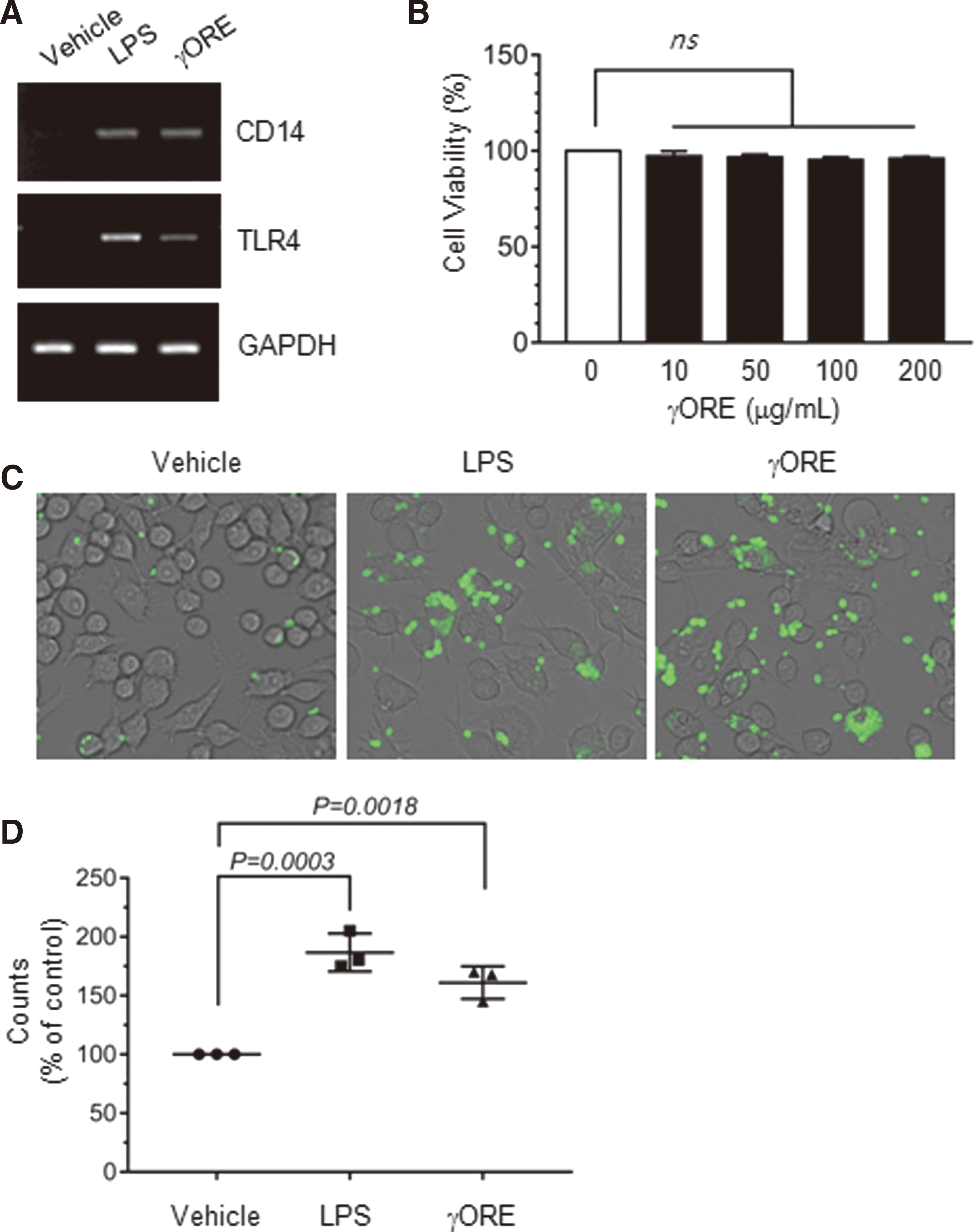
Effects of γORE on the stimulation of innate immune responses.
Phagocytosis is a crucial function of mature macrophages, which can trigger innate immune responses. To evaluate whether γORE stimulates phagocytic activity, RAW264.7 macrophages were incubated with latex beads tagged with a fluorescent dye in the presence or absence of γORE. LPS was used as a positive control. Exposure to 0.1 μg/mL LPS or 100 μg/mL γORE for 24 h potentiated the phagocytic uptake of the fluorescent beads compared with vehicle treatment (Fig. 3C). Relative phagocytic activity was determined by counting cells containing intracellular fluorescent beads. The phagocytic index was increased by 186.7% ± 16.07% (P = .0003 vs. vehicle, ANOVA followed by Sidak's test) and 161.0% ± 13.89% (P = .0018 vs. vehicle, ANOVA followed by Sidak's test) by exposure to LPS and γORE, respectively, compared with vehicle treatment (Fig. 3D). These data suggested that γORE ameliorates the innate immune response.
Effect of γORE on the expression of innate cytokines in THP-1 monocytes
Chemokines play a pivotal role in regulating immune responses. To evaluate the effect of γORE on the expression of inflammatory cytokines by monocytes, THP-1 cells were treated with 100 μg/mL γORE for 24 h, and cytokines secreted into the culture medium were detected and quantified using a cytokine antibody array consisting of a nitrocellulose membrane spotted with 36 cytokine antibodies in duplicate (Supplementary Fig. S1 and Fig. 4A; Supplementary Data are available online at
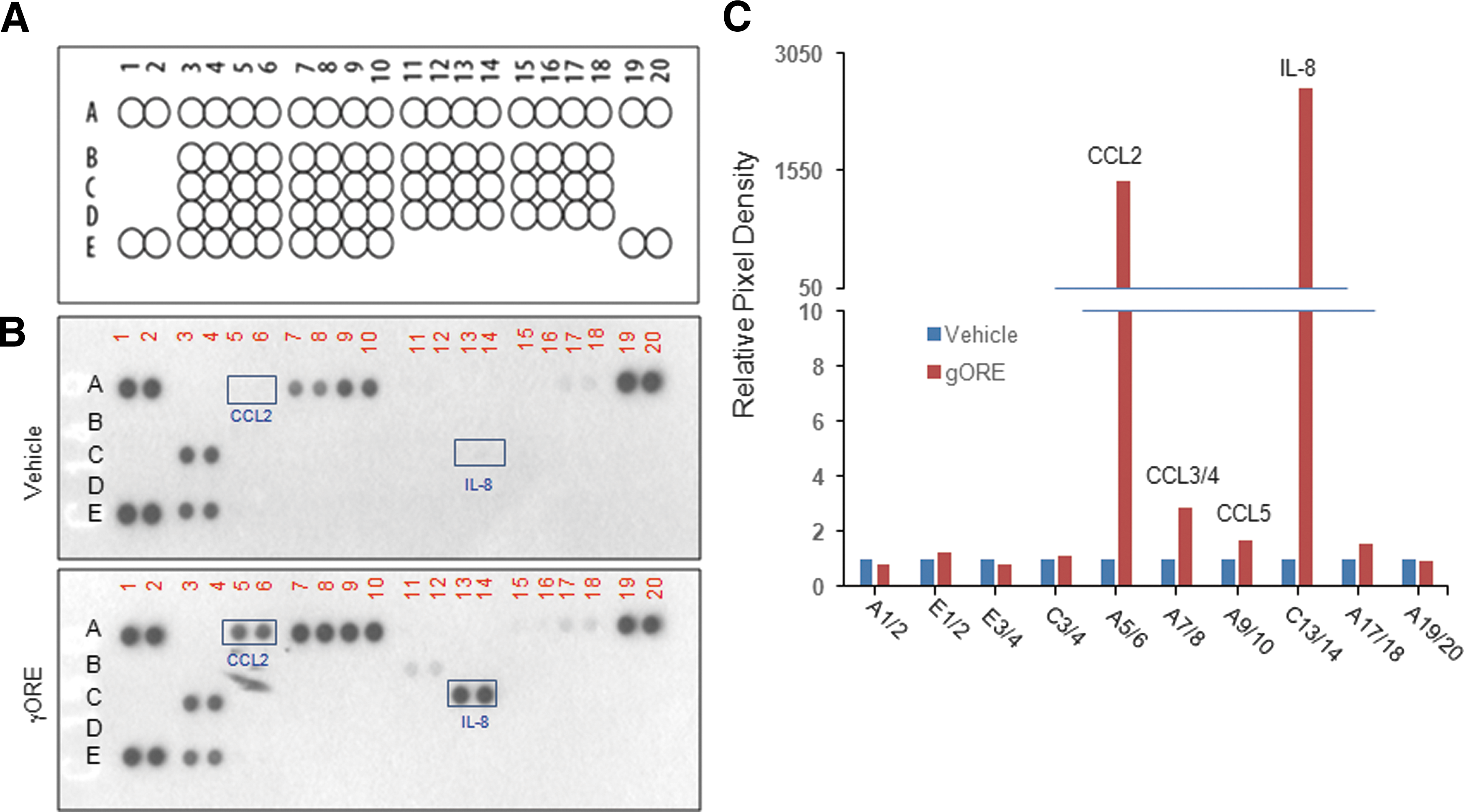
Effect of γORE on innate cytokine production.
Effect of γORE on the expression of IL-8 and CCL2 in THP-1 monocytes
CCL2 is a member of the CC chemokine subfamily of chemotactic chemokines, which plays an important role in recruiting innate immune cells. 25,26 IL-8 is an important innate cytokine that induces various cellular responses, including chemotaxis and activation of various immune cells. 27 We confirmed the effect of γORE on the upregulation of CCL2 and IL-8 mRNA expression in THP-1 cells by using RT-PCR. LPS (0.1 μg/mL) was used as a positive control. As shown in Figure 5A, the mRNA levels of IL-8 and CCL2 increased in a dose-dependent manner following γORE treatment. Quantitative densitometry showed that IL-8 mRNA increased 3.2 ± 0.30 (P = .0034), 5.7 ± 0.76 (P < .0001), and 6.9 ± 0.51 (P < .0001) times (Fig. 5B), and CCL2 mRNA levels increased 1.5 ± 0.25 (P = .5089), 3.0 ± 0.47 (P < .0007), and 6.8 ± 0.30 (P < .0001) times (Fig. 5C), after treatment with 10, 50, and 100 μg/mL γORE, respectively, compared with vehicle treatment. These data suggested that γORE stimulates the production of innate cytokines in monocytic THP-1 cells.
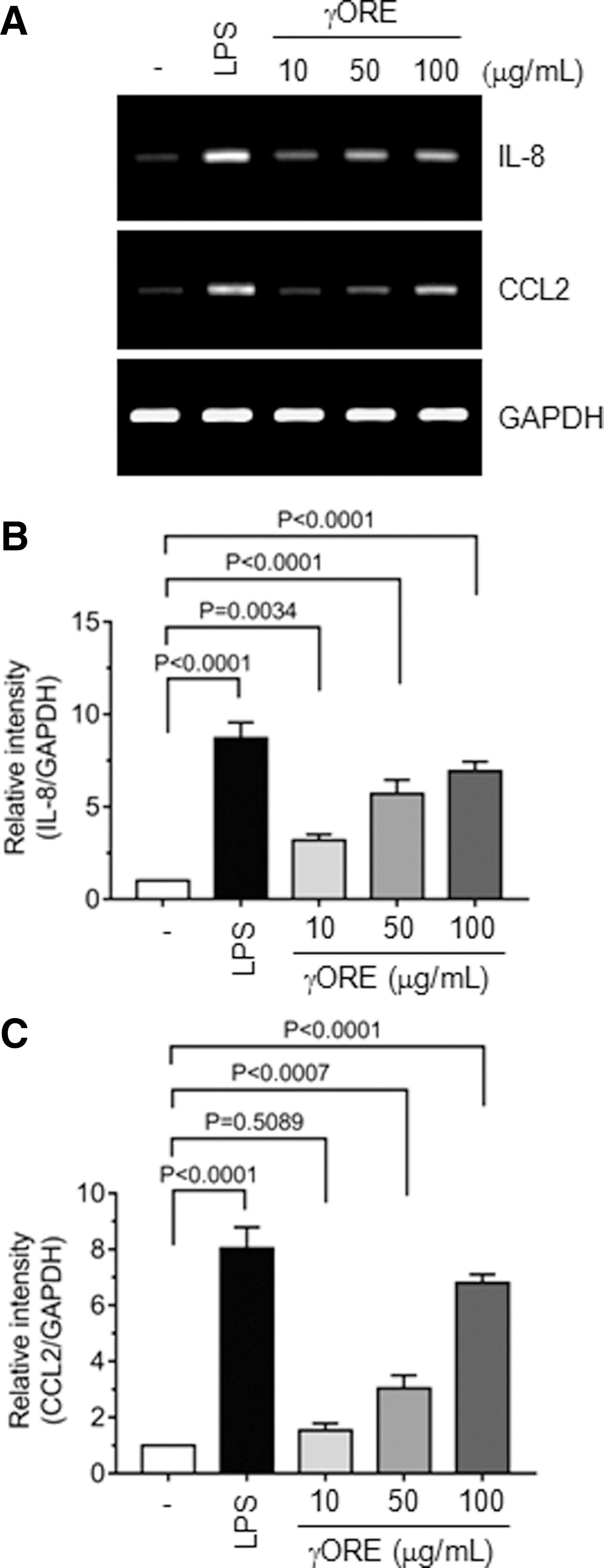
Effects of γORE on IL-8 and CCL2 production.
Effect of γ-oryzanol on the expression of IL-8 and CCL2 in THP-1 monocytes
To evaluate the role of γ-oryzanol in the γORE-induced stimulation of innate immunity, we utilized commercially available, purified γ-oryzanol. LPS (0.1 μg/mL) was used as a positive control. Treatment of THP-1 cells with γ-oryzanol (10–100 μg/mL) resulted in the accumulation of IL-8 and CCL2 mRNAs (Fig. 6A). IL-8 mRNA was increased 3.4 ± 0.28 (P = .0006), 4.3 ± 0.32 (P < .0001), and 5.6 ± 0.92 (P < .0001) times by 10, 50, and 100 μg/mL γ-oryzanol, respectively, compared with vehicle treatment (Fig. 6B). Similarly, CCL2 mRNA levels increased 3.3 ± 0.17 (P < .0001), 3.5 ± 0.20 (P < .0001), and 4.8 ± 0.42 (P < .0001) times after treatment with 10, 50, and 100 μg/mL γ-oryzanol, respectively (Fig. 6C). These data suggested that γ-oryzanol in the γORE contributes to the stimulation of innate cytokine gene expression.
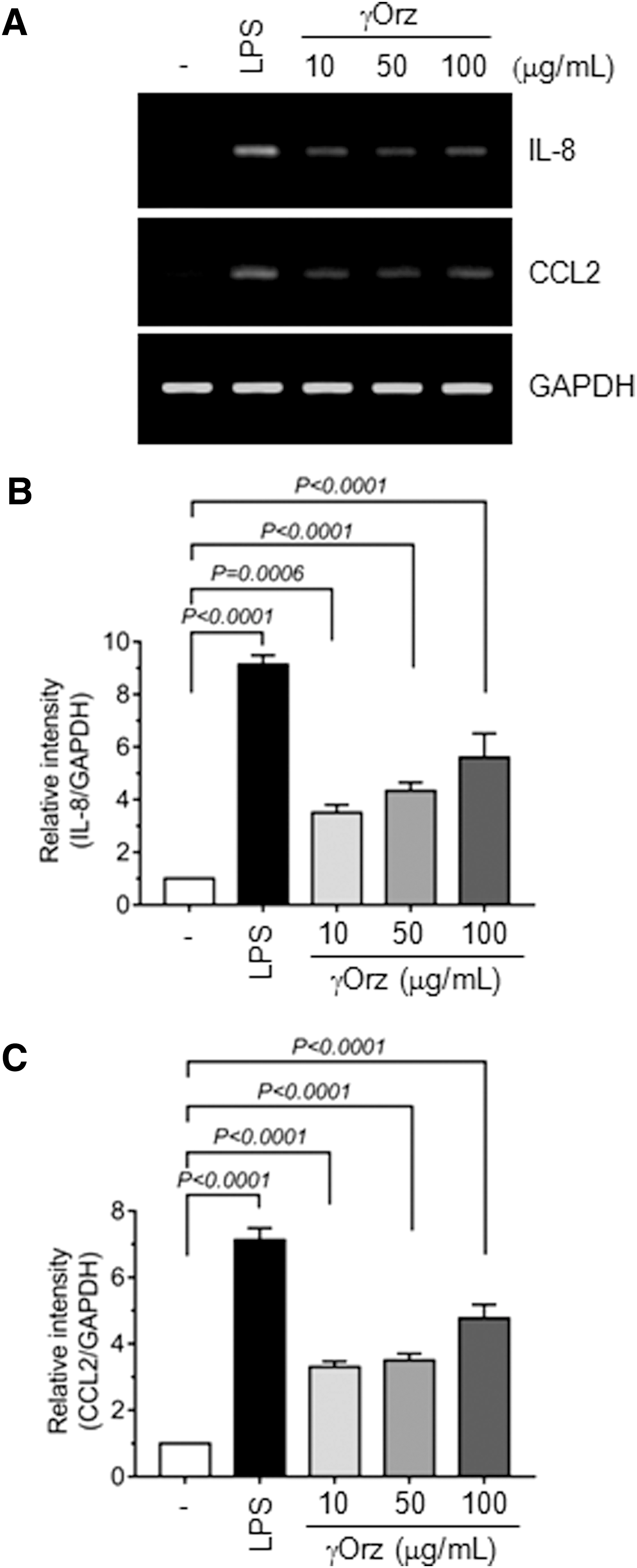
Effects of commercial, pure γ-oryzanol (γOrz) on IL-8 and CCL2 mRNA expression.
Effect of γ-oryzanol on phagocytic activity in RAW264.7 macrophages
The effects of γ-oryzanol and IL-8 induced by γ-oryzanol on phagocytosis were examined using fluorescent latex beads. γORE (100 μg/mL) was used as a positive control. RAW264.7 cells were treated with 100 μg/mL γ-oryzanol or 20 ng/mL IL-8 for 24 h. Phagocytic uptake of the fluorescent beads was increased upon exposure to γORE, γ-oryzanol, or IL-8, compared with vehicle treatment (Fig. 7A). Relative phagocytic activity was determined by counting the cells containing fluorescent beads. The percentage of cells that phagocytosed beads was 178% ± 9.54% (P = .0002), 162% ± 18.2% (P = .0009), and 177% ± 13.8% (P = .0002) upon exposure to γORE, γ-oryzanol, and IL-8, respectively (Fig. 7B). These data suggested that γ-oryzanol plays an important role in the ameliorative effect of γORE on the innate immune response.

Effects of γOrz on the stimulation of innate immune responses.
Discussion
Most previous studies have focused on anti-inflammatory effect of γ-oryzanol on LPS-induced inflammation. 12,17,19,28 The present study investigates the effect of γORE on the innate immune responses. Innate immunity is a nonspecific defense mechanisms that comes into play immediately, while adaptive immunity refers to antigen-specific immune response. Although innate immune responses are important for host protective mechanisms, chronic uncontrolled innate immune responses could lead to local and systemic harmful side effects on host tissues. Thus, immediate and efficient removal of pathogens is important for the protective functions against foreign pathogens. In the present study, we have found that γORE and its major component γ-oryzanol directly stimulates the phagocytic activity of RAW264.7 macrophages and facilitates the expression of TLR4 and innate cytokines, including CCL2 and IL-8, in THP-1 monocytes. These results suggest that γORE could be effective in promoting host primary surveillance system, contributing to reduce the risk of infectious inflammatory disease.
TLRs are important for recognition of many pathogenic microorganisms, and thereby plays an essential role in protective immune responses. 29 Of these, TLR4 recognizes bacterial LPS and triggers the innate immune responses through the activation of many transcription factors, including nuclear factor-κB (NF-κB), AP-1, and IRF3, 29 a prerequisite to the induction of adaptive immune responses. In this study, we determined the effect of γORE on the expression of TLR4 in monocytes. Our result showed that γORE upregulates both TLR4 and its coreceptor CD14 expression with no cellular cytotoxicity. It seems that γORE could be a beneficial ingredient to enhance the ability of initial recognition of invading pathogens.
Chemotactic migration of immune cells to the infection sites is important for the control of proper immune responses. Chemokines are a family of cytokines involved in the regulation of chemotaxis of various leukocytes to a site of infection. There are various types of chemokines, including CXC-motif and CC-motif chemokines. Of these, CCL2 is a member of the CC chemokine subfamily of chemotactic chemokines, which is involved in recruiting innate immune cells, such as dendritic cells, monocytes, and T-lymphocytes, to the inflammatory sites. 30 CCL2 is primarily secreted by monocytes, macrophages, and dendritic cells, and it plays an essential role in enhancing macrophage activities and bacterial clearance. 31,32 IL-8, also known as CXCL8 or monocyte-derived neutrophil chemotactic factor, is an important innate cytokine produced by monocytes and macrophages. 33 In humans, IL-8 induces migration and phagocytosis of macrophages, chemotaxis of neutrophils to the infection sites, leading to the activation of various cellular innate immune responses. 34 Our data showed that γORE and its major component γ-oryzanol effectively enhanced CCL2 and IL-8 expression in monocytes, as revealed by antibody array analysis, RT-PCR, and real-time PCR, suggesting that γORE may help to stimulate immune responses through enhancing innate immune cytokines in unstimulated immune cells.
At present, it remains to be determined how γORE stimulates the innate genes (TLR4, CCL2, and IL-8). Several signaling pathways, including NF-κB and the mitogen-activated protein kinases pathways, are linked to innate immune responses. 35 NF-κB subunits are the main positive regulators of innate immunity, triggering expression of numerous cytokines, chemokines, and inflammatory enzymes. 36 Previous reports have demonstrated that CCL2 expression is upregulated via NF-κB activation in response to stimulation with inflammatory cytokines such as IL-1 and TNFα in astrocytes 37 and kidney mesangial cells, 38 whereas multiple pathways are involved in NiCl2-induced CCL2 expression in endothelial cells. 39 These results suggest that CCL2 expression is regulated by multiple signaling pathways in cell context- and signal-dependent manners. As it has been demonstrated that γ-oryzanol inhibits LPS-induced adhesion molecule mRNA expression via NF-κB inhibition in endothelial cells as well as in monocytes, 28 and in RAW264.7 macrophages, 40 it seems likely that γORE-induced expression of innate cytokines might be, at least in part, mediated independently of the NF-κB-mediated pathway in monocytes. Further study will be required to fully elucidate the molecular signaling pathways mediating γORE-induced innate gene expression.
In summary, we found that γORE has an immunostimulating activity through upregulation of innate genes, including pathogen recognition receptor TLR4/CD14 as well as the innate cytokines CCL2 and IL-8 in monocytes, which can lead to enhancing innate immune responses. Purified γ-oryzanol exhibited similar effects. The innate immune system is a host defense system that protects against infectious microorganisms and other foreign invaders. Our results suggest that γORE might be beneficial for enhancing the innate immunity to prevent various pathogen-induced illness through the expression of innate cytokines. This study reinforces the medicinal effects of black rice bran extracts. γORE could be developed as a dietary supplement for enhancing the innate immune response. Further research is necessary to confirm the effects of γORE on the enhancement of innate immunity by using animal models.
Footnotes
Acknowledgments
This study was supported by the Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ011884012016; Rural Development Administration), Republic of Korea. This article was written as part of Konkuk University's research support program for its faculty on sabbatical leave in 2015.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.