
Abstract
Exercise enhances mitochondrial biogenesis in skeletal muscle. Increased mitochondrial function and content can contribute to the improvement in skeletal muscle function and the benefits of exercise by increasing the response to energy demands. The effect of standardized Kaempferia parviflora extract (KPE) on exercise performance was accessed in L6 myotubes and C57BL/6J mice. KPE significantly activated peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) and increased mitochondrial density in L6 myotubes. KPE also upregulated the expression of transcription factors for mitochondrial biogenesis (estrogen-related receptor-α [ERRα], nuclear respiratory factor-1 [NRF-1], and mitochondrial transcription factor A [Tfam]) through activation of PGC-1α in L6 myotubes. In vivo models including normal diet mice and high-fat diet obese mice showed that KPE effectively enhanced running endurance and increased the skeletal muscle weight/body weight ratio. Furthermore, these observations were associated with a significant upregulation of mitochondrial biogenesis regulatory genes in skeletal muscle tissue. KPE enhanced the protein expression of the sirtuin 1 (SIRT1)/adenosine monophosphate (AMP)-activated protein kinase (AMPK)/PGC-1α/peroxisome proliferator-activated receptor-δ (PPARδ) signaling pathway components in vitro and in vivo, acting as an exercise metabolism regulator. These results suggest that KPE has the potential to enhance exercise performance through mitochondrial biogenesis and the SIRT1/AMPK/PGC-1α/PPARδ signaling pathways.
Introduction
M
Peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) is essential for skeletal muscle energy metabolisms such as fatty acid β-oxidation, muscle fiber-type switching, and energy substrate transport, and is known as the primary stimulator of mitochondrial biogenesis. 4 PGC-1α exerts its effects by interacting with and coactivating various transcription factors. Particularly, PGC-1α interacts with estrogen-related receptor-α (ERRα) and nuclear respiratory factor-1 (NRF-1), sequentially triggering upregulation of mitochondrial transcription factor A (Tfam). ERRα and NRF-1 mediate the expression of the nuclear genes encoding mitochondrial enzymes and proteins, while Tfam controls the expression of mitochondrial encoded genes. In turn, these nuclear-encoded markers contribute to an increase in mitochondrial function, resulting in the activation of mitochondrial biogenesis and the enhancement of exercise performance. 1
PGC-1α is a downstream marker regulated by sirtuin 1 (SIRT1) and adenosine monophosphate (AMP)-activated protein kinase (AMPK). SIRT1 acts a sensor in response to metabolic stimuli, such as exercise and calorie restriction. 5 –7 Similar to SIRT1, AMPK is the prime sensor of fuel and energy status in skeletal muscle, and is stimulated by an elevation in the AMP/ATP ratio. AMPK is promoted by exercise, metabolic stress, hypoxia, and muscular contraction. 5,6 Peroxisome proliferator-activated receptor-δ (PPARδ), the most abundant form of PPARs in skeletal muscle, binds with PGC-1α and is highly involved in the development of oxidative type I muscle fibers. PPARδ improves exercise performance by almost 100% in untrained mice. 8 Furthermore, AMPK directly controls transcriptional activity of PPARδ in skeletal muscle. The interplay between AMPK and PPARδ reprograms the skeletal muscle transcriptome, which acts as an exercise mimetic. 8 Therefore, exercise performance is regulated by activating skeletal muscle mitochondrial biogenesis through upregulating the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway.
Kaempferia parviflora Wall. ex. Baker (K. parviflora), belonging to the Zingibercaceae family, is commonly referred to as black ginger and has been traditionally used to treat various illnesses, such as allergies, fatigue, sexual dysfunction, and ulcers. 9 –11 K. parviflora has been reported to exhibit antioxidative, anti-inflammatory, gastro-protective, anticancer, and antiviral effects. 10 Moreover, K. parviflora enhanced vasorelaxant activity, implying its beneficial effect on exercise performance. 9 The current study aimed to investigate whether K. parviflora extract enhances exercise performance by stimulating mitochondrial biogenesis and the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway in L6 myotubes and skeletal muscle of C57BL/6J mice.
Materials and Methods
Preparation of standardized K. parviflora extract
Rhizomes of K. parviflora were collected from a traditional market in Bangkok, Thailand. The dried rhizomes were crushed and extracted with 95% ethanol in a shaking water bath at 60°C for 3 h. Then, the extract was concentrated using a vacuum rotary evaporator (Laborata 4000 Efficient; Heidolph Instruments GmbH & Co. KG., Schwabach, Germany) to obtain the K. parviflora extract (KPE) with a yield of 8.9% (w/w). To standardize KPE preparation, the identification and quantification of 5,7-dimethoxyflavone (DMF) in KPE were performed by high-performance liquid chromatography (HPLC). HPLC was carried out using the YL9100 HPLC system (Younglin, Anyang, Korea) equipped with a Sunfire C18 column (150 mm × 4.6 mm id, 5 μm; Waters, Milford, MA, USA). The mobile phases consisted of methanol (phase A) and water (phase B) with a flow rate of 0.5 mL/min, and was programmed as follows: 0–10 min, 35–37% B; 10–20 min, 37–39% B; 20–30 min, 39–41% B; 30–40 min, 41–43% B; 40–50 min, 43–45% B. The detection wavelength was 256 nm and injection volume was 20 μL. Chromatographic peaks of KPE were identified as comparing retention times of pure DMF (Sigma-Aldrich, St Louis, MO, USA). Quantitative analysis was calculated based on the standard curve using a pure DMF. The standardized KPE contained 14.1% (w/w) DMF as a bioactive compound. 11
Chemical reagents
Fetal bovine serum (FBS) and Dulbecco's modified Eagle's medium (DMEM) were supplied by Hyclone Laboratories, Inc. (Logan, UT, USA). Protease inhibitor cocktail were purchased from Sigma-Aldrich. Primary antibodies against phosphorylated SIRT1, AMPK (p-AMPK), total AMPK (t-AMPK), and α-tubulin were supplied by Cell Signaling Technology (Beverly, MA, USA). Primary antibodies against PGC-1α and PPARδ were supplied by Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Cell culture and myogenic differentiation
L6 rat myoblast and COS-7 monkey kidney cells (American Type Culture Collection, Manassas, VA, USA) were cultured in DMEM containing 1% antibiotics (100 units/mL penicillin A and 100 μg/mL streptomycin) and 10% FBS, under a humidified atmosphere (5% CO2) at 37°C. Differentiation into L6 myotubes was induced by changing the medium to DMEM containing 2% FBS. The differentiation medium, containing the different KPE concentrations (1 and 10 μg/mL) was refreshed every 2 days until day 6.
Measurement of mitochondrial content
After differentiation was induced, the L6 cells were washed with Dulbecco's phosphate-buffered saline (DPBS) and incubated with 50 nM MitoTracker® Green FM (Invitrogen, Carlsbad, CA, USA) for 40 min. The mitochondrial contents were quantified by measuring fluorescence intensity (excitation/emission at 490/516 nm) using a GloMax®-Multi Detection System (Promega, Madison, WI, USA).
Luciferase reporter gene assay
To assess the PGC-1α and PPARδ binding activities of KPE (1 and 10 μg/mL), COS-7 cells (4 × 106 cells/plate) were transfected in 24-well plates with the luciferase reporter plasmid pGL3-PGC-1α-luc (Addgene, Cambridge, MA, USA) and pFA-PPARδ-luc (Stratagene, La Jolla, CA, USA) using the transfection reagent Lipofectamine (Invitrogen), as described previously. 12 Briefly, after transfection for 24 h, the cells were treated with KPE. Transcriptional activities were evaluated by using luciferase assay substrate (Promega) with a MicroLumatPlus LB 96V luminometer (Berthold, Wildbad, Germany).
Reverse transcription–polymerase chain reaction
Total RNA from L6 cells and homogenized soleus muscle tissues were analyzed by reverse transcription–polymerase chain reaction (RT-PCR), according to our previous method. 12 Table 1 shows the primer pairs (Bioneer, Daejeon, Korea) used. PCR amplification was conducted in a GeneAmp PCR System 2700 (Applied Biosystems, Foster City, CA, USA). The PCR products were analyzed by 1.5% agarose gel electrophoresis (110 V/30 min), detected with the G:BOX EF imaging system, and quantified with the Gene Snap program (Syngene, Cambridge, United Kingdom).
ERRα, estrogen-related receptor-α; mtDNA, mitochondrial DNA; NRF-1, nuclear respiratory factor 1; PGC-1α, peroxisome proliferator-activated receptor-γ coactivator-1α; Tfam, mitochondrial transcription factor A.
PGC-1α knockdown
PGC-1α knockdown by siRNA transfection was conducted according to our previous method. 12 L6 myoblast cells were transfected with siRNA-targeting PGC-1α (si-PGC-1α) or negative control siRNA (si-NC; Bioneer) using Lipofectamine RNAiMAX transfection reagent (Invitrogen). The transfected myoblasts were differentiated into myotubes with differentiation medium in the presence of KPE.
Western blot analysis
The concentrations of lysate protein from L6 cells and homogenized soleus muscle tissue by NP-40 (ELPIS-Biotech, Daejeon, Korea) supplemented with a protease inhibitor cocktail were quantified using the Bradford assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The protein was analyzed by western blot assay, according to our previous method. 12 Primary antibodies (1:1000 dilution) against SIRT1, p-AMPK, t-AMPK, PPARδ, PGC-1α, and α-tubulin were used. The membrane containing the bound primary antibodies was then incubated with goat anti-rabbit or goat anti-mouse horseradish peroxidase-conjugated IgG secondary antibodies (1:5000 dilution; Bethyl Laboratories, Inc., Montgomery, TX, USA). Antibody signals were visualized by the enhanced chemiluminescence detection system (Amersham Biosciences, Little Chalfont, United Kingdom) and scanned with the G:BOX EF imaging system (Syngene).
Animal experiments
Four-week-old male C57BL/6J mice (Orient Bio, Inc., Seongnam, Korea) were maintained in the Yonsei Laboratory Animal Research Center (Seoul, Korea) at 25°C ± 2°C and 55% ± 5% relative humidity with a 12 h light-dark cycle and were freely provided with food and tap water during the experimental period. The mice were separately fed a normal diet (ND, 10% kcal fat, 38057; Cargill Agri Purina, Seongnam, Korea) or high-fat diet (HFD, 45% kcal fat, D12451; Research Diets, Inc., New Brunswick, NJ, USA) for 8 weeks. Then, the mice were randomly assigned to four groups as follows: group 1 were fed the ND; group 2 were fed the ND with KPE at 200 mg/kg/day (ND+KPE); group 3 were fed the HFD; and group 4 were fed the HFD with KPE at 200 mg/kg/day (HFD+KPE), respectively. Animals were orally administrated with KPE for 10 weeks. After sacrificing the mice, the soleus and gastrocnemius muscles were separated and stored at −70°C. The protocol for the animal experiment was approved by the Institutional Animal Care and Use Committee (IACUC) of Yonsei University (Permit No. IACUC-2013-0102).
Exercise performance test
The exercise performance was tested to define the running exercise capacity. A variable speed belt treadmill (Columbus Instruments, Columbus, OH, USA), enclosed in a plexiglass chamber, with a stimulus device, consisting of a shock grid attached to the rear of the belt (Panlab, Barcelona, Spain) was used. Animals were acclimated to the test conditions using a habituation protocol before the running test. The mice were run at a speed 16.2 m/min for 10 min with a 5° inclination. For the real running test, the experiments started at a speed 10.8 m/min with the treadmill set at 5° inclination. The speed gradually increased from 10.8 to 24.6 m/min and then maintained constant until the mice reached exhaustion. Exhaustion was defined as when the mice were unable to run for 10 sec despite the electric shock. The electric shock grid was set to deliver 0.2 mA, which caused uncomfortable shock but did not physically injure the animal.
Micro-computed tomography imaging
Micro-computed tomography (micro-CT) experiments were conducted with an animal positron emission tomography/CT/single photon emission tomography system (Inveon, Siemens, Washington, DC, USA) at the Pohang Center for Evaluation of Biomaterials, Pohang Technopark (Pohang, Korea).
Quantification of mitochondrial DNA content
The mitochondrial DNA (mtDNA) in the soleus muscle tissue of the mice was measured by RT-PCR, according to our previous method. 12 Table 1 shows the primer pairs (Bioneer) used. Results were represented as the relative number of mitochondrial genomes per diploid nuclei. The rate of mtDNA/genomic DNA was determined by measuring the relative density of the band.
Statistical analysis
Descriptive data were reported as the mean ± standard deviation of three independent experiments (in vitro) or of eight mice per group (in vivo). All statistical analyses were conducted with SPSS 21.0 software (SPSS, Inc., Chicago, IL, USA). One-way ANOVA (analysis of variance), followed by a Scheffe's test was used for the in vitro data. The differences between groups were evaluated by the unpaired Student's t-test for the in vivo data. P < .05 was considered statistically significant.
Results
KPE improves mitochondrial biogenesis by increasing PGC-1α activity in L6 myotubes
To evaluate the increase in PGC-1α activity by KPE, PGC-1α plasmid was transfected into COS-7 cells. As shown in Figure 1A, KPE dose-dependently increased PGC-1α binding activity by 182% and 313% at 1 and 10 μg/mL compared with the untreated cells, respectively. Also, the mitochondrial density was markedly increased by KPE in differentiated L6 myotubes (Fig. 1B), which indicated an increase in the mitochondrial mass of skeletal muscle. Moreover, KPE effectively upregulated the gene expression of PGC-1α, which leads to activation of mitochondrial biogenesis-related transcription factors including ERRα, NRF-1, and Tfam in L6 myotubes (Fig. 1C). Next, L6 myoblasts transfected with si-PGC-1α or si-NC were used to elucidate the role of KPE on PGC-1α expression associated with mitochondrial biogenesis. The gene expression of PGC-1α and its target gene of mitochondrial biogenesis transcription factors, NRF-1 and Tfam, were markedly downregulated by the si-PGC-1α, as compared to si-NC, in absence and presence of KPE. Moreover, KPE treatment increased the gene expression of mitochondrial biogenesis transcription factors of NRF-1 and Tfam, which were decreased by the si-PGC-1α (Fig. 1D). These results reveal that KPE promotes the biogenesis of mitochondria by increasing PGC-1α activity in L6 myotubes.

Effect of KPE on mitochondrial biogenesis in L6 myotubes. COS-7 cells transfected with each reporter plasmid were treated with the different KPE concentrations.
KPE enhances exercise performance and increases skeletal muscle weight in mice
To examine whether KPE could improve exercise performance, the running distance and time to exhaustion were measured by using a treadmill in ND mice and HFD obese mice. Both the running distance and time were significantly improved in KPE-treated ND mice, by 42.4% and 32.7%, respectively, compared to ND mice (Fig. 2A). Likewise, the running distance and time increased by 47.5% and 47.4%, respectively, in the KPE-treated HFD obese mice compared to HFD obese mice (Fig. 2B). Next, to further evaluate the effect of KPE on skeletal muscle weight, we quantified the hind limb skeletal muscle area using micro-CT analysis and measured the soleus and gastrocnemius muscles weights. Micro-CT analysis showed that the hind limb skeletal muscle area was markedly increased by KPE in both ND and HFD obese mice (Fig. 2C, D). Also, the ratio of skeletal muscle weight/body weight, expressed as the relative number of soleus and gastrocnemius muscles weights per body weight, was significantly increased, by 22% and 21%, respectively, in the KPE-treated ND and HFD obese mice compared to the corresponding nontreated mice (Fig. 2E, F). These data suggest that KPE effectively enhances exercise performance and increases skeletal muscle weight in nonobese and obese individuals.

Effect of KPE on exercise performance and skeletal muscle weight. After 8 weeks of dietary manipulation to induce obesity, mice were orally administrated ND, ND with KPE at a dose of 200 mg/kg/day (ND+KPE), HFD (HFD), and HFD with KPE at a dose of 200 mg/kg/day (HFD+KPE) for 10 weeks. Running distance and time were monitored by treadmill at the end of KPE treatment in
KPE activates mitochondrial biogenesis in skeletal muscle tissue of mice
To verify that the increase in exercise performance of KPE-treated mice accompanied an improvement in mitochondrial biogenesis, we evaluated the mitochondrial biogenesis transcription factors and mtDNA content in skeletal muscle tissue of KPE-administrated mice. The expression of ERRα, PGC-1α, Tfam, and NRF-1 involved in mitochondrial biogenesis was increased in the skeletal muscle tissue of KPE-treated ND mice and KPE-treated HFD obese mice (Fig. 3A). For both the ND and HFD obese mice treated by KPE, the mtDNA content also significantly increased, by 2.0- and 1.7-fold, respectively, compared to the nontreated ND and HFD obese mice (Fig. 3B). These data suggest that KPE enhances mitochondrial function and mass in both ND and HFD obese mice.

Effect of KPE on mitochondrial biogenesis in skeletal muscle tissue.
KPE stimulates SIRT1/AMPK/PGC-1α/PPARδ signaling pathway in L6 myotubes and mice
To confirm whether KPE activates exercise-mediated signaling pathway, the protein expressions of SIRT1, AMPK, PGC-1α, and PPARδ were evaluated in both the L6 myotubes and skeletal muscle tissue of the mice. KPE effectively activated the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway by increasing the protein levels of SIRT1, phosphorylated AMPK, PGC-1α, and PPARδ in L6 myotubes and skeletal muscle tissue of mice (Fig. 4A, B). Moreover, KPE significantly increased the PPARδ binding activity in COS-7 cells (Fig. 4C). These data suggest that KPE can trigger signaling cascade that regulates energy metabolism and is responsible for the stimulation of the SIRT1/AMPK/PGC-1α/PPARδ signaling cascade.
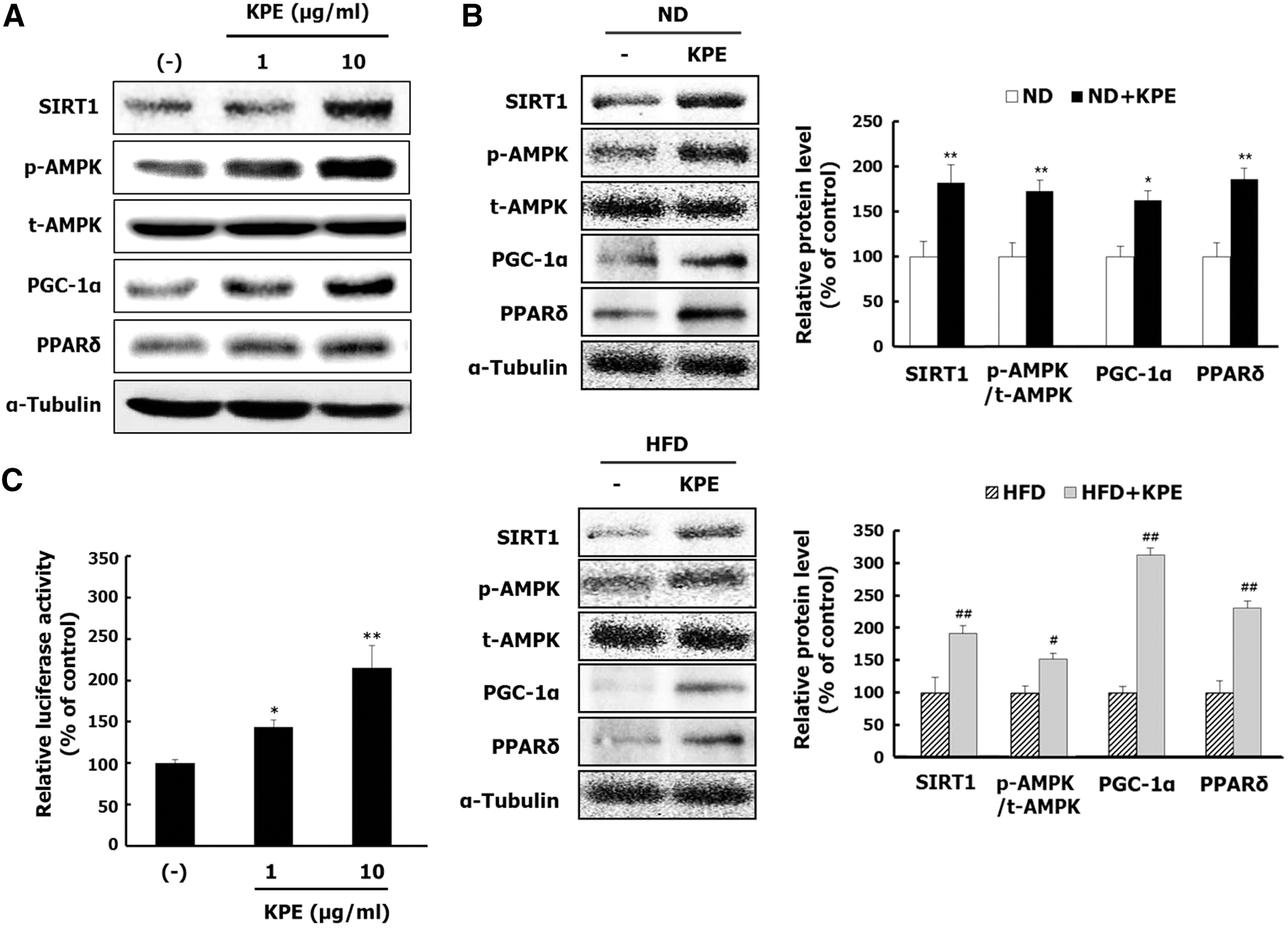
Effect of KPE on the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway in vitro and in vivo. L6 cells were cultured with the differentiation medium in the different KPE concentrations for 6 days. The protein levels of SIRT1, AMPK, PGC-1α, and PPARδ
Discussion
Natural exercise mimetics focus on increasing the exercise performance for nonobese, and obese, individuals. 13 Increased physical activity contributes to improving metabolic homeostasis and energy expenditure, which can result in lowering the relative risk of obesity-related metabolic diseases. 13 According to a previous study, KPE enhances physical fitness performance and muscle endurance by improving energy metabolism in normal condition ddY mice. 14 Furthermore, in clinical human studies, KPE was shown to enhance the muscle strength of soccer players and improve physical fitness performance of the elderly, through increasing serum antioxidant enzyme activities. 9,15 However, there are few reports evaluating the effect of KPE on improving physical activity in an obese model. This study investigated whether KPE could increase exercise performance in obese and nonobese mice by stimulating the mitochondrial biogenesis and the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway.
PGC-1α controls mitochondrial biogenesis and changes in muscle fiber characteristics by orchestrating several key transcription factors including ERRα, Tfam, and NRF-1. 16 KPE increased both the activity and the expression of PGC-1α, which directly elevated mitochondrial density and upregulated the expression of mitochondrial transcription factors in L6 myotubes (Fig. 1). Binding of PGC-1α to NRF-1 enhances the expression of Tfam responsible for mtDNA transcription, replication, and maintenance. ERRα has been linked to the regulation of oxidative metabolism and is a cofactor for PGC-1α. 12 The increase in PGC-1α activity by KPE is required to improve mitochondrial function and mass, which is a hallmark of exercise-induced changes. 1 Previously, phytochemical compounds, such as resveratrol, 5 quercetin, 17 and chitooligosaccharide 18 were shown to enhance exercise performance by elevating skeletal muscle mitochondrial function.
Exercise performance is closely associated with oxidative metabolism of skeletal muscle by mediating mitochondrial biogenesis. 19 Abnormalities of energy metabolism in skeletal muscle can lead to lowered exercise performance, depending on the mitochondrial oxidative function of skeletal muscle. 19 KPE extended the running capacity and increased skeletal muscle weight in ND and HFD obese mice, by improving mitochondrial biogenesis (Fig. 2). This result supported that KPE activated mitochondrial biogenesis transcriptional factors and increased mtDNA numbers in skeletal muscle tissue in the ND and HFD obese mice (Fig. 3). A high content of oxidative (slow-twitch) fibers enhances skeletal muscle function to prevent muscle wasting and leads to increased skeletal muscle weight. 8 The increased exercise performance of KPE-treated mice may stimulate the muscle fiber type transformation from fast- to slow-twitch fiber, enhancing the gene level of mitochondrial biogenesis regulatory factors and increasing the skeletal muscle weight. Furthermore, HFD obese mice show a low exercise performance, increase oxidative stress, impair mitochondrial metabolism, and disturb the biogenesis of mitochondria in skeletal muscle. 19 KPE improved the exercise performance of HFD obese mice through activation of mitochondrial biogenesis, supporting that KPE prevented mitochondrial dysfunction by reducing obesity-induced oxidative stress. Thus, the increased exercise performance and skeletal muscle weight by KPE is dependent on the stimulated mitochondrial biogenesis, suggesting that KPE can improve exercise-induced mitochondrial biogenesis in skeletal muscle.
The exercise-mediated signal cascades including SIRT1, AMPK, PGC-1α, and PPARδ stimulate exercise performance, without exercise, and develop oxidative myofibers. 8 SIRT1 and AMPK regulate the activities of both PGC-1α and PPARδ that control energy expenditure and mitochondrial function in response to exercise. 5,20 KPE upregulated the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway (Fig. 4), supporting that KPE can increase the exercise capacity of mice. Particularly, the upregulation of PPARδ in KPE-treated mice may promote the formation of oxidative slow-twitch muscle fibers, leading to increased skeletal muscle weight. Thus, these results indicate that the exercise performance and skeletal muscle weight are dependent on activation of an exercise metabolism signaling pathway, such as the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway in the skeletal muscle of mice.
Collectively, KPE enhanced endurance exercise capacity by promoting mitochondrial biogenesis and the SIRT1/AMPK/PGC-1α/PPARδ signaling pathway. KPE serves as an exercise mimetic and supplement based on its ability to improve exercise capacity. Clinical trials in nonobese and obese individuals are needed to confirm the enhancement effect of KPE on exercise performance in humans.
Footnotes
Acknowledgments
This research was supported, in part, by the Commercializations Promotion Agency for R&D Outcomes (COMPA), funded by the Ministry of Science, ICT and Future Planning (MSIP) (2016K000107).
Author Disclosure Statement
No competing financial interests exist.