
Abstract
Cancer stem cells, a small subpopulation of cells with stem cell-like characteristics found within most solid tumors, are widely reported to be responsible for the malignancy of aggressive cancer cells, and targeting these cells presents a sound therapeutic strategy for reducing the risk of tumor relapse. In the present study, we examined the effects of an extract of Saccharina japonica (ESJ) on glioblastoma stem cells (GSCs). Saccharina japonica is a member of the Phaeophyceae (brown algae) family, which displays biological activities, including antitumor effects. ESJ inhibited the sphere-forming ability of GSCs in vitro as evidenced by neurosphere formation and limiting dilution assays. Treatment with ESJ partially induced apoptosis, reduced cell invasiveness, and sensitized GSCs to ionizing radiation. In addition, ESJ inhibited the maintenance of stemness in GSCs by suppressing the expression of epidermal growth factor receptor (EGFR)/EGFR variant III (EGFRvIII) and Notch intracellular domain. Intriguingly, the observed ESJ-induced suppression also appeared to induce the proteasomal degradation of EGFR/EGFRvIII. Our results indicate that ESJ could be considered a potent therapeutic adjuvant that targets GSCs.
Introduction
G
It is widely accepted that one of the main causes of therapeutic failure is the presence of stem-like cancer cells called glioblastoma stem cells (GSCs), which reside in GBM. 4 GSCs possess characteristics similar to those of neural stem cells, but with tumorigenic potential. 5,6 GSCs are known to contribute to the therapeutic resistance of GBM tumors and promote their initiation, progression, and recurrence. 7,8 In general, while non-GSCs can be eliminated by conventional therapies, including drugs and radiation, GSCs resist therapy and promote recurrence in the tumor microenvironment. 9 –12
Recently, the potential use of natural products in the field of cancer chemoprevention has gained attention. 13,14 Natural products are a rich source of bioactive compounds and are comparatively less toxic than existing chemotherapeutic drugs. 13,14 Our previous study suggested that emodin isolated from Rheum palmatum was an effective inhibitor of sphere-forming ability and stemness signaling and was able to induce cell death in GSCs in vitro. 15 Therefore, the development of natural products targeting GSCs could be considered as an alternative strategy to deal with the devastating effects of cancer.
Saccharina japonica, a member of the Phaeophyceae (brown algae) family, is generally produced in Asian countries, including China, Japan, and Korea. It has a long history of use not only as a good source of food but also as an oriental medicine drug to cure many diseases. 13,14 A variety of polysaccharides, which are abundant in brown algae, such as laminarans, alginic acids, and sulfated polysaccharides (fucoidans), 16 have demonstrated anticancer effects in colon cancer and breast cancer. 17 –21 Therefore, in the present study, we investigated the effect of an extract of S. japonica (ESJ) on GSCs.
Materials and Methods
Preparation of extract from S. japonica
The brown alga S. japonica was collected at Daegin harbor, Gangwon province, Korea, and kept in a deep freezer at −70°C. The specimens (wet weight 1 kg) were chopped and then immersed in absolute methanol (3 L) with mechanical stirring for 1 day at room temperature. The methanol extracts were evaporated under vacuum at 40°C and then reextracted with ethyl acetate. Final crude extract (1.8 g) was obtained by evaporating the ethyl acetate fraction. ESJ was dissolved in dimethyl sulfoxide (DMSO) at 100 mg/mL concentration (stock solution) and kept at −20°C. ESJ stock solution was diluted to working concentration (10 mg/mL) with DMSO and kept at 4°C during the experiments.
Reagents
Epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) were obtained from R&D Systems. Notch-1 and Nanog antibodies were obtained from Millipore. Nestin, Oct4, CD133, C-Myc, ZEB1, Twist, and β-actin were purchased from Santa Cruz Biotechnology. Phospho-AKT [p-AKT (T308 and S473)], p-STAT3 (Y705), p-ERK1/2, nonphosphorylated-β-catenin (n-p-β-catenin), Musashi, SOX2, Notch-1, Snail, and Slug were obtained from Cell Signaling Biotechnology. Anti-EGF receptor (EGFR) was obtained from BD Sciences (Becton Dickinson). Fibronectin was obtained from Sigma-Aldrich. Vimentin was purchased from Thermo Fisher Scientific.
Cell culture
The human GSCs were generously gifted by Dr. Soeda (X01) 22 (Gifu University School of Medicine, Gifu, Japan), Dr. Kang (0315) 23 (Yonsei University, Seoul, Korea), and Dr. Nakano (83 and MD30) 24 (Ohio State University, Columbus, OH, USA). GSCs were cultured in a floating sphere state in Dulbecco's modified Eagle medium (DMEM)/F12 medium (Corning) supplemented with 20 ng/mL of EGF, 10 ng/mL of bFGF, 0.1% B27 (GIBCO), and 1% antibiotic–antimycotic solution (Welgene) (complete medium) and were maintained in a humidified incubator at 37°C under an atmosphere of 5% CO2.
Sphere formation and limiting dilution assay
GSCs were dissociated with accutase (MILLIPORE) into single cells and resuspended in complete medium. For sphere formation, single GSCs were seeded in a 6-well plate containing 2 mL of complete media per well at 1 × 103 cells/mL. Cells were treated with or without indicated concentrations of ESJ in the figures. After 7 days of incubation, spheres were attached to the plate by adding 10% fetal bovine serum (FBS) overnight, stained with 0.05% crystal violet dissolved in 20% methanol, and counted.
For limiting dilution assay, single GSCs were seeded with decreasing numbers of cells (100, 50, 25, 12, 6, and 3) per well in 96-well plates in 100 μL of complete medium. After 7 days of incubation, wells without spheres were counted and plotted. P-values were calculated using software. *
Western blot analysis
GSCs pretreated with ESJ were lysed in RIPA buffer [50 mM Tris-HCl (pH 7.4), 100 mM NaCl, 5 mM EDTA, 0.5% Nonidet P-40, phosphatase inhibitor cocktail set II (Calbiochem), and protease inhibitor cocktail tablet (Roche)]. The protein contents were determined by the Bio-Rad protein assay reagent (Bio-Rad). The proteins were separated using 8–12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The membranes were blocked with 5% skim milk in TBST for 1 h at room temperature and then incubated with the indicated primary antibodies overnight at 4°C with gentle shaking. After extensive washing with TBST, the membrane was developed with a peroxidase-conjugated secondary antibody for 2 h at room temperature. After washing thrice with TBST for 10 mins, membranes were visualized by enhanced chemiluminescence (Amersham) according to the manufacturer's protocol.
Irradiation and drug treatment
For measuring ionizing radiation (IR) resistance, GSCs were seeded in 6-well plates at 1 × 103 cells/mL. Cells were pretreated with or without ESJ (2 μg/mL) for 1 h and then exposed to γ-rays from a 137Cs γ-ray source (BIOBEAM8000, 2.6 Gy/min, Gamma-service Medical GmbH) at the indicated dose rate. After 7 days of incubation, spheres were attached by adding 10% FBS, washed with phosphate buffered saline (PBS), and stained with 0.05% crystal violet dissolved in 20% methanol. After washing thrice with distilled water, colonies were counted.
Matrigel™-coated transwell invasion assay
Invasion assays were performed using Transwell chambers with the polycarbonate nucleopore membrane (6.5 mm in diameter, 8-μm pore size) (Corning). The upper part of the nucleopore membrane was covered with growth factor-reduced Matrigel (100 μg/cm2) (Corning) and coated by drying the Matrigel solution in the clean bench for 1 h. Cells were pretreated with or without ESJ for 72 h before invasion assays. Cells (1 × 105) suspended in 100 μL of DMEM/F12 were seeded into the upper chamber. The lower compartments were filled with 600 μL of the medium with EGF (20 ng/mL). After incubation at 37°C for 18 h, remaining cells on the upper surface of the filters were wiped out with a cotton swab, and the invaded cells on the lower surface were fixed and stained with the Diff-Quik solution I and II, respectively (Sysmex). Invasiveness was determined by counting cells in three microscopic fields per well, and the extent of invasion was expressed as the average number of cells per microscopic field.
Fluorescence-activated cell sorting analysis
To examine cell cycle analysis by flow cytometry, GSCs were pretreated with ESJ, harvested at indicated times, and dissociated into single cells. Cells were fixed with 75% ethanol, incubated at −20°C for at least 1 h, and washed with PBS. Next, cells were resuspended in PBS with 0.1 mg/mL RNase A, 50 mg/mL propidium iodide (PI), and 0.05% Triton X-100 for 40 mins at room temperature, washed with PBS, and subjected to flow cytometric analysis.
To observe ESJ-induced cell death by flow cytometry, GSCs were measured by Annexin-V and PI staining, according to the manufacturer's protocol. After ESJ treatment, cells were harvested at indicated times and dissociated into single cells. Cells were incubated in binding buffer containing Annexin-V and PI in the dark for 15 min at room temperature. The stained samples were analyzed on an FACSCalibur (Becton Dickinson) within 1 h. All experiments were performed in triplicate.
Reverse transcriptase–polymerase chain reaction
High-quality total RNA was isolated from GSCs by using TRIsure (BIOLINE) as described in the manufacturer's protocol. Complimentary DNA was synthesized using the cDNA synthesis kit (BIOLINE). Reverse transcriptase–polymerase chain reaction (RT-PCR) for EGFR (1044-bp) and EGFR variant III (EGFRvIII) (243-bp products) was performed, as described previously. 22 Oligonucleotide primer sequences were as follows: EGFR/EGFRvIII forward: 5′ CTT CGG GGA GCA GCG ATG CGA C 3′ and EGFR/EGFRvIII reverse: 5′ ACC AAT ACC TAT TCC GTT ACA C 3′; and 18S RNA forward primer 5′-AAACGGCTACCACATCCAAG-3′ and reverse 5′-CGCTCCCAAGATCCAACTAC-3′.
Statistical analysis
Data are presented as mean ± SEM of at least three independent experiments. Significant differences were analyzed with Student's t-test and were considered at *P < 0.05, **P < 0.01, and ***P < 0.001. Statistical analysis and graphing were performed using Microsoft Excel 2013 and GraphPad Prism 6.0 software.
Results
ESJ inhibits sphere-forming activity of GSCs
First, we examined whether ESJ affected the sphere-forming ability of GSCs. Treatment with ESJ suppressed the sphere-forming ability in four different GSC lines in a dose-dependent manner (Fig. 1A). We observed that treatment with 5 μg/mL of ESJ almost completely inhibited the sphere-forming ability in GSC lines. To confirm the effect of ESJ on GSCs, we performed a limiting dilution assay, which is an alternative way to assess sphere-forming ability activity in vitro. As shown in Figure 1B, pretreatment with ESJ significantly inhibited the sphere-forming ability of GSCs in a dose-dependent manner. These results suggest that ESJ effectively suppresses the sphere-forming property of GSCs. To confirm the use of primary GSCs in this study, we checked the expressions of stem cell and differentiation markers in stem cell culture condition and differentiation condition, respectively (Supplementary Data and Supplementary Fig. S1; Supplementary Data are available online at
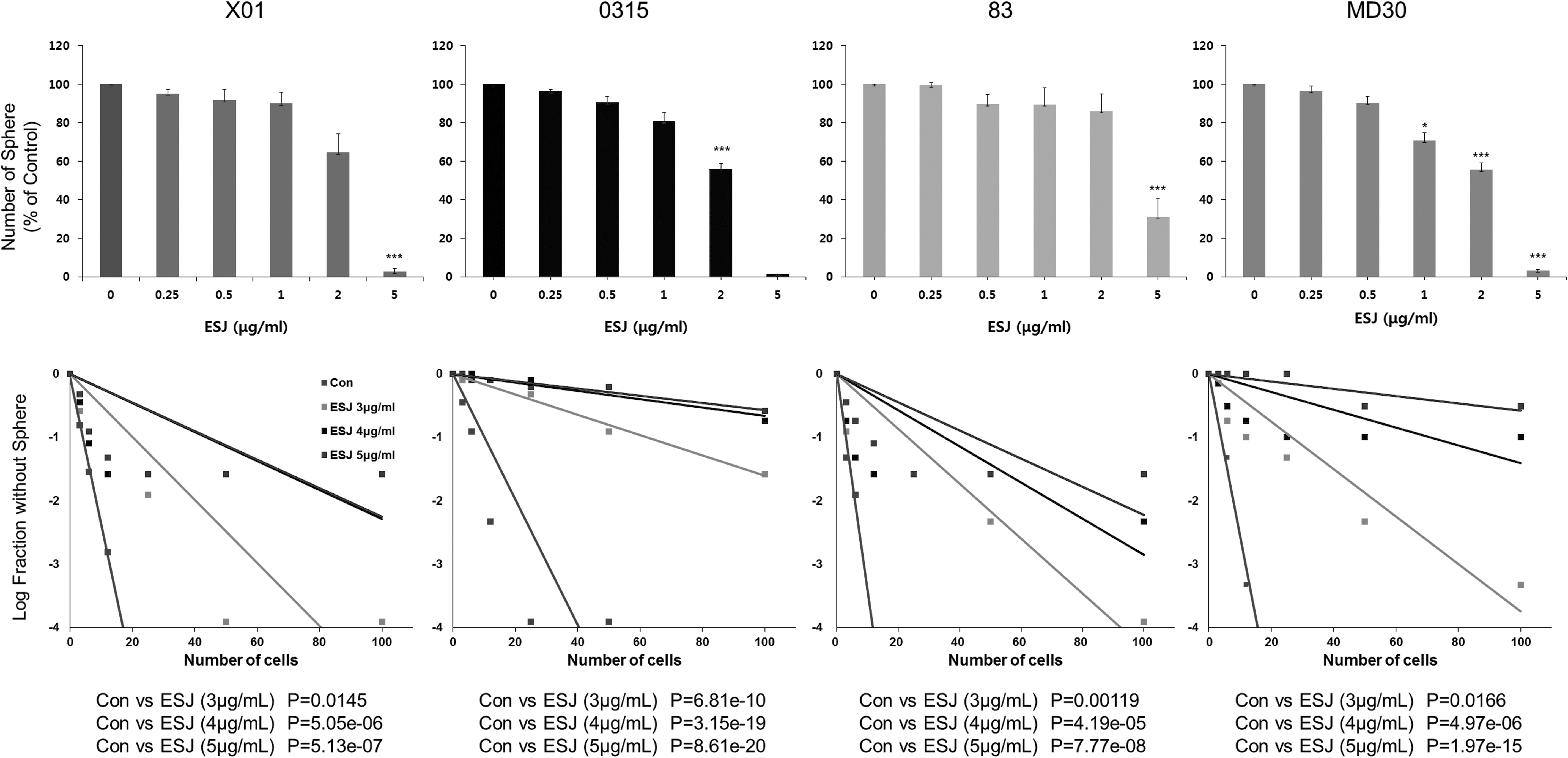
Effect of ESJ on self-renewal activity of GSCs.
ESJ inhibits stemness signaling of GSCs by proteasomal degradation of EGFR/EGFRvIII
Next, we attempted to determine the molecular mechanism responsible for ESJ-induced suppression of the sphere-forming capacity of GSCs. As it has been reported that ESJ suppresses EGFR signaling and overexpression of EGFR/EGFRvIII is a common feature of GBM, 25 we examined whether ESJ affects EGFR/EGFRvIII-mediated stemness signaling in GSCs. As shown in Figure 2A, ESJ suppressed the expression of EGFR/EGFRvIII in GSCs.

Effect of ESJ on the EGFR/EGFRvIII expression and associated signaling pathways in GSCs.

Intriguingly, we found a slow-migrating EGFR/EGFRvIII band in the Western blot analysis, a phenomenon we previously observed when studying ubiquitin-mediated proteasomal degradation of EGFR/EGFRvIII by emodin in GSCs. 15 Consistent with our previous finding, RT-PCR analysis determined that ESJ did not affect the transcription level of EGFR/EGFRvIII (Fig. 2B). To verify the mechanism of EGFR/EGFRvIII degradation by ESJ, we pretreated GSCs with MG132, an inhibitor of proteasome-dependent degradation. As expected, MG132 treatment interrupted the ESJ-induced degradation of EGFR/EGFRvIII in GSCs (Fig. 2C). Since suppression of EGFR/EGFRvIII signaling resulted in downregulation of stemness-associated genes, 15 we also examined the changes by Western blot analysis. We found that ESJ treatment of GSCs decreased the expression of stemness-related signaling molecules, including Notch, n-p-β-catenin, and Sox2, in a dose- and time-dependent manner (Fig. 2D). These data suggest that ESJ induces proteasomal degradation of EGFR/EGFRvIII, followed by suppression of the Notch, β-catenin, and STAT3 signaling pathways that are responsible for the sphere-forming capacity of GSCs.
ESJ leads to partial cell death in GSCs
We examined the effect of ESJ on cell cycle progression in GSCs. ESJ treatment did not significantly change the overall cell cycle, but after a 72-h treatment, it induced a slight but significant increase of the sub-G1 fraction (Fig. 3A, C).
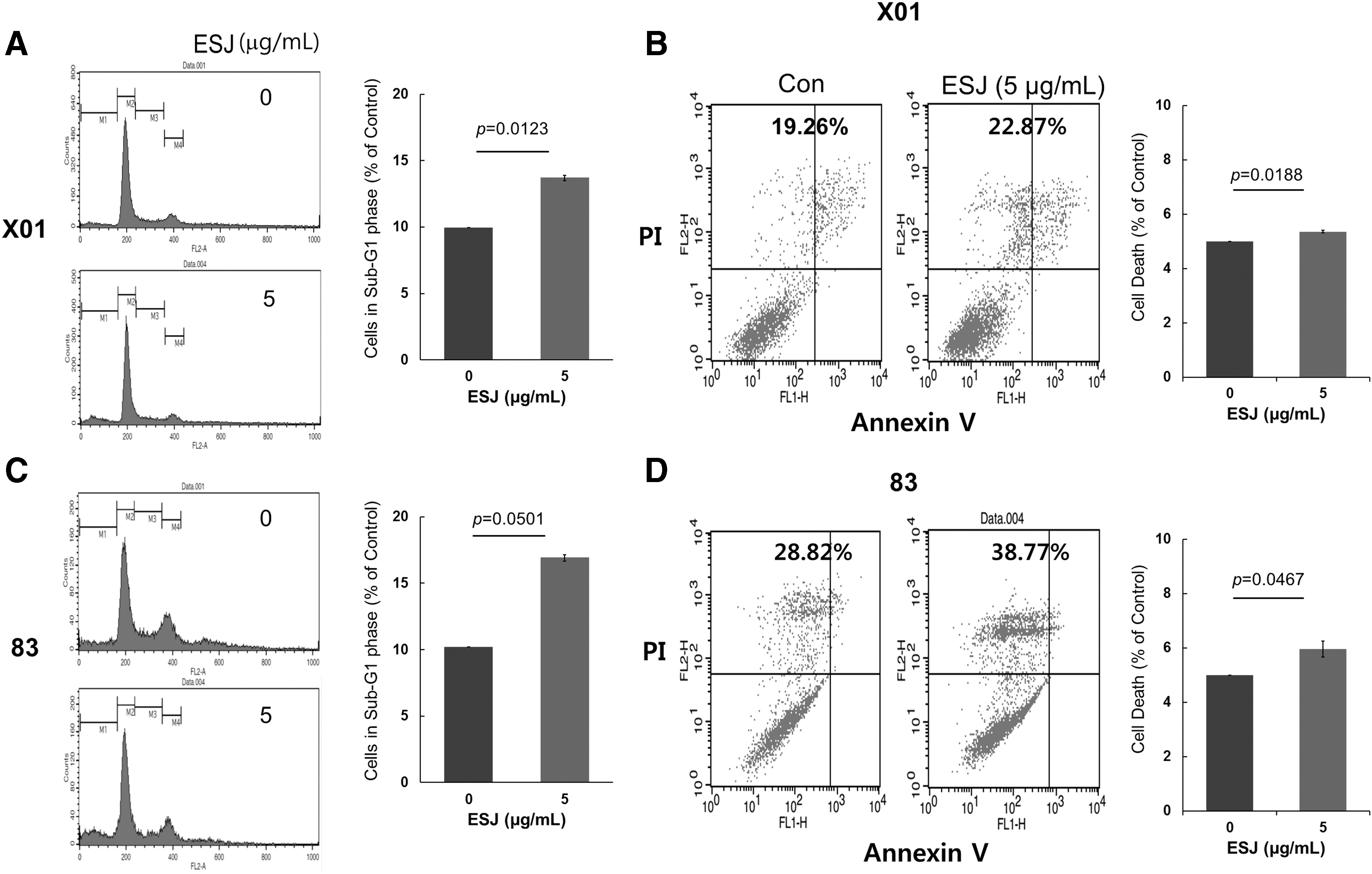
Effect of ESJ on cell death of GSCs. Evaluation of cell death by cell cycle analysis and Annexin-V and PI staining, followed by flow cytometry analysis.
As the population of the sub-G1 fraction of the cell cycle is indicative of increased cell death, we performed an Annexin-V/PI staining assay. The analysis showed that ESJ slightly, but significantly, induced cell death (Fig. 3B, D). These data indicate that ESJ has the potential to induce partial cell death in GSCs.
To examine the effect of ESJ on normal cell damage, we treated normal human astrocytes (NHAs) with ESJ. As shown in Supplementary Figure S3 (Supplementary Data are available online at
ESJ suppresses invasion of GSCs
In this study, we examined the effect of ESJ on the invasion activity of GSCs. Invasion is one of the hallmarks of cancer 26 and causes malignant glioma to invade the surrounding normal brain tissue. 27 Pretreatment of GSCs with ESJ almost completely suppressed the invasion of GSCs through a Matrigel-coated Transwell (Fig. 4A). Additionally, the expression of Snail and Slug, mesenchymal markers that are known to be related to the invasion of glioma cells, 28,29 was downregulated in a time-dependent manner (Fig. 4B). Other mesenchymal markers, including fibronectin, vimentin, Twist, and Zeb1, were changed marginally by ESJ treatment. Therefore, ESJ effectively blocked the invasion of GSCs, possibly through downregulation of the mesenchymal markers, Snail and Slug.
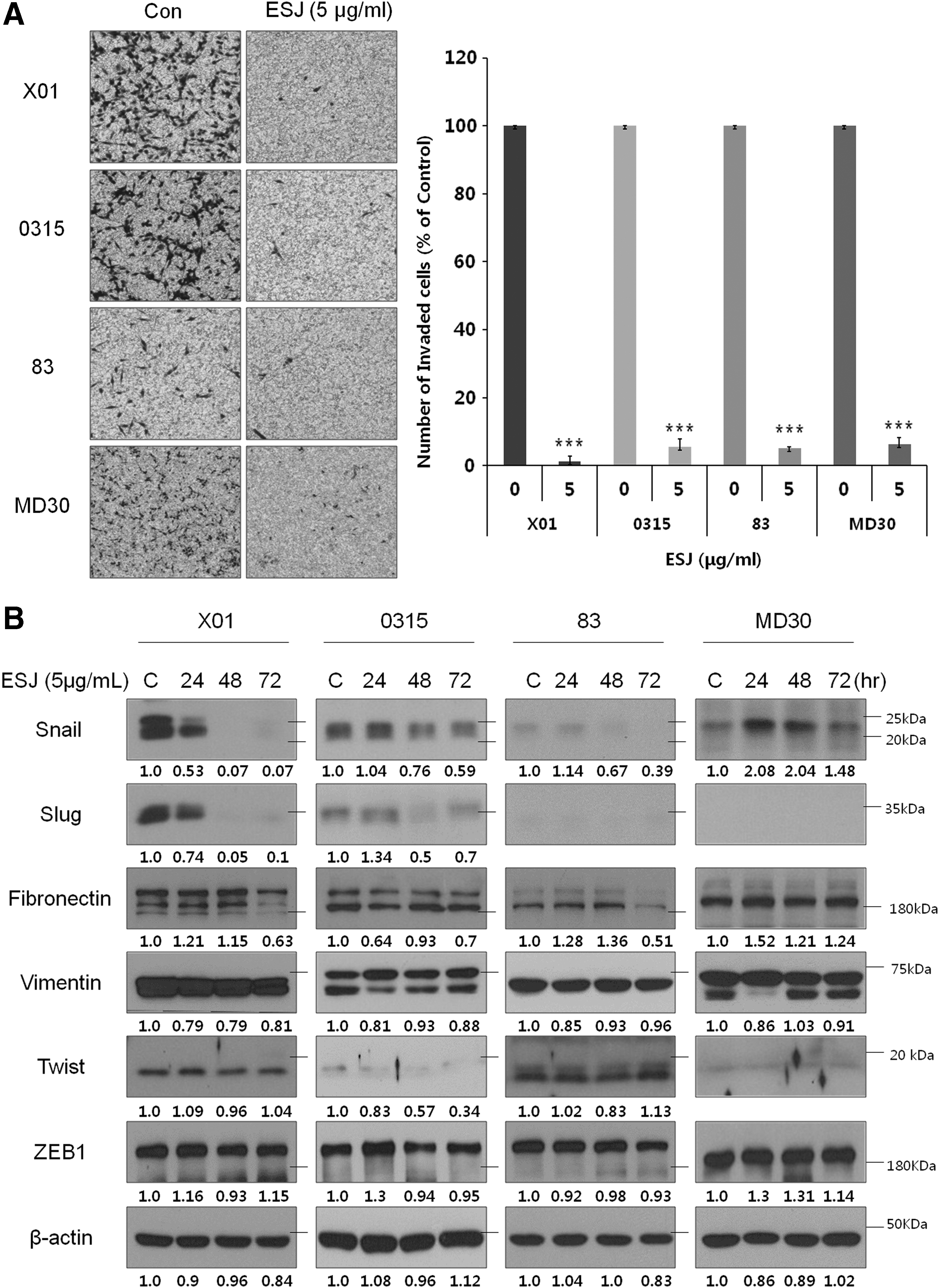
Effect of ESJ on the invasiveness of GSCs.
ESJ sensitizes GSCs to IR
GSCs are generally resistant to IR, but can become sensitized to it by a Notch blockade, using pharmacologic inhibitors of γ-secretase, or Notch shRNA. 9,30 As our data showed that ESJ downregulated Notch expression (Fig. 2D), we examined whether ESJ sensitizes GSCs to IR. Pretreatment of GSCs with 2 μg/mL ESJ, at which concentration it causes 25% inhibition in the rate of sphere formation, led to an increase in IR sensitivity in all four GSC lines (Fig. 5).
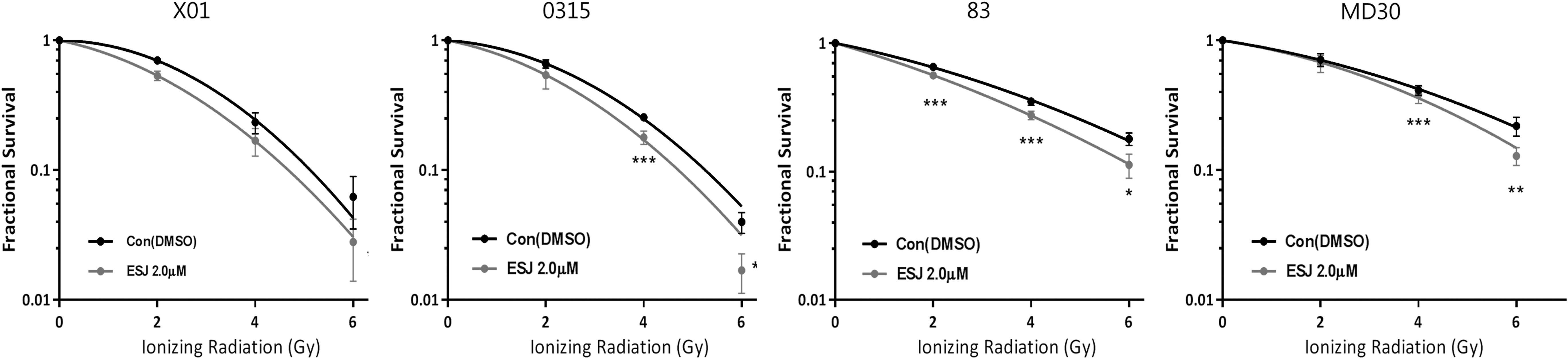
Effect of ESJ on the radiosensitivity of GSCs. Sphere-forming survival assays of GSCs at indicated doses of ionizing radiation in the Figure with or without 2 μg/mL ESJ. After 7 days of incubation, spheres were attached to the plate by adding 10% of FBS and stained with 0.05% of crystal violet. Attached spheres were counted. Values represent mean ± SEM. *P < 0.05, **P < 0.01, ***P < .001. DMSO, dimethyl sulfoxide.
Overall, our data strongly suggest that ESJ effectively suppresses the clonogenic activity of GSCs, by inducing degradation of EGFR/EGFRvIII and suppression of downstream signaling pathways, and sensitizes the cells to IR. Therefore, ESJ might be an effective adjuvant agent for combination therapy with IR to target GSCs.
Discussion
Glioblastoma, one of the most lethal cancers, is extremely resistant to conventional radio- and chemotherapies, with incurable recurrence often arising following surgery. 3 Many studies have shown that the malignant traits of cancer, including therapeutic resistance, severe tumor angiogenesis, invasion, metastasis, and recurrence, are caused by the presence of cancer stem cells (CSCs), a small subpopulation of cells with stem-like properties. 7,8 Therefore, research on therapies targeting CSCs will be essential for the development of new strategies against malignant cancers. Possible sources of CSC-targeting agents are naturally occurring dietary compounds; it has been observed that many natural compounds exhibit anticancer effects such as cancer growth inhibition, self-renewal, and promotion of apoptosis signaling in various types of cancer cells and CSCs. 13 –17,19,21,31 –35
In the present study, we investigated the effect of ESJ, found in brown algae, on the biology of GSCs. We found that ESJ inhibited the sphere-forming capacity of GSCs and suppressed the stemness signaling pathways, such as Notch, β-catenin, and STAT3 pathways, by inducing the degradation of EGFR/EGFRvIII. In addition, ESJ partially induced cell death, significantly inhibited invasion, and increased the sensitivity to IR in GSCs. Based on these results, it is possible that ESJ contains components that may be effective in targeting GSCs in combination with IR therapy.
Over the past 50 years, natural marine compounds have become increasingly popular as anticancer agents. In particular, several reports have shown that natural products from marine algae exhibit antioxidative, anti-inflammatory, and anticancer activities. 36 The anticancer activity of brown alga, S. Japonica, has been well established. The compounds best known for their biological activities against cancer are polysaccharides named fucoidans. Indeed, polysaccharides extracted from ESJ have been reported to induce apoptosis in various cancer cell lines 19,37,38 and suppress tumor growth and metastasis in vivo. 39 –41 In this study, we investigated the effect of ESJ on the biology of patient-derived GSCs. We found that ESJ significantly suppressed the sphere-forming activity of GSCs, possibly by inducing proteasomal degradation of EGFR/EGFRvIII, followed by suppression of downstream stemness signaling pathways, including Notch, n-p-β-catenin, and STAT3 pathways, in GSCs. Further studies will be required to identify which of the active ingredients of ESJ are responsible for inhibition of GSC sphere-forming activities.
EGFR amplification and the presence of its deletion mutant EGFRvIII are common features of malignant GBM. 43 Specifically, EGFRvIII is responsible for the maintenance of stemness, therapeutic resistance, and tumor progression in GBM cells. 42,43 In our previous study, we found that emodin, an anthraquinone compound present in rhubarb, strongly inhibited the stemness of GSCs by blocking the interaction of EGFR/EGFRvIII with Hsp90, which led to the degradation of EGFR/EGFRvIII and inhibition of downstream stemness signaling. 15 ESJ also induced proteasomal degradation of EGFR/EGFRvIII in GSCs. However, we failed to detect an inhibitory effect of ESJ on the interaction between EGFR/EGFRvIII and Hsp90, which indicated that EGFR/EGFRvIII degradation by ESJ occurred through a different mechanism.
In conclusion, our results indicate that certain components of ESJ are able to inhibit the sphere-forming and invasion activities of GSCs while sensitizing the cells to IR. We demonstrated that this probably occurred through degradation of EGFR/EGFRvIII, followed by suppression of downstream signaling pathways.
Footnotes
Acknowledgment
This work was supported by the Bio-Synergy Research Project (NRF-2014M3A9C4066487) of the Ministry of Science, ICT and Future Planning through the National Research Foundation.
Authors' Contribution
M.P. conceived and designed the experiments; J.K. performed the experiments; S.C. and J.L. analyzed the data; W.S. contributed reagents/materials/analysis tools; and M.P. and J.K. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.