
Abstract
This study evaluated whether long-term supplementation with dietary yerba mate has beneficial effects on adiposity and its related metabolic dysfunctions in diet-induced obese mice. C57BL/6J mice were randomly divided into two groups and fed their respective experimental diets for 16 weeks as follows: (1) control group fed with high-fat diet (HFD) and (2) mate group fed with HFD plus yerba mate. Dietary yerba mate increased energy expenditure and thermogenic gene mRNA expression in white adipose tissue (WAT) and decreased fatty acid synthase (FAS) mRNA expression in WAT, which may be linked to observed decreases in body weight, WAT weight, epididymal adipocyte size, and plasma leptin level. Yerba mate also decreased levels of plasma lipids (free fatty acids, triglycerides, and total cholesterol) and liver aminotransferase enzymes, as well as the accumulation of hepatic lipid droplets and lipid content by inhibiting the activities of hepatic lipogenic enzymes, such as FAS and phosphatidate phosphohydrolase, and increasing fecal lipid excretion. Moreover, yerba mate decreased the levels of plasma insulin as well as the homeostasis model assessment of insulin resistance, and improved glucose tolerance. Circulating levels of gastric inhibitory polypeptide and resistin were also decreased in the mate group. These findings suggest that long-term supplementation of dietary yerba mate may be beneficial for improving diet-induced adiposity, insulin resistance, dyslipidemia, and hepatic steatosis.
Introduction
O
Although a diet rich in fat induces obesity in most rodents, there may be variable responses in weight gain, glucose tolerance, insulin resistance, circulating triglycerides, and other parameters depending on the strain. Among mice, the C57BL/6J mouse strain is susceptible to HFD-induced obesity and particularly mimics human metabolic disorders such as central adiposity, hyperinsulinemia, insulin resistance, and hypercholesterolemia when fed an HFD. 2,3 Moreover, the excessive consumption of HFD has been suggested as a useful tool to cause NAFLD. 4
Numerous studies have shown that the consumption of foods rich in phytochemicals provides health benefits such as improvements in body weight and related metabolic disorders. 5 Yerba mate (dried leaf of the plant Ilex paraguariensis) is a traditionally consumed plant in many South American countries, particularly Argentina, Uruguay, Paraguay, southern and central-western Brazil, the Chaco region of Bolivia, and southern Chile. Various biological compounds, such as phenolic acids (chlorogenic acid and caffeic acids), flavonoids (quercetin, rutin, and kaempferol), caffeine, and saponins have been identified in yerba mate. 5 There is evidence that yerba mate has many pharmacologic activities, including antioxidant, anti-inflammatory, antiatherogenic, and glucose-lowering properties. 6 –8 Some studies have also reported its potential for the improvement of obesity. In HFD-fed Swiss strain mice, supplementation of aqueous extract of yerba mate (1 g/kg body weight) for 8 weeks decreased body weight, adiposity, and plasma and hepatic lipid levels. 6,7 In addition, it was demonstrated that the antiadiposity effect was associated with downregulated adipogenic gene expression in white adipose tissue (WAT) and upregulated expression of thermogenic genes in brown adipose tissue (BAT). 7,9 Moreover, administration of aqueous yerba mate extract (1 g/kg body weight) for 4 weeks decreased body weight by inhibiting food intake in C57BL/6J mice fed with HFD. 10 However, little study has been conducted to date on the effects of long-term supplementation with yerba mate and its underlying antiadiposity, antisteatotic, and hypolipidemic mechanisms are still not fully understood.
Accordingly, this study was performed to examine the effects and potential underlying the mechanisms of long-term supplementation with dietary yerba mate on HFD-induced obesity and related metabolic abnormalities, such as insulin resistance, dyslipidemia, and NAFLD in C57BL/6J mice.
Materials and Methods
Animals and diets
We purchased 20 male C57BL/6J mice from Jackson Laboratory (Bar Harbor, ME, USA) at 4 weeks of age. All mice were housed individually in stainless steel cages, and placed in a room where the temperature was kept at 22°C ± 2°C, the relative humidity at 40% ± 10%, and the light at a 12-h light–12-h dark cycle. After a 1-week acclimation period, the mice were randomly divided into two weight-matched groups and fed their respective experimental diets for 16 weeks as follows: HFD (control group; 20% [w/w] fat based on the AIN-76 diet plus 1% [w/w] cholesterol) and HFD with yerba mate (mate group; 0.62% [w/w], ∼400 mg/kg body weight/day) (Establecimiento Las Marías, Corrientes, Argentina). We incorporated the dried yerba mate powder directly into the HFD, and chlorogenic acid is a representative compound in yerba mate (34%, w/w). HFD contained 40 kcal% fat, 17 kcal% protein, and 43 kcal% carbohydrate with the fat sources consisting of lard (85% [w/w] of total fat) and soybean oil (15% [w/w] of total fat). The mice had free access to their assigned diet and the food consumption and body weight were measured daily and weekly, respectively, during the feeding period. At the end of the experimental period, the animals were anesthetized with isoflurane (5 mg/kg body weight; Baxter Healthcare, Deerfield, IL, USA) following a 12-h fasting period. Blood was drawn from the inferior vena cava into a heparin-coated tube and plasma was obtained by centrifuging the blood at 1000 g for 15 min at 4°C. After blood collection, the liver, six different locations of WATs (epididymal, perirenal, retroperitoneal, mesenteric, subcutaneous, and interscapular WAT), and interscapular BAT were removed, rinsed with physiological saline, and then weighed. Among these, the liver and epididymal WAT were snap-frozen in liquid nitrogen and stored at −70°C until subsequent analysis. All animal procedures were in compliance with and approved by the Ethics Committee for animal studies at Kyungpook National University (Approval No. KNU 2014-133).
Energy expenditure
Energy expenditure was measured at 14 weeks of feeding on the experimental diets using an indirect calorimeter (Oxylet; Panlab, Cornella, Spain). The mice were placed into individual metabolic chambers at 22°C ± 2°C, with free access to food and water. The 24-h energy expenditure was calculated using the volume of CO2 versus volume of oxygen (VCO2/VO2) ratio.
Blood analysis
Concentrations of plasma triglyceride, total cholesterol, and high-density lipoprotein (HDL)-cholesterol were determined enzymatically using commercial kits (Asan Pharm, Seoul, Korea), and plasma free fatty acid concentration was determined using a commercial kit from Wako (Tokyo, Japan). The atherogenic index and ratio of HDL cholesterol to total cholesterol (HTR) were calculated from lipid profiles as follows: atherogenic index = [(total cholesterol)–(HDL cholesterol)]/(HDL cholesterol) and HTR = [(HDL cholesterol)/(total cholesterol)] × 100.
At the end of the experimental period, fasting blood glucose concentrations were monitored in the venous blood drawn from the tail vein using a glucometer (Gluco Dr supersensor, Allmedicus, Korea). Fasting plasma glucose levels were determined using a commercially available kit (Asan Pharm, Seoul, Korea), and fasting plasma insulin levels were determined using a multiplex detection kit from Bio-Rad (Hercules, CA, USA). Homeostasis model assessment of insulin resistance (HOMA-IR) was calculated according to the homeostasis of the assessment as follows: HOMA-IR = [fasting plasma insulin concentration (mU/L) × fasting plasma glucose concentration (mg/dL) × 0.05551]/22.5. An intraperitoneal glucose tolerance test (IPGTT) was performed during the 15th week after beginning the diet experiments. The mice were fasted for 12 h and then injected intraperitoneally with glucose (0.5 g/kg body weight). Blood glucose concentrations were measured in tail vein blood using the Gluco Dr supersensor glucometer at 0, 30, 60, and 120 min after glucose injection.
Concentrations of plasma leptin, gastric inhibitory peptide (GIP), and resistin were determined using a multiplex detection kit (Bio-Rad, Hercules, CA, USA). Plasma aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were determined using enzymatic kits (Asan, Seoul, Republic of Korea).
Lipid analyses in the liver and feces
Hepatic lipids were extracted using the method of Folch et al. 11 and extracts were dried under nitrogen gas. The dried lipid residues were dissolved in 1 mL of ethanol. Triton X-100 and a sodium cholate solution prepared in distilled water were added to 200 μL of the dissolved lipid solution for emulsification; free fatty acid, triglyceride, and cholesterol contents were estimated using the same method as used for the plasma lipids analyses.
Before the mice were sacrificed, feces were collected for 7 consecutive days and dried overnight. Lipids from dried feces were extracted using the method of Folch et al., 11 with a slight modification; fecal lipids were extracted using chloroform and methanol (2:1, v/v) for 24 h at 4°C and were centrifuged at 900 g for 10 min. Thereafter, the supernatant was collected, dried at 50°C, and dissolved in ethanol. Fecal lipid levels were also analyzed with the same enzymatic kit used for the plasma analysis.
Morphological analysis of liver and epididymal fat
The liver and epididymal WAT were fixed in 10% (v/v) paraformaldehyde/phosphate-buffered saline and embedded in paraffin for staining with hematoxylin and eosin. Stained areas were viewed using an optical microscope (Zeiss Axioscope, Oberkochen, Germany) with a magnifying power of × 200.
Measurement of enzyme activity
To measure the lipid-regulating enzyme activity in the liver, we prepared mitochondrial, cytosolic, and microsomal fractions according to the method reported by Hulcher and Oleson, 12 with a slight modification, and protein concentrations were determined using the Bradford method. 13 Activity of fatty acid synthase (FAS) was determined in the cytosolic fraction by monitoring the malonyl coenzyme A-dependent oxidation of NADPH at 340 nm, 14 where the activity represented the oxidized NADPH nmol/min/mg protein. Mitochondrial fatty acid β-oxidation was measured spectrophotometrically by monitoring the reduction of NAD+ to NADH at 340 nm in the presence of palmitoyl-CoA as described by Lazarow. 15 The microsomal phosphatidate phosphohydrolase (PAP) activity was determined using the method of Walton and Possmayer. 16
RNA isolation and real-time quantitative reverse transcription-polymerase chain reaction
For detection of gene expression, total RNA was extracted from epididymal WAT using TRIzol reagent (Invitrogen Life Technologies, Grand Island, NY, USA) according to the manufacturer's instructions. DNase digestion and subsequent reprecipitation in ethanol were used to remove any DNA and phenol contamination, respectively. For quality control, RNA purity and integrity were evaluated using the Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA).
Total RNA (1 μg) was reverse transcribed into cDNA using the QuantiTect® reverse transcription kit (Qiagen, Hilden, Germany), and mRNA expression was quantified by real-time quantitative reverse transcription-polymerase chain reaction (RT-PCR) using the SYBR green PCR kit (Qiagen, Hilden, Germany) and the CFX96TM real-time system (Bio-Rad, Hercules, CA, USA). The values for gene expression were normalized to GAPDH expression, and the relative gene expression was calculated using the 2−ΔΔCt method. Primer sequences and cycling conditions are available upon request.
Statistical analysis
All data are expressed as the mean ± SEM. Statistical analysis was carried out using the statistical package for the social sciences software (SPSS) program (version 18; SPSS, Inc., Chicago, IL, USA). A student's t-test was used to assess the differences between groups.
Results and Discussion
This study showed that supplementation of HFD with yerba mate (∼400 mg/kg body weight) for 16 weeks significantly decreased final body weight and body weight gain compared to HFD control mice (Fig. 1A, B). Moreover, morphological observations indicated that epididymal adipocyte size in the yerba mate group was smaller than that in the control group (Fig. 1C). The weight of epididymal, visceral (sum of epididymal, perirenal, mesenteric, and retroperitoneal WAT weight), and total WAT (sum of visceral, subcutaneous, and interscapular WAT weight) was also lower in the yerba mate group than in the control group, although supplementation did not alter the weight of subcutaneous WAT (Fig. 1D). These results support previous findings showing that yerba mate protects against body weight gain and adiposity. 17 –19

Effect of yerba mate on body weight
However, there have been contradictory reports on the effectiveness of yerba mate for lowering plasma leptin, which is produced by adipocytes and whose circulating concentration appears to reflect total body fat. 20 Pang et al. 17 reported that supplementation with yerba mate (∼100 mg/kg body weight) for 60 days decreased plasma leptin levels in HFD-fed rats. Consistent with this finding, this study also observed that yerba mate significantly decreased plasma leptin levels (Fig. 1E). In C57BL/6J mice fed HFD, supplementation with aqueous extract of yerba mate (500 mg/kg body weight) for 4 weeks also decreased body weight, adiposity, and blood leptin levels. 10 Conversely, Hussein et al. 18,19 reported an increase in plasma leptin levels after supplementation with yerba mate in the metabolic syndrome model, Tsumura Suzuki obese diabetic mice, and in HFD-fed ddY mice, which may be related to the different models, species, or doses used in these studies.
Weight gain and excessive accumulation of body fat are caused by positive energy balance, which is a result of an excess of food energy intake relative to energy expenditure. In this study, food intake did not differ between groups (Fig. 1F). However, this study demonstrated, for the first time to the best of our knowledge, that yerba mate-supplemented mice exhibited significantly increased energy expenditure compared to the control group (Fig. 1G). In addition, yerba mate significantly upregulated mRNA expression of thermogenic genes, such as UCP1 and CIDEA, in epididymal WAT (Fig. 1H), although there was no significant difference in BAT weight (data not shown). Mitochondria-rich BAT is a key site of heat production; however, thermogenesis is also increased in WAT, which contains patches of brown-like cells (“brite” or “beige” adipocytes). 21,22 The brown-like adipocytes within WAT express thermogenic genes, including UCP1 and CIDEA, and share similar thermogenic characteristics to BAT. 23 The mRNA expression of UCP1, which uncouples oxidative phosphorylation from ATP synthesis and dissipates the energy for thermogenesis, 24 is generally downregulated in WAT by feeding of HFD, and upregulation of UCP1 in WAT protects against obesity. 25,26 The expression of CIDEA, which has been reported to have a role in regulating thermogenesis and obesity, 27 has also been downregulated in the WAT of HFD-induced obese mice. 25
These thermogenic genes (UCP1 and CIDEA) are controlled by PPARγ coactivator-1 α (PGC1α), a key regulator of energy metabolism. 28 Deficiency of PGC1α in WAT downregulates the expression of such thermogenic genes. 29 PPARα is also known to induce UCP1 expression, 30 and activation of PPARα by synthetic ligands induces a brown fat-like gene program in WAT. 31 Notably, this study showed that yerba mate-supplemented mice also exhibited higher mRNA expression of PGC1α and PPARα in epididymal WAT compared to the control group (Fig. 1H). Therefore, it is possible that the antiadiposity effect of yerba mate is related to the increased energy expenditure, in part, by inducing the formation of brown fat-like cells in WAT, without reduction in food intake. Similar to our findings, in HFD-fed Swiss strain mice, aqueous yerba mate extract did not alter food intake, but improved adiposity by upregulating thermogenesis-related genes (PGC1α and UCP1) in BAT, although they did not determine energy expenditure. 6,7 In contrast, two other studies have reported food intake-lowering effects of aqueous yerba mate extract in HFD-fed ddY mice and Tsumura Suzuki obese diabetic mice. 18,19 The discrepancy may be due, in part, to the differences in preparation (e.g., powder vs extract or different method of extraction), composition, and doses of yerba mate or animal models.
In this study, we also observed that yerba mate downregulated the mRNA expression of FAS, which catalyzes the first committed step in de novo lipogenesis, in epididymal WAT (Fig. 1H). Recently, Lodhi et al. 32 have reported that deletion of adipose FAS in mice not only ameliorated HFD-induced obesity but also increased energy expenditure and thermogenic gene expression in WAT, suggesting that the inhibition of adiopose tissue lipogenesis reprograms thermogenesis to decrease diet-induced obesity.
In addition to the antiobesity effect described above, this study further demonstrated that yerba mate decreased levels of fasting plasma insulin and HOMA-IR, a well-known insulin resistance predictor, and improved glucose tolerance in HFD-fed mice, although it did not alter fasting blood glucose concentrations (Fig. 2A–D). Moreover, yerba mate significantly decreased the circulating level of resistin and GIP (Fig. 2E), which are adipokines proposed to promote hyperinsulinemia and insulin resistance. 1,33 Circulating resistin level is elevated in the obese state and is proposed to link obesity with insulin resistance. 1,34 Previous studies have also reported a strong relationship between HFD-induced obesity and hypersecretion of GIP, a gastrointestinal hormone that is released from the duodenum after nutrient intake and stimulates postprandial insulin secretion. 35 –37 Chronic reduction of GIP secretion protects against HFD-induced obesity and insulin resistance in mice. 35 Therefore, the beneficial effects of yerba mate on the secretion of resistin and GIP may lead to an amelioration of hyperinsulinemia and insulin resistance in HFD-induced obese mice.
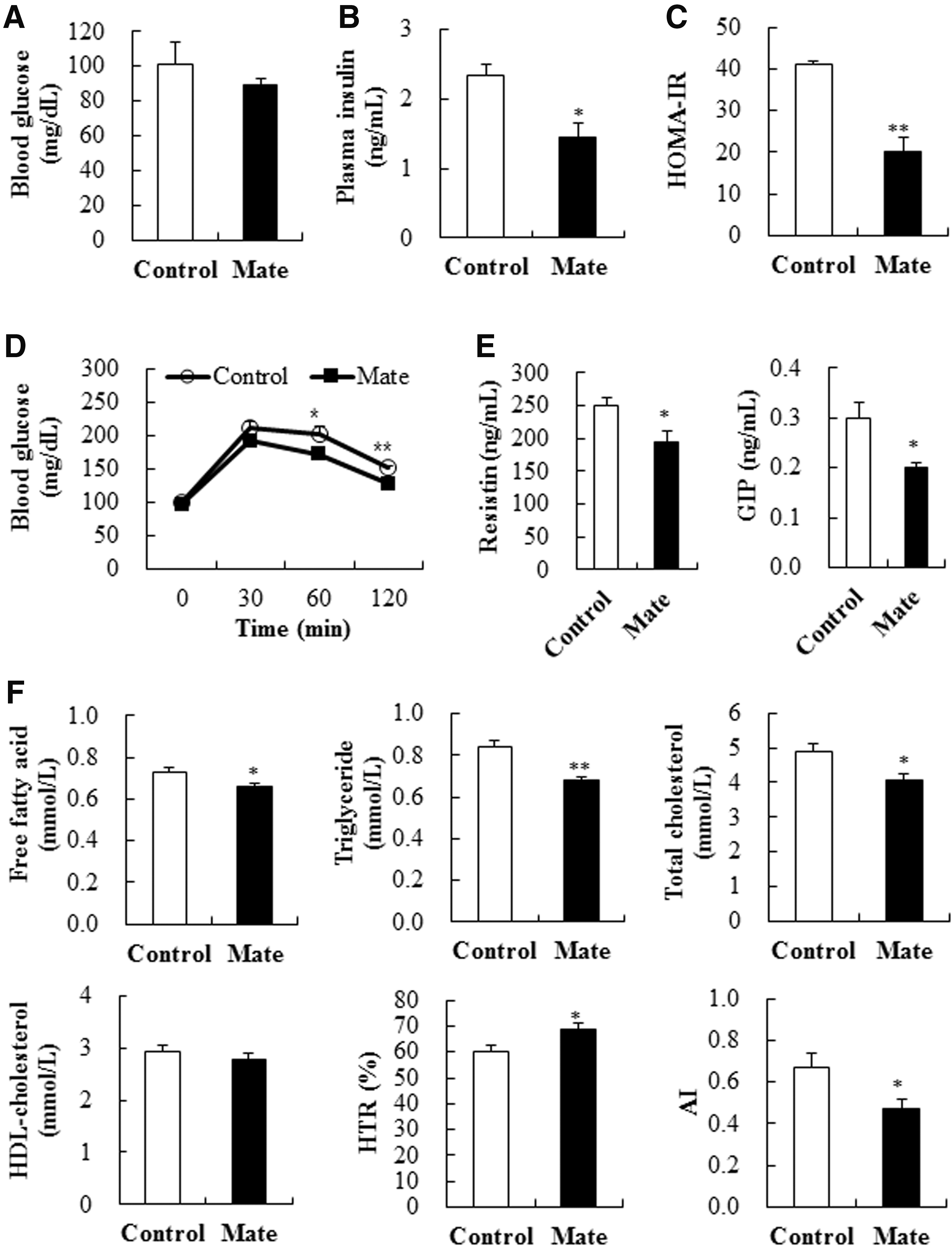
Effects of yerba mate on circulating glucose
The improved dyslipidemia and hepatic steatosis observed in yerba mate-supplemented obese mice may also provide an explanation for the enhanced insulin sensitivity, as hyperlipidemia and hepatic steatosis are often associated with insulin resistance and β-cell dysfunction. 38 In the obese state, plasma free fatty acid concentrations are commonly elevated, primarily consequent to their release from enlarged adipose tissue, and elevated circulating free fatty acids inhibit the antilipolytic action of insulin that further increases free fatty acid release into the circulation. 39 Moreover, under fasting conditions, plasma free fatty acids constitute the main substrate for hepatic triglyceride production in the form of very large density lipoproteins that are secreted into the plasma, 40 and insulin resistance promotes the delivery of free fatty acid to the liver, resulting in the stimulation of hepatic triglyceride synthesis and secretion. 41 Insulin resistance is also positively associated with hepatic cholesterol synthesis and increases the risk for coronary artery disease even in the absence of hyperglycemia. 42,43 In this study, we observed that yerba mate significantly decreased plasma free fatty acids, triglycerides, total cholesterol, and AI levels, whereas it increased HTR (Fig. 2F). In addition, yerba mate-supplemented obese mice exhibited markedly decreased hepatic free fatty acid, triglyceride, and cholesterol contents and lipid droplet accumulation, as well as plasma AST and ALT levels, which are used as an indicator of liver cell damage and NAFLD 44 (Fig. 3A–C).
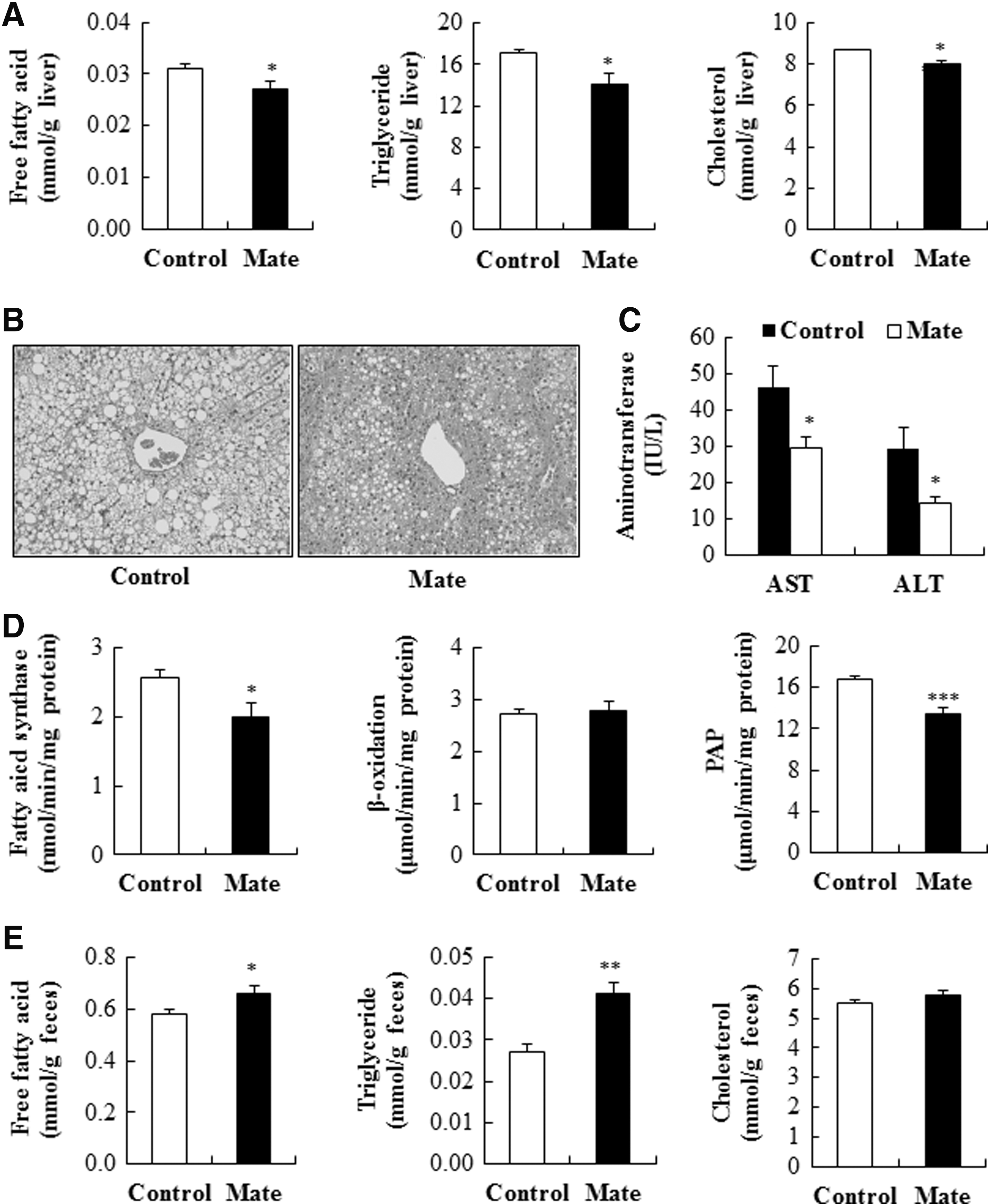
Effects of yerba mate on hepatic lipid contents
Along with increased free fatty acid delivery to the liver from lipolyzed adipose triglycerides, hepatic steatosis can be caused by increased de novo lipogenesis and decreased fatty acid oxidation in the liver. In this study, although yerba mate did not affect hepatic β-oxidation, the activities of hepatic FAS and PAP, key regulatory enzymes in the synthesis of fatty acids and triglycerides, respectively, were significantly lowered in yerba mate-supplemented obese mice compared to control obese mice (Fig. 3D), suggesting it may have inhibited de novo lipogenesis. Moreover, supplementation of yerba mate significantly increased fecal free fatty acid and triglyceride excretion (Fig. 3E). Consequently, the coordinated action of decreasing circulating lipids and de novo lipogenesis in the liver, while increasing fecal lipid excretion appeared to contribute to the beneficial effects of yerba mate on dyslipidemia and hepatic steatosis in the HFD-induced obese mice. In turn, such effects are also closely linked with the increased insulin sensitivity induced by yerba mate in these animals as well.
In conclusion, this study first demonstrated that long-term supplementation of dietary yerba mate increased energy expenditure and upregulated mRNA expression of thermogenic genes in WAT, whereas it downregulated mRNA expression of FAS in WAT, which may result in reduced adiposity in HFD-induced obese mice. Moreover, yerba mate improved dyslipidemia and hepatic steatosis through a decrease in hepatic lipogenic enzyme activity with a simultaneous increase in fecal lipid excretion, and these beneficial effects appeared to be related to the amelioration of insulin resistance induced by HFD. Figure 4 illustrates the possible mechanisms by which yerba mate may improve metabolic disturbances in HFD-induced obese mice. These findings support previous reports regarding the antiobesity effect of yerba mate and suggest that yerba mate may represent a useful natural candidate for the control of obesity and obesity-related insulin resistance, dyslipidemia, and hepatic steatosis. However, further studies are needed to determine maximum tolerated dose of oral yerba mate powder.
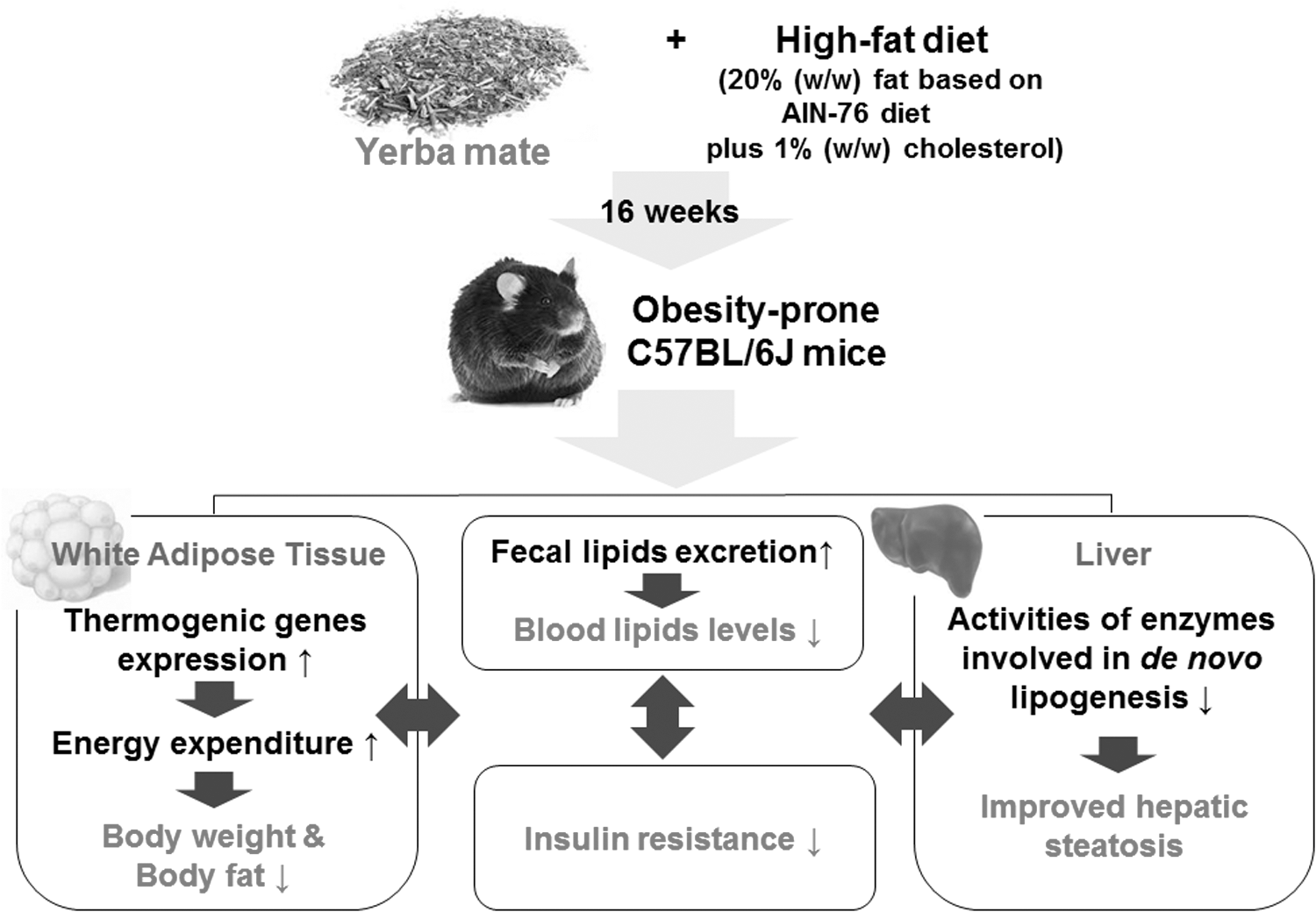
Potential mechanisms of action of yerba mate in the regulation of HFD-induced obesity and its related metabolic disturbances. The long-term supplementation of yerba mate increases energy expenditure and mRNA expression of thermogenic genes in WAT and decreases fatty acid synthase mRNA expression in WAT, indicating that these changes may be potential mechanisms for ameliorating the adiposity and insulin resistance induced by HFD. Moreover, the antiadiposity and insulin-sensitizing action of yerba mate may be associated with the improved dyslipidemia and hepatic steatosis, which may be caused by decreased hepatic lipogenic enzyme activities as well as increased fecal lipid excretion.
Footnotes
Acknowledgment
This work was supported by a Research Grant of Pukyong National University (2016).
Author Disclosure Statement
No competing financial interests exist.