
Abstract
Porphyra tenera, also known as nori, is a red algal species of seaweed. It is cultivated in Asia for culinary purposes. We report that P. tenera extract (PTE) enhances the immune response in mouse macrophages. We found that P. tenera extract regulates the NF-κB IκB kinase (IKK) signaling pathway, and we assessed the expression and translocation of p65, a subunit of NF-κB, in RAW264.7 mouse macrophage cells after treatment with PTE. We also investigated the effects of 10% ethanol PTE (PTE10) in RAW264.7 cells. The production of IL-10, IL-6, TNF-α, and IFN-γ was induced by PTE treatment of the macrophages, and PTE also enhanced p-IκB and p-AKT. PTE10 showed no cytotoxicity at 10–20 μg/mL in RAW264.7 cells. PTE10, in fact, increased cell viability at 24 h, stimulated macrophage cells, and induced the phosphorylation of Akt. Akt stimulates IKK activity through the phosphorylation of IKKα and enhances immune activity through the activation of NF-κB. In this study, NF-κB activation was induced by increasing p-NF-κB and p-IKK. A subunit of NF-κB, p65, was located in the nucleus and increased the expression of various cytokines. PTE thus enhanced the immune response through IκB-α immunostimulation signaling in RAW264.7 cells. PTE10 has potential therefore for development of future treatments requiring immune system stimulation.
Introduction
P
The human immune system is responsible for fighting foreign invaders of the body, such as bacteria and viruses. Poor nutrition has been shown to result in increased incidence of infection and slow recovery and healing from infection. Cytokines (IL-4, IL-1β, and IL-2) released from innate immune cells play key roles in the immune response. These cytokines are the source of regulatory signals that initiate immune response to pathogens. 3 We have reported previously that PTE possesses an immunostimulatory function. 4 The cell signaling mechanism underlying this is still unknown. In this study, we evaluated the mechanism of immunostimulation enhanced by P. tenera extract (PTE) and assessed the effects of PTE on RAW247.6 cells in vitro. We discuss PTE and summarize recent advances in the exploration of its function, focusing on those related to the modulation of immune responses.
NF-κB is an important transcription factor for the immunoglobulin κ gene in mature B-lymphoid cells and the protection of cells against external stimuli. 5 NF-κB is a nuclear factor that can bind to the κB site in the immunoglobulin κ light-chain enhancer. The presence of constitutive NF-κB activity in RAW264.7 cells indicates the importance of NF-κB in the regulation of the immune response. PTE can enhance the immune response, which is characterized by the activation of macrophages and natural killer cells, and the expression of various cytokines in a dose-dependent manner. Using this information, we tested the potential for PTE to modulate immune reactions when orally administered.
The results of the present study support the hypothesis of a strong interaction between NF-κB and the phosphatidylinositol 3-kinase (PI3K)/AKT pathway in the modulation of the immune system in RAW264.7 cells. The serine/threonine kinase Akt, also known as protein kinase B, is activated by lipid products of PI3K. 6 Akt phosphorylates various protein targets. We have found that PTE regulates Akt, ERK, and JNK activities by inducing phosphorylation. It is widely known that the NF-κB signaling cascade interacts with several parallel pathways, including the signaling cascades initiated by PI3K and Akt. Akt, functioning as a key downstream target of PI3K, activates IκB kinase (IKK) complexes. Sequentially, IL-10, IL-6, TNF-α, and IFN-γ have been found to be produced by macrophages in vitro upon stimulation by PTE. We propose therefore that PTE is an immunostimulant.
Finally, our present data suggest that the functional effects of 10% ethanol PTE (PTE10) are dependent on the phosphorylation of the NF-κB subunit p65 through Akt-mediated IκBα. Thus, we demonstrate that the immune-enhancing functions of PTE10 occur via the phosphorylation and degradation of IκB and the phosphorylation of p65 through Akt activity. PTE10 is therefore an effective immunostimulant and, importantly, comes from a natural source.
Materials and Methods
Cell culture
All cell lines were obtained from the American Type Culture Collection. 293T cells and RAW264.7 cells were cultured in Dulbecco's modified Eagle's medium (Hyclone; GE Healthcare Life Sciences, Pittsburgh, USA) and supplemented with 10% fetal bovine serum (Hyclone) and 1% antibiotics and antimycotics (Hyclone). All cell lines were grown at 37°C in a 5% CO2 incubator.
Preparation of PTE10
In all, 10 kg extractions of P. tenera were performed with 600 L 10% ethanol solvent for 3 h at 80°C, yielding PTE10. The solvent was filtered and concentrated down to 60 L before being freeze dried. The dried extracts were then mixed and dissolved in water for 10 min. The supernatants were reserved after centrifugation at 17,000 g for 10 min.
Preparation of lipopolysaccharides and red ginseng extract
Lipopolysaccharides (LPSs) were prepared from Escherichia coli O127:B8 (cat. No. L4516; Sigma-Aldrich, St. Louis, MO, USA). After 30% red ginseng extract (RGE; Korea Ginseng Corporation, Daejeon, Korea) was filtered with a 0.2 μM syringe filter, the RGE was diluted to 5%.
Cell viability assay
Cell viability was determined using the WST-1 (Ez-Cytox; Daeil Lab, Seoul, Korea) assay. Cells were plated in 96-well plates at a density of 1–3 × 103 cells/well. Then they were incubated at 37°C in a CO2 incubator. Different concentrations of RGE and LPS or PTE10 were added to each well. After incubation for 6, 12, 24, or 48 h, 10 μL WST-1 reagent was added to each well and the wells were incubated again for 90 min at 37°C in a CO2 incubator. The absorption was measured at 450 nm (reference wavelength 600–650 nm) with a micro plate reader (Model 550; Bio-Rad Laboratories, Hercules, CA, USA). All WST-1 assay results are presented as mean (±SD) of three independent experiments.
RNA extraction and quantitative real time-polymerase chain reaction analysis of cytokine expression
Total RNA was isolated with the RNA EasySpin Kit according to the manufacturer's instructions (Intron Biotechnology, Seongnam, Korea). Total RNA from each sample was reverse transcribed with random primers using a StrataScript™ Reverse Transcriptase Kit (Agilent Technology, Santa Clara, CA, USA) according to the protocols of the manufacturer. All samples were normalized to human GAPDH and expressed as fold induction. All reactions were done in triplicate. Relative expression and standard deviations were calculated using the comparative quantification method. The primers used were as follows: mIL-10, 5′- GCTCTTACTGACTGGCATGAG-3′ and 5′-CGCAGCTCTAGGAGCATGTG-3′; mIL-6, 5′-GGACCAAGACCATCCAATTCATCTTGAAA-3′ and 5′-GACCACAGTGAGGAATGTCCACAAA-3′; mTNF-α, 5′-CATCTTCTCAAAAT TCGAGTGACAA-3′ and 5′-TGGGAGTAGACAAGGTACAACCC-3′; mIFN-γ, 5′-GCATCTTGGCTTTGCAGCT-3′ and 5′-CCTTTTTCGCCTTGCTGTTG-3′; mActin, 5′-CTATGAGCTGCCTGACCGTC-3′ and 5′-AGTTTCATGGATGCCACAGG-3′.
Western blot analysis
For western blotting, RAW264.7 cells were washed with cold PBS, scraped off, and harvested. Cell extracts were prepared with lysis buffer (50 mM Tris–HCl [pH 7.5], 150 mM NaCl, 1% NP-40, 10 mM NaF, 10 mM sodium pyrophosphate, and protease inhibitors) and incubated for 30 min on ice. The lysates were centrifuged at 20,000 g for 10 min at 4°C. Total cell lysate protein was separated on 8% and 12% SDS-PAGE gels and transferred to nitrocellulose membranes. The membranes were blocked by incubation for 2 h in 5% (w/w) nonfat Difco™ skim milk (BD Biosciences, Franklin Lakes, NJ, USA) blocking buffer with 1 × PBST. The blocked membranes were incubated for 2 h or overnight at 4°C with the primary antibody. After being washed three times with 1 × PBST, the membranes were incubated with the secondary HRP-conjugated antibody for 1 h. The results were then subjected to western blot analysis and visualized in a developer.
Nucleus and cytoplasm fractionation
After the preincubation of Raw264.7 cells (1 × 106 cells/mL) for 18 h, PTE (10 and 20 μg/mL) or LPS (1 μg/mL) were added and incubated with the cells for 6 h. We washed the harvested cells two times with PBS, centrifuged them at 3000 rpm for 3 min at 4°C, removed the supernatant, added 1 mL Sol A buffer (10 mM Tris [pH 7.4], 10 mM KCl, 3 mM MgCl2, 0.5% NP-40, and protease inhibitor cocktail), resuspended them, chilled them for 20 min on ice, and then centrifuged them again at 4000 rpm for 5 min at 4°C. We transferred the supernatant into tube A, added 200 μL Sol B buffer (20 mM Tris [pH7.9], 0.42 M NaCl, 0.2 mM EDTA, 10% glycerol, 2 mM DTT, and protease inhibitor cocktail), and resuspended them with a syringe five times. Next, we chilled them for 20 min on ice, centrifuged them again at 13,000 rpm for 20 min, and transferred the supernatant into tube B, where tube A contained cytosol protein and tube B contained nuclear protein extract.
Luciferase assays: NF-κB transcription activity
To measure NF-κB transcriptional activity, 293T cells were transiently cotransfected with TLR4 expression plasmid and reporter construct β-gal or NF-κB-Luc plasmids. The Renilla luciferase reporter plasmid was included as an internal control. The cells were harvested and extracted. Then luciferase activity was measured, according to the manufacturer's instructions (Promega, Madison, WI, USA). All reporter activities were normalized relative to Renilla luciferase activities and are presented as mean (±SD) of three independent experiments.
Confocal microscope analysis
To perform confocal assay staining, cells were plated in a confocal dish (101350; SPL, Seoul, Korea) at a density of 1–2 × 104 cells/plate and were incubated 37°C in a CO2 incubator. After preincubating RAW264.7 cells (1–2 × 104 cells/mL) for 24 h, PTE10 (10 and 20 μg/mL) or LPS (1 μg/mL) and RGE (5%) samples were added and incubated with the cells for 6 h. We then washed the cells three times using PBS 300 mL/well and treated them with 2% PFA 300 mL/well in 0.1% Triton X-100 for 10 min fixation. We added 0.02 M glycine PBS 300 mL/well for three washes and 10% BSA (in PBST) in 300 mL/well and then incubated the cells at 37°C in a CO2 incubator. The primary antibody was diluted 1:250 in a 300 mL/well for 1 h incubation with 300 mL/well PBS for three washes, and the secondary antibody for 300 mL/well FITC (green) for 1 h.
Statistical analyses
Statistical analyses were performed using ANOVA and SPSS software (SPSS, Inc., Chicago, IL, USA). A statistical threshold of P < .05 was considered statistically significant.
Results
AKT activation and cytotoxic effects of PTE10
The cytotoxicity of PTE10 in RAW264.7 cells was examined using a WST-1 assay after treatment with this agent for 6, 12, 24, and 48 h. A previous study determined the cytotoxicity of PTE10 to be in the range of 10–40 μg/mL. 3 Therefore, we selected 10 and 20 μg/mL concentrations for our present experiments. As is shown in Figure 1A, PTE10 had no cytotoxic effects in RAW264.7 cells at 10–20 μg/mL, but in fact increased cell viability at 24 h. To investigate PTE10-mediated immune enhancement, we examined the signaling pathways of AKT and Ik-Bα via LPS or PTE10 treatment of RAW264.7 cells from 0 to 48 h. The expression of AKT and Ik-Bα was determined by western blot. AKT and Ik-Bα were phosphorylated after exposure of the cells to LPS or PTE10. The p-AKT and p-Ik-Bα expression levels were also increased initially (Fig. 1B), but were then reduced between 24 and 48 h after treatment. These results demonstrated a hyperphosphorylation of AKT and IκBα by PTE10. The effects of different doses of PTE10 extract on cell proliferation and the production of immunostimulants suggested that the increased cell proliferation was a major outcome of the AKT pathway activation. Thus, our findings suggested that the immunostimulant effects of PTE10 are comparable to those of RGE.
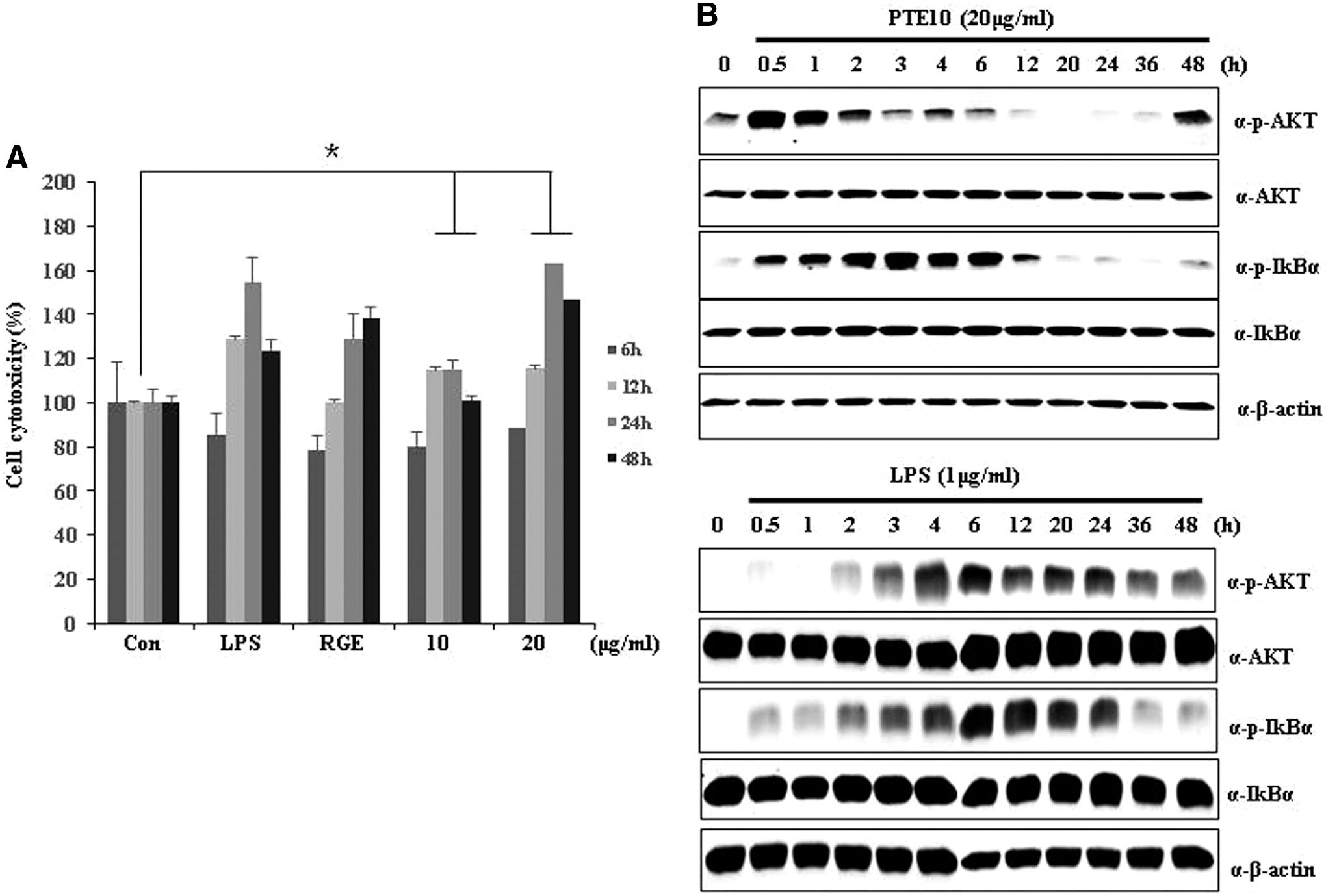
The cytotoxicity of PTE and LPS in RAW264.7 cells. AKT and IκBα signaling response and cytotoxic response to LPS (1 μg/mL) and 10% ethanol PTE (PTE10) (20 μg/mL) in Raw264.7 cells.
PTE10 induces NF-κB signaling
We previously reported that PTE10 induces the phosphorylation of AKT, JNK, and ERK. 3,21 We assessed these signaling pathways to determine the activation of NF-κB in our current analysis. NF-κB was first identified as a regulator of the activation of the immune response and inflammation signaling. We examined the expression of p-IκBα and p-AKT after treatment with PTE10 or LPS and RGE on the NF-κB signaling pathway. As shown in Figure 2A, PTE10 induced the expression of p-AKT and p-IκBα by more than the control. This indicates that PTE10 dramatically stimulates the immune response by increasing the expression of p-IκBα in a dose-dependent manner. These events also involve the activation of the IKKs that phosphorylate IκB. 6 The degradation of IκB uncovers its nuclear localization sequence and enables NF-κB dimers to translocate to the nucleus, bind to the κB motifs present in the promoters of many genes, and regulate transcription. In this system, the binding of NF-κB will induce the transcription of IκB genes and thus provide a mechanism for the activation of NF-κB pathways. 7,8 Hence, we measured NF-κB transcription activity in 293T cells following LPS and RGE or PTE10 treatment using a luciferase assay. A representative result is presented in Figure 2B. NF-κB transcription activity was comparably increased by PTE10 and RGE. Therefore, PTE10 has a positive effect on the activation of NF-κB signaling and can thus influence immunostimulation and adjust the NF-κB signaling pathway.
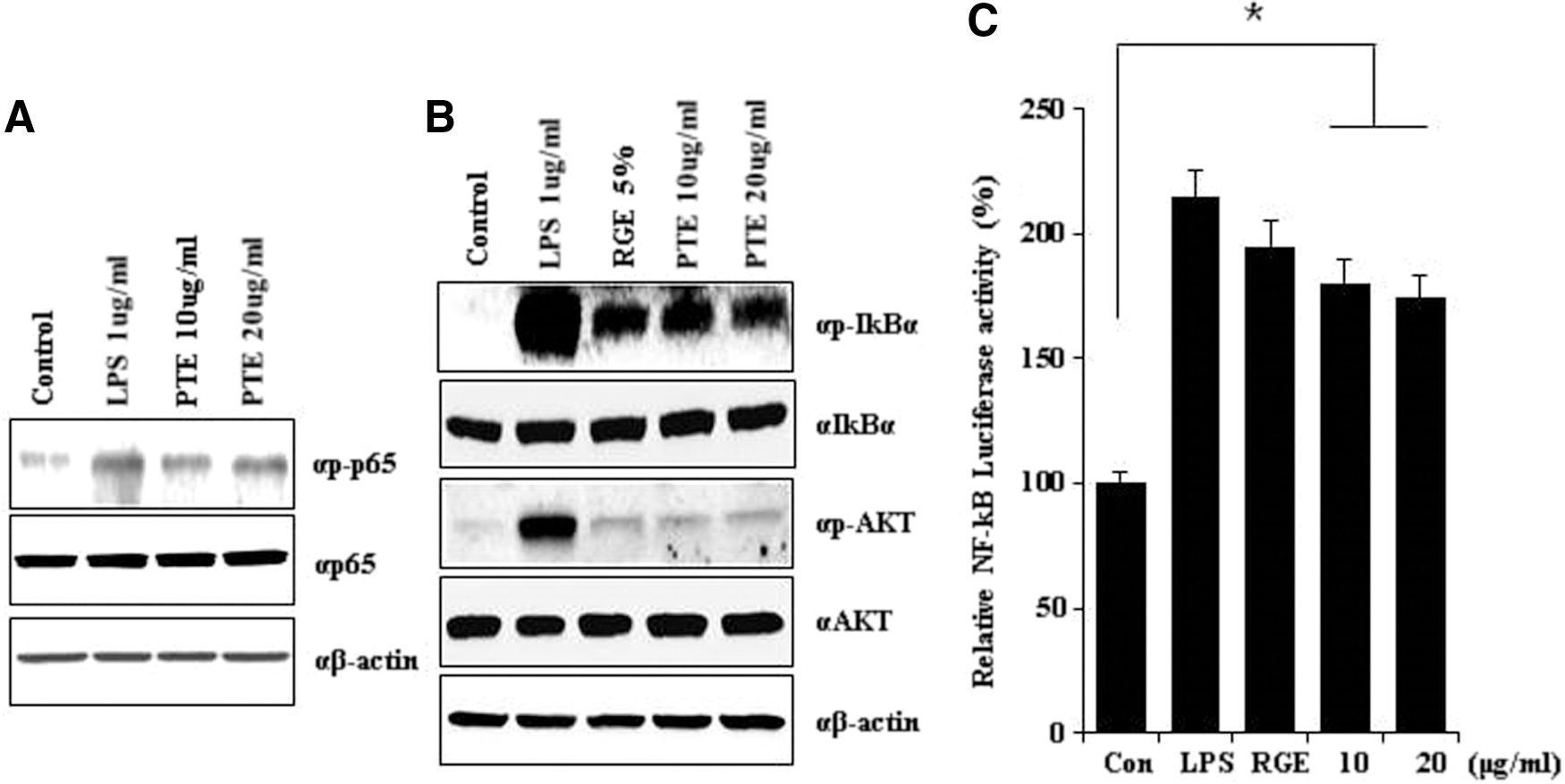
PTE10 induces the expression of transcription factors. PTE10 enhances the p-p65 level and NF-κB promoter activity in RAW264.7 cell lines.
PTE10 induces the nuclear translocation of NF-κB subunit p65
We tested the translocation of the NF-κB subunit p65 in mouse macrophage RAW264.7 cells treated with RGE and LPS (positive control), PTE10 at 10 μg/mL, PTE10 at 20 μg/mL, or distilled water (DW) (control) treatment. When PTE10 was used, the localization of p65 was changed in RAW264.7 cells. It was further observed that p65 enters the nucleus following PTE10 treatment. Western blot analysis indicated that NF-κB pathway activation is the mechanism whereby PTE10 influences the immune system (Fig. 3A).
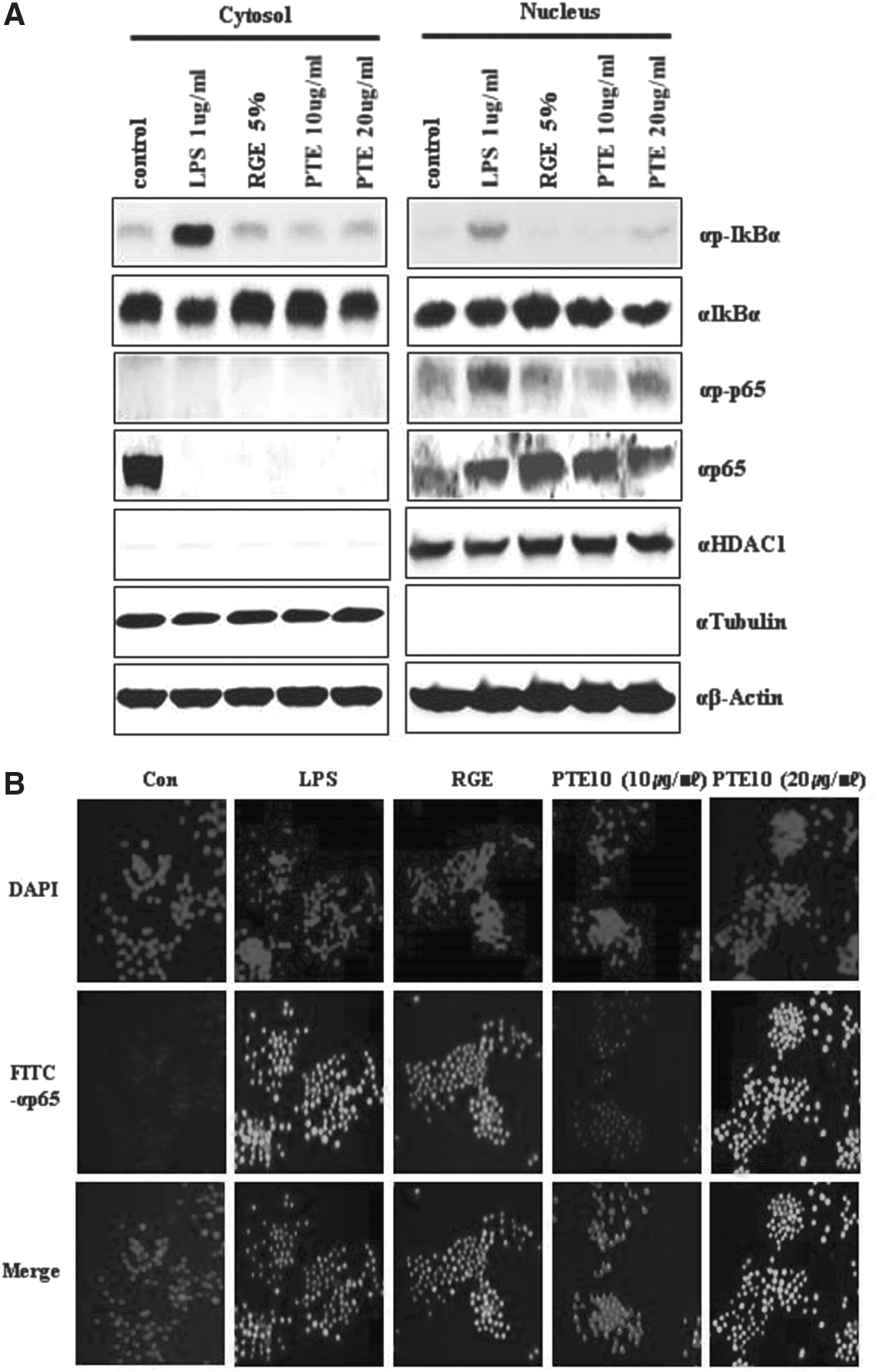
Localization of NF-κB and IκBα in RAW264.7 cells. Nuclear and cytoplasmic extracts were made from RAW264.7 cells following PTE10 treatment. p-p65 was found to be more highly expressed in the nuclear fraction.
To confirm the location of the p65 NF-κB subunit, we performed confocal microscope analysis of RAW264.7 cells after PTE10 treatment. The inactive p50/p65 heterodimer of NF-κB is normally located in the cytoplasm in a complex with its IκB inhibitory unit. Immunostimulation by PTE10 caused a dissociation of NF-κB from IκB and a rapid translocation of free NF-κB to the nucleus. These results confirmed the nuclear translocation of p65 in PTE10-treated RAW264.7 cells (Fig. 3B).
Activation of cytokines by PTE10
A previous study measured the secretion of cytokines such as IL-1β, IL-2, IL-4, and iNOS following PTE10 treatment of RAW264.7 cells. NO upregulation by PTE10 was accompanied by increasing iNOS expression, according to quantitative real time (qRT)-polymerase chain reaction (PCR) analysis (data not shown). 3 NF-κB plays a crucial role in immune activation and inflammatory response through the regulation of genes encoding proinflammatory cytokines, adhesion molecules, chemokines, growth factors, and inducible enzymes such as COX2 and iNOS. 9,10,16 –20,23,24 In addition, NF-κB can be activated within minutes by a variety of stimuli, including inflammatory cytokines such as TNF-α, interleukin-1, T-cell activation signals, growth factors, and stress inducers. 11,22
We evaluated the ability of PTE10 to enhance the immune system using PTE10 treatment of RAW264.7 cells. The proinflammatory cytokines IFN-γ, TNF-α, and IL-6 have been reported to be potent immune stimulatory cytokines and IFN-γ immunomodulators in activated macrophages and T-lymphocytes. IL-10, a cytokine with anti-inflammatory properties, has a central role in infection through limiting the immune response to pathogens and preventing damage to the host. Understanding the specific molecular events that regulate the production of IL-10 will help answer remaining questions that are important for the design of new strategies of immune intervention. Therefore, we examined the expression of cytokine mRNA by qRT-PCR after treatment with PTE10 or LPS and RGE (positive control). As shown in Figure 4, PTE10 induced greater IFN-γ, IL-6, TNF-α, and IL-10 expression than LPS. Taken together, these results indicate that PTE10 dramatically stimulates immune system activity by increasing the expression of cytokines in a dose-dependent manner (Fig. 4).
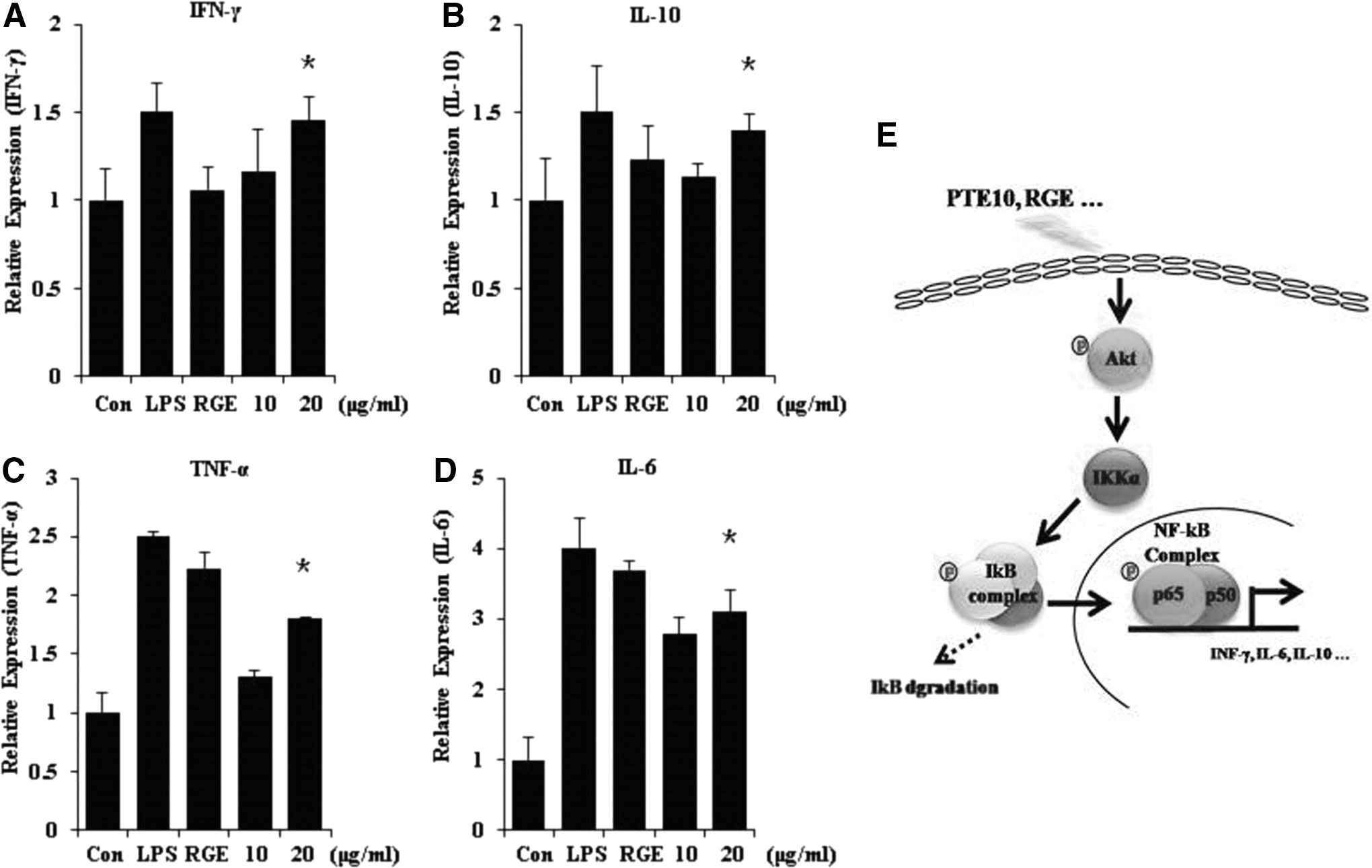
Proinflammatory cytokines are induced by PTE10. Proinflammatory and anti-inflammatory cytokines are induced in RAW264.7 cells by PTE10 treatment. Cytokines were assayed in extracts from the cells following treatment with LPS, RGE, and PTE10.
Discussion
P. tenera is known for its use in the Korean dish gimbab and in various other dishes. However, there are few reports on the effects of P. tenera on the immune response. In a previous study, we evaluated the effects of different PTEs on immunity. The extracts were obtained from P. tenera using industrial processes. Immunostimulation is beneficial for fighting infections, and although many artificial immunostimulants have been developed by pharmaceutical companies, they have undesirable side effects. 12,13 Therefore, natural compounds that stimulate the immune system with few or no side effects represent a promising alternative. 14,15 We found that PTE10 enhances both NO production and cytokine (IL-1β, IL-2, IL-4, IFN-γ, and iNOS) expression. Importantly, PTE is a novel immunostimulant from a natural product and enhanced cytokines in RAW264.7 cells.
In our present study, PTE10 was found to improve the NF-κB pathway as part of the immune response. The main organ of immunity in the human body is the spleen, and increases in cytokine production in splenocytes are a key indicator of immune reactivity. For this reason, we tested the immunomodulatory and cytotoxic effects of PTE10 on RAW264.7 cells. This is the first study to demonstrate the nontoxic, immune-stimulating effects of P. tenera for 6–24 h in vitro. PTE10 showed no cytotoxicity at 10–20 μg/mL dose levels in RAW264.7 cells. In fact, it increased cell viability at 24 h at this concentration range. It also increased cytokine expression (IFN-γ, IL-6, TNF-α, and IL-10). PTE10 has immunostimulatory effects, enhancing the secretion of IFN-γ, IL-6, TNF-α, and IL-10. We confirmed cytokine secretion using the human cytokine antibody array. Transcription factors, such as NF-κB and IκB genes, alter cytokine expression.
PTE10 can regulate immunostimulation through the NF-κB pathway. The mechanism by which PTE10 functions is the relocalization of NF-κB. PTE10 thus activates the immune system through the NF-κB pathway. NF-κB can be activated within minutes by a variety of stimuli, including inflammatory cytokines such as TNF-α. The proinflammatory cytokines (IFN-γ, TNF-α, and IL-6) are reported to be potent immunostimulatory cytokines. IFN-γ is an immunomodulator in activated macrophages and T lymphocytes. In addition, NF-κB activation is regulated by the IκB complex. As shown in Figures 1 and 2, IκB was phosphorylated by PTE10. As one factor of the NF-κB pathway, IκB regulates NF-κB phosphorylation through NF-κB upregulation. IκBa expression is also related to the immune system. In addition, we report NF-κB activation by the IκB complex.
Taken together, our present results indicate that PTE10 dramatically stimulates immune system activity by increasing the expression of cytokines in a dose-dependent manner. Through its regulation of the NF-κB pathway, cytokine activity is regulated by PTE10. As PTE10 is a natural immunostimulant found in food, its properties may lead to new immune-stimulating functional foods in the future.
In conclusions, in a previous study, we found that PTE10 enhanced the secretion of cytokines in the splenocytes and macrophages of mice. In this study, we further investigated PTE10 to determine whether immune system activity occurs via the NF-κB pathway. PTE10 activates various cytokines, such as proinflammatory cytokines IFN-γ and TNF-α, via phosphorylated IκBa and NF-κB. We thus found that PTE10 regulates the immune system through NF-κB signaling. The effects of immunization through the NF-κB pathway were assessed using RAW264.7 cells. PTE10 is a good immunostimulatory agent and has good potential in the future development of treatments that require immune system enhancement.
Footnotes
Acknowledgment
This research was supported by a grant from the Marine Biotechnology Program (Project No. PJT200672), funded by the Ministry of Oceans and Fisheries, Korea.
Author Disclosure Statement
No competing financial interests exist.