
Abstract
Grifola gargal Singer, a medicinal mushroom, has been found to be effective for the prevention and treatment of various chronic inflammatory diseases. However, the effects of G. gargal on allergic diseases are unknown. The present study investigated the effect of G. gargal extract on allergic bronchial asthma. Asthma was induced in mice by ovalbumin sensitization and inhalation. The grade of asthma was compared between mice fed with chow containing G. gargal extract and mice given standard chow. The human mast cell and eosinophilic cell lines were used for in vitro studies. G. gargal extract significantly reduced airway hyperresponsiveness, lung eosinophilic infiltration, lung interleukin (IL)-13 expression, and plasma IgE level and significantly increased IL-10 plasma levels compared to untreated control mice. Spleen regulatory T cells were significantly increased in mice treated with the G. gargal extract compared with untreated control mice. G. gargal extract significantly suppressed expression of cytokines in mast cells and eosinophils compared with control cells. Overall, these observations show that G. gargal extract augments the lung population of regulatory T cells and ameliorates allergic inflammation and airway hyperresponsiveness in mice with allergic bronchial asthma, suggesting the potential therapeutic benefit of G. gargal extract in allergic diseases.
Introduction
T
Grifola gargal Singer is an edible mushroom, with anti-inflammatory properties, that commonly grows in the southern regions of South American forests. 10,11 G. gargal is characterized by forming multipetaloid fruit bodies and by its peculiar almond flavor. The anti-inflammatory activity has been attributed to the abundant content of ergothioneine and ergocalciferol (vitamin D2) in the mycelia and fruit body of the mushroom. 11,12 Three novel sterols (gargalols) and a new sphingosine with osteoclast-forming suppressive activity have been also recently isolated from the fruit body of G. gargal. We have also previously demonstrated that by using an experimental mouse model, G. gargal extracts can prevent the progression of atherosclerotic disease by enhancing the population of regulatory T cells and by stimulating the secretion of chemokines that promote vascular repair. 10 In a clinical study, we also demonstrated that consumption of G. gargal is associated with decreased urinary deoxypyridinoline, a specific marker of bone resorption and osteoclastic activity. 11
The objective of the present study was to evaluate the efficacy of oral G. gargal extract (GGE) in a murine model of allergic bronchial asthma. The suppressive effects of gargalols on activated human mast cells and human eosinophil cells were also investigated through in vitro experiments.
Materials and Methods
Preparation of GGE
The G. gargal was provided by the Iwade Research Institute of Mycology Corporation (Tsu, Japan). Data on the mushroom are available online at MycoBank (Deposition No. MB#331520) and in the DNA Data Bank of Japan (
Experimental animals
Female BALB/c mice aged 8–12 weeks, weighing 17–23 g, were purchased from Nihon SLC (Hamamatsu, Japan). All animals were housed in a pathogen-free environment. Mice were kept on a constant 12-h light–12-h dark cycle in a temperature- and humidity-controlled room and were given water and standard mouse chow ad libitum before the beginning of the study. All experimental animals received humane care and all procedures were performed following guidelines described in the Guide for the Care and Use of Laboratory Animals issued by the National Academy of Sciences and published by the National Institutes of Health. The experimental animal protocol was approved by the Mie University's Committee on Animal Investigation (Approval No. 25-20-change1-1; Date September 12, 2016).
Experimental design
For induction of allergic bronchial asthma, mice were sensitized by intraperitoneal injection of 10 μg of aluminum-precipitated ovalbumin (OVA) on day 0 followed by booster injections on days 7, 14, and 21. 14 Animals that were injected with saline (SAL) by the intraperitoneal route on the same days were used as controls. Once the mice were sensitized, they were exposed to aerosolized 2% OVA during 30 min/day for six consecutive days (days 22, 23, 24, 25, 26, and 27 after the first sensitization with intraperitoneal OVA). 14 A UN-511 ultrasonic nebulizer (UN-511; Azwell, Osaka, Japan) was used for aerosol generation. Animals that received intraperitoneal injection of SAL and exposed to aerosolized SAL for six consecutive days were used as controls.
Mice were categorized into four treatment groups: (1) nonsensitized mice that received intraperitoneal injections of SAL and fed with normal food (normal food/SAL group, n = 7); (2) nonsensitized mice that received intraperitoneal injections of SAL and fed with GGE diet (GGE food/SAL group, n = 4); (3) OVA-sensitized mice fed with normal food (normal food/OVA group, n = 8); and (4) OVA-sensitized mice fed with GGE diet (GGE food/OVA group, n = 9). Airway hyperresponsiveness was measured on day 26 using a whole-body plethysmograph system (Buxco Electronics, Inc., Sharon, CT, USA), and mice were rechallenged with aerosolized 2% OVA before being sacrificed.
Measurement of airway hyperresponsiveness
Enhanced pause was measured as a marker of airway hyperresponsiveness after exposing mice to increasing concentrations of aerosolized methacholine using a whole-body plethysmograph system (Buxco Electronics, Inc.) as previously described. 15
Collection of bronchoalveolar lavage fluid and plasma and tissue homogenization
Twenty-four hours after the last OVA challenge, mice were sacrificed by an overdose of pentobarbital injected by the intraperitoneal route to obtain samples for biochemical and histological examinations. Bronchoalveolar lavage fluid (BALF) was obtained by cannulating the trachea with a 20-gauge needle before infusing the lungs twice with 0.75 mL of SAL as previously described. 16 The BALF samples were centrifuged, the cell pellets were resuspended, the number of total cells counted, and differential cell counts were determined after cytospin centrifugation and staining with May–Grunwald–Giemsa (Merck, Darmstadt, Germany). Blood was sampled by cardiac puncture and collected into tubes containing heparin, and then, plasma was prepared and stored at −80°C until analysis. The lung tissues were placed in a protease inhibitor cocktail (Nacalai Tesque, Kyoto, Japan) and homogenized using a Tomy Micro Smash MS-100R (Tomy Digital Biology, Tokyo, Japan) as previously described. 16
Biochemical analysis
Total protein in BALF was measured using a commercial dye-binding assay (Bio-Rad Laboratories, Hercules, CA, USA). Monocyte chemoattractant protein (MCP)-1, interleukin (IL)-4, IL-5, and IL-10 were measured using enzyme immunoassay kits from R&D System (Minneapolis, MN, USA).
Histological examination of lung tissues
Histological examinations were performed using hematoxylin and eosin (H&E) staining following standard methods. Briefly, the lung was excised, fixed in 4% formalin, and embedded in paraffin. Sections of 5 μm thick were deparaffinized, hydrated, and then used for H&E or immunohistochemical staining. Histopathological assessment by light microscopy of randomly selected sections was performed by a pathologist who was blinded to the experimental group. The lung sections stained with H&E were examined under a light microscope. An investigator blinded to the treatment group took microphotographs of the lung fields of all mice. Microphotographs were taken using an Olympus BX50 microscope with a plan objective combined with an Olympus DP70 digital camera (Tokyo, Japan). A WinROOF image processing software (Mitani Corp., Fukui, Japan) for Windows was used to quantitate the grade of cellular infiltration in the lung fields.
RNA extraction and reverse transcription–polymerase chain reaction
Total RNA was extracted using Sepasol RNA I SuperG (Nacalai, Osaka, Japan) according to the instructions of the manufacturer. One microgram of total RNA was denatured at 65°C for 10 min and then reverse-transcribed using ReverTra Ace Reverse Transcriptase (TOYOBO) and Oligo (dT) primer in a volume of 20 μL following the manufacturer's protocol. Amplification of cDNA was done by polymerase chain reaction (PCR) using gene-specific primers under the following conditions: 10 sec at 94°C, 20 sec at 60°C, and 40 sec at 72°C for 28–35 cycles. Supplementary Table S2 describes the sequences of the primers. Electrophoresis of the PCR products was performed on an agarose gel followed by ethidium bromide staining.
Flow cytometry analysis
To evaluate the number of CD4+CD25+ lymphocytes, splenic lymphocytes were incubated with the anti-CD16/32 antibody for 30 min before treatment with the following specific monoclonal antibodies: fluorescein-conjugated anti-CD25, phycoerythrin (PE)-conjugated anti-CD3, and PE-Cy5-conjugated anti-CD4 antibodies. Antibodies against CD4 (rat IgG2b, clone: GK1.5) and CD25 (rat IgG1, clone: PC61) were purchased from BioLegend (San Diego, CA, USA).
Cell culture
The human mast cell line HMC-1 was obtained from ATCC (Manassas, VA, USA) and maintained in Iscove's modified Dulbecco's medium (IMDM), supplemented with 10% fetal bovine serum (FBS), 2 mM
The human eosinophilic cell line EoL-1 was obtained from the RIKEN Cell Bank (Tsukuba, Japan) and maintained in the RPMI-1640 medium supplemented with 10% FBS, 100 IU/mL penicillin, 50 μg/mL streptomycin, and
Assays of mast cell and eosinophil activation
Human HMC-1 mast cell lines and eosinophil EoL-1 cell lines were cultured in the presence of GGE (100 μg/mL) or purified compounds from GGE for 1 h before stimulating with thrombin (50 U/mL) for 24 h. Control cells were treated with solvent alone. After 24 h, the supernatants and cells were collected to measure cytokine expression by immunoassays or RT-PCR.
In separate experiments, the anti-inflammatory potency of GGE was compared with the extract of a different mushroom (Agaricus blazei Murrill), which has been also shown to have anti-inflammatory property. HMC-1 cells were cultured in the presence of the same concentrations of each mushroom extract for 1 h before stimulation with thrombin (50 IU), and then, the mRNA expression of inflammatory cytokines was evaluated after 24 h of culture.
To evaluate the inhibitory effect of the three newly isolated sterols (gargalols) from G. gargal Singer on mast cells and eosinophils, HMC-1 mast cells and Eol-1 eosinophils were cultured in the presence of each gargalol compound (GC1, GC2, GC3) for 1 h before stimulation with thrombin and evaluation of the expression of inflammatory cytokines after 24 h.
Statistical analyses
Data are presented as mean ± standard error of the mean (SEM). Statistical differences were calculated by analysis of variance with post hoc analysis using Fisher's protected least significant difference test. All statistical analyses were performed using the StatView 5.0 package (Abacus Concepts, Berkeley, CA, USA). A P < .05 was considered as statistically different.
Results
GGE attenuates airway hyperresponsiveness
Airway hyperresponsiveness, as measured by the enhanced pause (Penh), was significantly increased in sensitized mice fed with normal food (normal food/OVA) compared with their counterpart mice fed with GGE (GGE food/OVA) and with nonsensitized mice (normal food/SAL, GGE food/SAL). No significant difference in airway hyperresponsiveness was observed between normal food/SAL and GGE food/SAL groups (Fig. 1A).
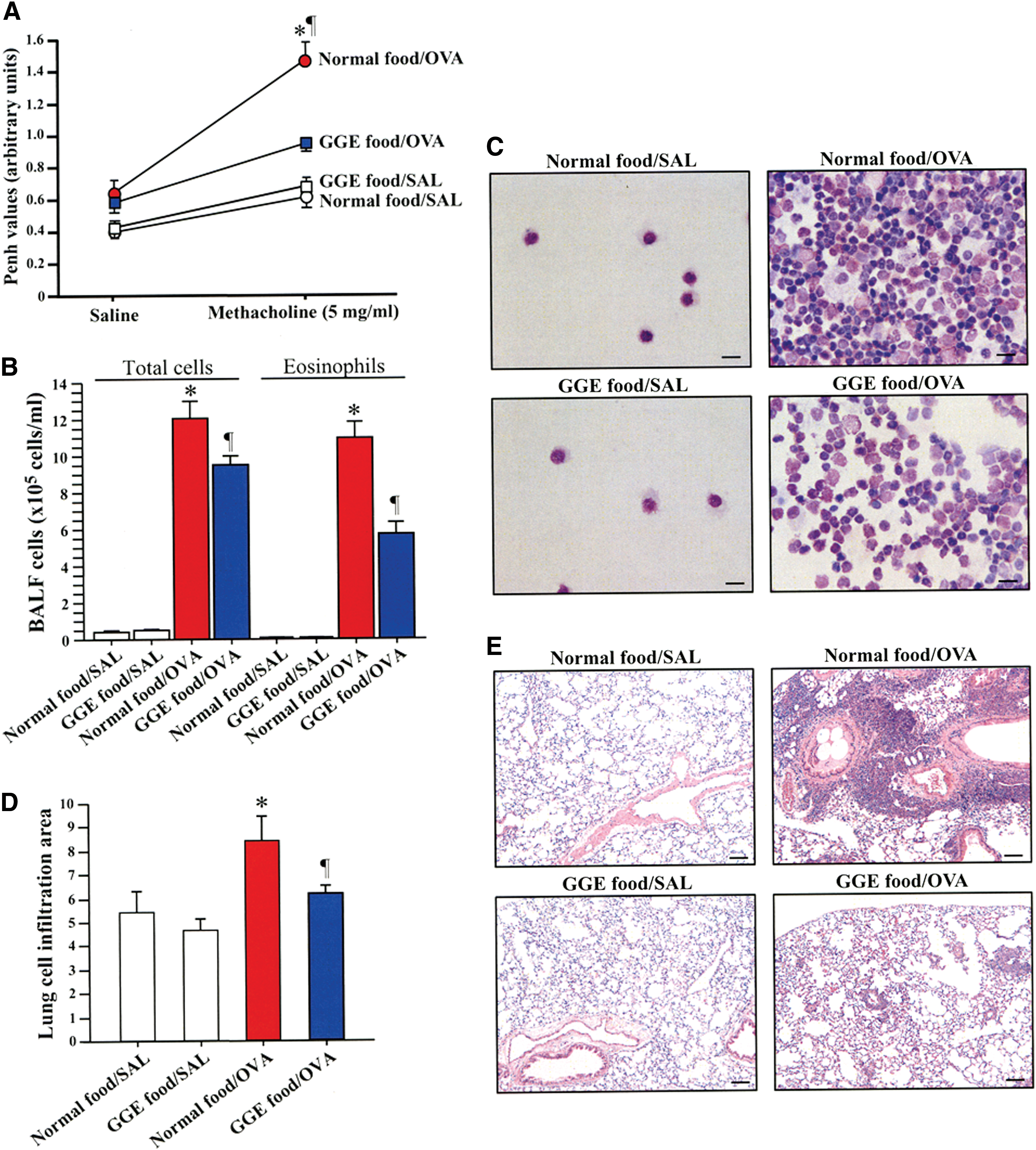
GGE protects against airway hyperresponsiveness and lung inflammation. Airway hyperresponsiveness was measured by enhanced pause (Penh;
GGE reduces lung inflammation
The total number of cells, the differential count of eosinophils in BALF samples, and the grade of cell infiltration in the lungs as detected by H&E staining of lung tissue were significantly reduced in GGE food/OVA mice compared with normal food/OVA mice (Fig. 1B–E). No significant difference in the total number of cells, in differential count of eosinophils, or in lung cell infiltration was observed between the normal food/SAL and GGE food/SAL groups.
Reduced levels of inflammatory cytokines and IgE in allergic mice receiving GGE
The BALF concentration of IL-13 tended to be low in sensitized GGE food/OVA mice compared with sensitized normal food/OVA mice (Fig. 2A). The plasma levels of IL-4 and IL-5 were significantly elevated in the normal food/OVA group compared with levels in nonsensitized normal food/SAL and GGE food/SAL groups, but they were significantly reduced in sensitized GGE food/OVA group compared with the normal food/OVA group (Fig. 2A). The lung tissue levels of IL-5, IL-13, and MCP-1 and the plasma and lung tissue levels of IgE were significantly decreased in the GGE food/OVA group compared with the normal food/OVA group (Fig. 2B, C). No significant differences were observed between normal food/SAL and GGE food/SAL groups.
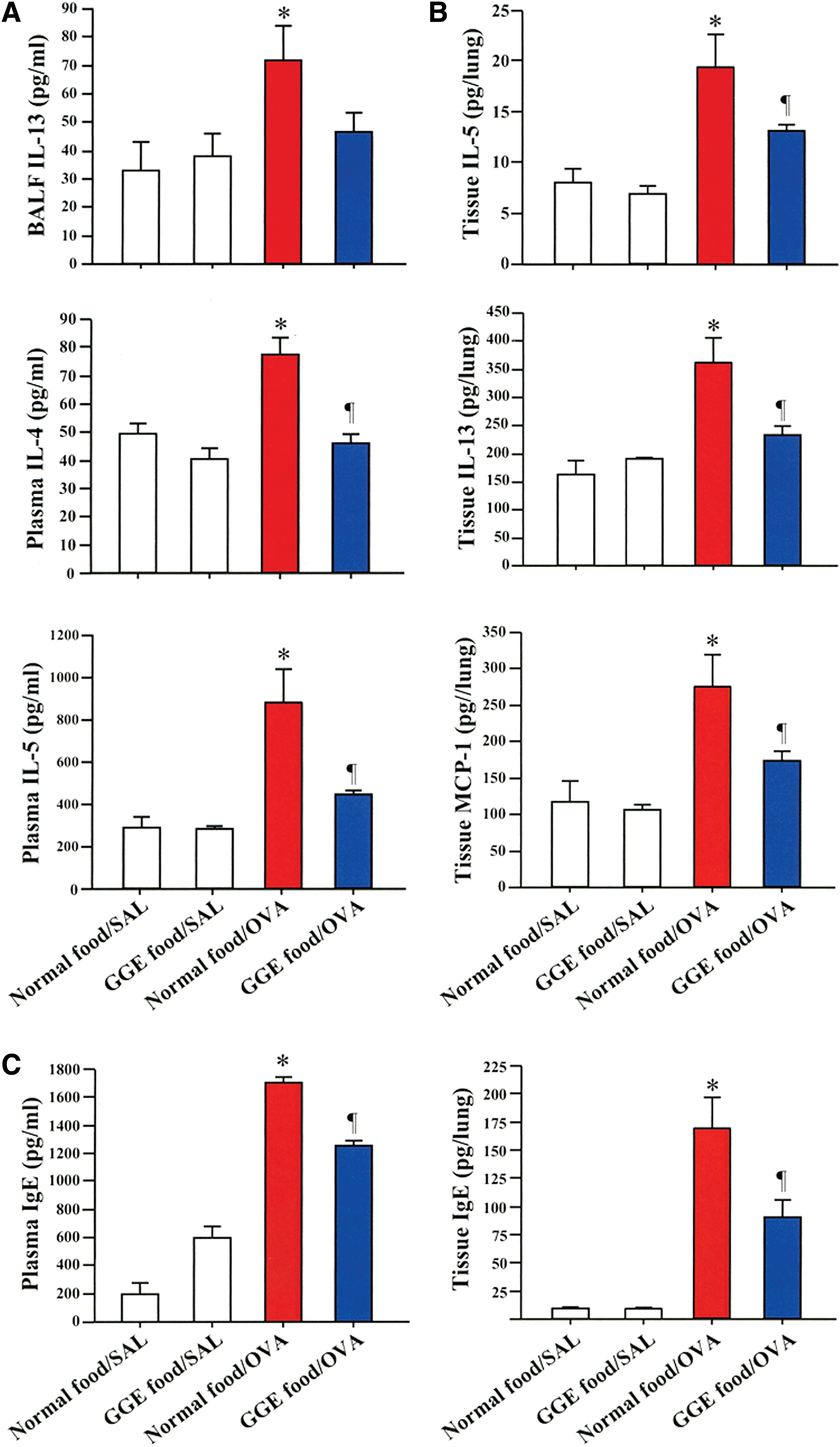
Reduced levels of cytokines and IgE after GGE treatment. The concentrations of cytokines
GGE increases the population of regulatory CD4+ T cells
To determine the effects of GGE on regulatory T cell population, spleen cells were collected from each mouse of all groups and analyzed using flow cytometry. The percentage of CD4+ CD25+ T cells, which mainly comprise regulatory T cells, was significantly increased among spleen cells isolated from GGE food/SAL and GGE food/OVA mice compared with spleen cells isolated from their counterpart normal food/SAL and normal food/OVA mice (Fig. 3A, B).

The regulatory T cell population is increased by GGE. Spleen cells were isolated and treated with anti-CD4+ and anti-CD25+ antibodies, and the number of Tregs was assessed by flow cytometry and expressed as percentage of total cells
The BALF concentration of IL-10, a cytokine that promotes differentiation of regulatory T cells, was significantly elevated in allergic mice treated with baits containing GGE (GGE food/OVA) compared with their counterpart mice fed with normal baits (normal food/OVA; Fig. 3C). No significant difference was found between nonsensitized normal food/SAL and GGE food/SAL groups.
GGE suppresses mast cell activation
To further investigate the inhibitory mechanism of GGE on allergic reactions, we examined its effects on the production of proinflammatory cytokines from thrombin-stimulated mast cells. The mast cell line HMC-1 was treated with GGE (100 μg/mL) and/or thrombin (50 U), and the expression of cytokines was monitored by immunoassays and reverse transcription-polymerase chain reaction (RT-PCR). The concentration of tumor necrosis factor (TNF)-α in the cell supernatant was significantly decreased in the presence of 100 μg/mL or more concentrations of GGE compared to control cells (Fig. 4A). Culture of mast cells in the presence of 100 μg/mL GGE also significantly decreased the concentration of IL-8, MCP-1, IL-5, and osteopontin (Fig. 4B) compared with control cells. The mRNA expressions of IL-5 and IL-13 were significantly reduced in cells cultured in the presence of GGE compared with control cells (Fig. 4B).

Suppression of mast cell activation by GGE. Human HMC-1 mast cell lines were cultured in the presence of GGE (100 μg/mL) for 1 h before addition of thrombin (50 U/mL) for 24 h. Control cells were treated with solvent alone. The expression of cytokines was evaluated by immunoassays and by RT-PCR
Potent anti-inflammatory effect of GGE compared with A. blazei Murrill extract
Previous studies have shown that A. blazei extract (ABE) also has anti-inflammatory activity under allergic conditions. 17,18 Therefore, we compared the anti-inflammatory potency of similar concentrations (100 μg/mL) of GGE and ABE in HMC-1 mast cell lines stimulated with thrombin. The mRNA expressions of MCP-1 and IL-13 were significantly inhibited in cells cultured in the presence of GGE but they were not significantly inhibited in the presence of ABE compared with cells stimulated with thrombin alone (Supplementary Fig. S1). The mRNA expression of IL-5 was significantly decreased by both GGE and ABE compared to cells stimulated with thrombin alone, but the IL-5 mRNA expression was significantly reduced in cells treated with GGE compared with cells treated with ABE (Supplementary Fig. S1).
Inhibitory effect of gargalols (GC1, GC2, GC3) on mast cells and eosinophils
The concentration of MCP-1 was significantly reduced and the concentrations of IL-13 and IL-8 tended to be lower in the supernatant of thrombin-stimulated HMC-1 mast cells cultured in the presence of GC1 (Supplementary Fig. S2). Changes in the concentration of cytokines were less pronounced in cells treated with GC2 or GC3 (data not shown).
The concentration of IL-13 was significantly decreased and that of IL-8 tended to be low in cell supernatants from Eol-1 cells cultured in the presence of GC1 compared with cells cultured in the presence of thrombin alone (Supplementary Fig. S3A). The mRNA expressions of both IL-13 and IL-8 were significantly decreased in Eol-1 cells treated with GC1 compared with cells treated with thrombin alone (Supplementary Fig. 3B).
Discussion
The results of this study show that extract from G. gargal increases the population of regulatory T cells, attenuates hyperresponsiveness and Th2-mediated allergic responses in a mouse model of bronchial asthma, and inhibits secretion of proinflammatory cytokines from mast cells and eosinophils.
Inflammation in asthma
Bronchial asthma is characterized by airway inflammation, bronchoconstriction, and excessive response to external stimuli or hyperresponsiveness. 5 The inflammatory component is mainly mediated by activation of Th2 cells leading to exaggerated extracellular release of cytokines and chemokines, including IL-4, IL-5, IL-13, and chemokines such as MCP-1 and elevated levels of allergen-specific IgE. 6 This proinflammatory environment results in increased lung recruitment of activated eosinophils and allergen/IgE-mediated increased activation of mast cells and basophils. 5 In the present study, we showed that mice with allergic bronchial asthma fed with extracts prepared from the medicinal mushroom G. gargal have significantly less airway hyperreactivity, reduced lung level of Th2 cytokines and MCP-1, with pronounced reduction in the number of eosinophils infiltrating the lungs, compared with allergic mice fed without the mushroom extract. Overall, these observations suggest a beneficial effect of G. gargal supplement against allergic bronchial asthma.
Regulatory T cells in allergy
We measured changes in the systemic regulatory T cell population to search for the possible mechanism for the suppressive effect of the mushroom extract. Regulatory T cells comprised a group of T cells that are essential for regulation of the immune system activation to prevent the development of immune-mediated diseases (allergy, autoimmunity) and unnecessary expansion of effector T cells. 19 Differentiation and increase in the population of regulatory T cells are regulated by both IL-10 and transforming growth factor-β1. 20 Increased numbers of regulatory T cells in the airway and peripheral blood are associated with amelioration of airway hyperresponsiveness and airway inflammation in chronic bronchial asthma. 21 In this study, we showed that ingestion of GGE significantly increased the concentration of IL-10 and the number of CD+/CD25+ T cells in the lungs of mice with allergen-mediated asthma compared with untreated mice, suggesting the possible mediation of regulatory T cells in the protective activity of GGE against allergic asthma. Enhancement of dendritic cell-induced differentiation of T cells to CD4+CD25+Foxp3+ T cells by GGE reported in a previous study supports the role of regulatory T cells in the beneficial effect of G. gargal in allergic bronchial asthma. 10
Mast cells in allergic responses
A type I reaction, including allergic asthma and hay fever, involves the IgE-mediated activation of mast cells and basophils in affected tissues leading to the release of Th2 cytokines (IL-5, IL-13), chemokines (IL-8, MCP-1), and bronchoconstricting factors (histamine, leukotrienes). 22 The release of Th2 cytokines further promotes allergic responses, while the secretion of chemokines increases the recruitment of inflammatory cells at sites of allergen entry. 5,6 Here we cultured mast cells in vitro in the presence or absence of GGE and evaluated the grade of mast cell activation by measuring the expression of cytokines. The protein secretion of TNF-α, IL-8, and MCP-1 and the mRNA expression of IL-5 and IL-13 were significantly downregulated when the cells were cultured in the presence of GGE compared with control cells. This anti-inflammatory activity of GGE was significantly stronger than the extract of A. blazei, a mushroom that has been previously reported to have antiallergic activity. Previous studies showing that a component of G. gargal can activate the 5′ adenosine monophosphate-activated protein kinase, and that the 5′ adenosine monophosphate-activated protein kinase negatively regulates cell signaling in mast cells suggest the potential role of this protein kinase in the suppressive activity of GGE. 23,24 Overall, these observations support the hypothesis that GGE protects against allergic asthma by inhibiting the activation of mast cells.
Active components in G. gargal
Many compounds found in mushrooms have been reported to exhibit immune-modulatory, anticarcinogenic, antiviral, antioxidant, and anti-inflammatory properties. 25 –29 For example, the extracts of A. blazei and Pleurotus eryngii have been shown to protect against allergic responses in in vivo and in vitro experimental models. 17,30 GGE contains several components, including β-glucan, polyphenols, and flavonoids. β-glucan can inhibit the expression of cytokines (TNF-α, IL-6) and chemokines (MCP-1) and flavonoid glycosides can prevent allergic responses and the production of nitric oxide synthase and cytokines. 31 –33 Here we isolated three novel sterols (GC1, GC2, GC3) from G. gargal, assessed their inhibitory properties, and found that one of the compounds (GC1) significantly suppresses the secretion of IL-13, IL-8, and MCP-1 from mast cells and the expression of IL-13 and IL-8 from eosinophils. In addition to supporting the protective activity of GGE, these findings indicate that sterol components of GGE are also important for the protective activity against allergy.
In brief, the results of this investigation demonstrated that extract from G. gargal increases the population of regulatory T cells, inhibits the expression of proinflammatory cytokines, and attenuates inflammation in an experimental model of allergic bronchial asthma, and that the sterol component of the mushroom can directly suppress the secretory activity of mast cells and eosinophils.
Footnotes
Acknowledgments
This research was supported, in part, by grants from the Ministry of Education, Culture, Sports, Science and Technology of Japan (Kankenhi No. 24591128) and, in part, by a research grant from the Iwadekin Research Institute of Mycology. The funders had no role in the study design, data analysis, decision to publish, or preparation of the manuscript.
Author Disclosure Statement
E.H., T.M., and T.S. are employees of Iwade Research Institute of Mycology. Other authors declare no financial conflicts of interest regarding this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.