
Abstract
Luteolin (LU) is a widely distributed flavonoid with multitarget effects. The objective of this study was to determine whether LU could reduce the ischemia–reperfusion injury of the spinal cord (SCII) in a rat model. Forty-eight rats were divided into four groups: sham, SCII, SCII+L-LU (50 mg/kg), and SCII+H-LU (100 mg/kg). Abdominal aortic occlusion was carried out for 40 min in all groups. Hindlimb motor functions were evaluated using the Tarlov scoring system. Nissl and terminal deoxynucleotidyl transferase-mediated dUTP biotin nick end labeling (TUNEL) staining were used to detect cell survival and apoptosis in the spinal cord. Spinal cord samples were taken for determination of malondialdehyde, xanthine oxidase, superoxide dismutase, and glutathione peroxidase activities. The levels of tumor necrosis factor-α, interleukin (IL)-1β, and IL-18 were assessed using ELISA kits to examine the inflammatory responses in the spinal cord. Western blot analysis was used to examine the expression of nuclear factor erythroid 2-related factor (Nrf2) and nod-like receptor pyrin domain-containing 3 protein (NLRP3) levels. We found that LU pretreatment significantly improved the locomotor function of rats after SCII, increased neuron survival, and inhibited apoptosis in the spinal cord. Furthermore, the oxidative stress and inflammatory response were significantly suppressed upon treatment with LU. Finally, LU upregulated Nrf2 levels and downregulated NLRP3 protein expression in SCII tissues. Thus, LU exhibited a neuroprotective effect following SCII by alleviating oxidative stress and inhibiting inflammatory responses and cell apoptosis. The possible mechanism may be related to the activation of Nrf2 and inhibition of NLRP3 inflammasome pathway.
Introduction
S
Oxidative stress, a mediator of the secondary injury process that occurs following primary mechanical injury to the spinal cord, destroys the balance of the redox states and contributes to the development of SCII. 3 Extensive evidence demonstrated that the predominant form of oxygen radical-induced oxidative damage that occurs in injured nervous tissue is lipid peroxidation. 4 Moreover, large amounts of proinflammatory cytokines, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-18, can induce neuronal apoptosis and even death. 5,6 Recent studies have shown that mitochondrial production of reactive oxygen species (ROS) and inflammation may be closely linked through a redox sensor known as nod-like receptor pyrin domain-containing 3 protein (NLRP3). 7 The maturations and secretions of pro-IL-1β and pro-IL-18 require the activation of caspase-1 that, together with the activation of the NLRP3 inflammasome, induces the inflammatory cascade and subsequently recruits apoptosis-associated proteins. 8 Furthermore, apoptosis is another prominent feature leading to neuronal cell death and hampering the neurorepair process subsequent to the SCII state. 9 Unfortunately, a proven effective therapy for neuroprotection following SCII is still not available at present.
Luteolin (LU), a widely distributed flavonoid with multitarget effects, plays an important role in antioxidant, 10,11 anti-inflammatory, 12,13 and neuroprotective properties 14 of disease. LU was found to not only improve the host system by modifying the levels of enzymatic antioxidants and lipid peroxides but also reduced the levels of glutathione peroxidase (GSH-Px) and the inflammatory cytokine IL-2 in plasma or liver tissue of a mouse liver carcinogenesis model. 15 In addition, LU could activate nuclear factor erythroid 2-related factor (Nrf2)-antioxidant response element-mediated gene expression and induce anti-inflammatory activities in human hepatoma HepG2 cells. 16 Moreover, LU prevented H2O2-induced apoptosis in H9C2 cells through modulating oxidative stress and apoptosis-associated signaling pathways. 17 However, the neuroprotective effects of LU on SCII have not yet been systematically investigated.
This study investigated whether pretreatment with LU reduces SCII-associated inflammation and apoptosis, as well as neurological, histopathological, and oxidative damage, using a rat infrarenal abdominal aortic clamping model. We also examined the contribution of Nrf2 and NLRP3 protein expression.
Materials and Methods
Animals
Male Sprague-Dawley rats (180–200 g) were obtained from the Experimental Animal Center of Hebei Medical University (Animal licenses no.: SCXK [Hebei 2013-1-003]; animal certification no.:1301020). Animals were allowed to acclimatize for 1 week before the experiments. They were housed in groups of four per cage in a room that was maintained as a standard light-controlled room (12-h light/12-h dark cycle) and a constant temperature (25°C ± 1°C) and humidity (50–60%). All rats were allowed free access to food and water. All experiments conformed to the National Institutes of Health guidelines for the Care and Use of Laboratory Animals and with the European Communities Council Directive of 24 November 1986 (86/609/EEC). Every effort was made to minimize the number of animals used and their suffering.
Materials
LU (purity >98%, CAS: ZL201125; Nanjing Zelang Biotechnology Company, Jiangsu, China) was resuspended in 0.05 g/mL sodium carboxymethylcellulose (CMC-Na) solution. Thionine reagent (CAS: R310360; Sigma-Aldrich, Co., St. Louis, MO, USA) was dissolved in 0.01 g/mL normal saline. All treatments were carried out between 08:30 and 10:30 a.m. to minimize the effects of circadian rhythm. Malondialdehyde (MDA), xanthine oxidase (XO), superoxide dismutase (SOD), and GSH-Px detection kits were from Nanjing Jiancheng Bioengineering Institute (Nanjing, China).
Study groups
Forty-eight rats were randomly allocated to the following four groups (n = 12/group): (1) Sham, in which rats received daily intragastric injections of the vehicle (CMC-Na, 5 mL/kg) for 14 days, after which they underwent laminectomy where their abdominal aorta was exposed, but they did not undergo aortic occlusion clamp; (2) SCII, in which rats received daily intragastric injections of vehicle for 14 days before transient global spinal cord ischemia was induced; (3) SCII+L-LU, in which rats received daily intragastric injections of 50 mg/kg LU for 14 days before SCII; and (4) SCII+H-LU, in which rats received daily intragastric injections of 100 mg/kg LU for 14 days before SCII. All cross-clamped groups were reperfused after spinal cord ischemia for 40 min, before the occlusion clamp was removed. 18
Rat model of SCII
SCII was induced in rats as previously described. 18 Rats were anesthetized by intraperitoneal injection of 3.5 mL/kg 6% chloral hydrate (Sigma-Aldrich, Co.) before the surgical procedure. Core body temperature was maintained at 36°C ± 0.5°C. Animals were placed in a supine position for the surgery. They were shaved from abdomen to leg, and the surgical area was marked, cleaned, and draped. After sterile preparation, a 10-cm midline incision was made, and the abdominal aorta was exposed using a conventional approach. Before clamping, heparin (130 U/kg) was administered intravenously for 5 min for anticoagulation. The aorta was clamped ∼1 cm below the left renal artery using two bulldog clamps. SCII was induced by occlusion of the abdominal aorta (between the renal arteries and iliac bifurcations) for 40 min. When the pulsation of the femoral artery disappeared, the clamps were removed and return of the aortic pulse was verified. The wound was closed in layers with 3-0 silk sutures after surgical and ischemia interventions. Subsequently, rats were given free access to food and water.
At 48 h after reperfusion, rats were euthanized by deep anesthesia and transcardiac perfusion with about 250 mL ice-cold normal saline was carried out. The lumbar spinal cord (L2–5 segments) of rats was quickly removed. The samples were carefully dissected and divided into two sections. One of the sections was used to observe cell survival and apoptosis by Nissl and Terminal deoxynucleotidyl transferase-mediated dUTP biotin nick end labeling (TUNEL) staining, respectively. The other parts of samples were flash frozen in liquid nitrogen and then stored at −80°C for further analyses, including the detection of oxidative stress and inflammatory factors using ELISA and protein expression of Nrf2 and NLRP3 using Western blot.
Evaluation of neurological status
After reperfusion, two independent study observers blinded to the treatment groups of the animals evaluated the hindlimb motor function before surgery, and 24 and 48 h after surgery according to the Tarlov scoring system. 18 A score between 0 and 4 was assigned to each animal as follows: 0 = spastic paraplegia and no movement of the lower limbs, 1 = spastic paraplegia and slight movement of the lower limbs, 2 = good movement of the lower limbs, but unable to stand, 3 = able to stand, but unable to walk normally, and 4 = complete recovery and normal gait–hopping.
Nissl staining
After neurological evaluation, the rats were decapitated and a 5-mm-thick coronal section was fixed in 4% paraformaldehyde for 72 h, dehydrated in alcohol, cleared with xylene, and embedded in paraffin. The sections were sliced at 5-μm thickness and were Nissl stained with 1% thionin. In every fifth slice (three slices per animal), we counted the number of surviving intact cells per mm length of the spinal cord. The results were expressed as the average number of cells within each frame per section. To reduce counting bias, cell counting was performed by two independent investigators blinded to treatment history using high magnification light microscopy. 10
TUNEL assay
Spinal cord slices were embedded in paraffin as previously described 18 and 5-mm-thick sections were deparaffinized in 100% xylene and a descending ethanol series. The sections were stained with hematoxylin and processed. DNA fragments were determined using an ApopTag in situ apoptosis detection kit according to the manufacturer's instructions (ApopTag; Oncor, Gaithersberg, MD, USA). The DNA nick was labeled and the sections were counterstained with hematoxylin. The labeled apoptotic neurons with diaminobenzidine were defined as TUNEL positive and apoptotic index (AI) was used to quantify the number of TUNEL-positive cells. Subsequently, five nonadjacent fields in each section were randomly chosen to count the positive cells and total number of neurons. The AI was calculated as AI = (number of positive cells/total number counted) × 100%.
Biochemical examination
Spinal cord tissues were homogenized in normal saline and centrifuged at 4000 g for 20 min. The upper supernatants were collected and stored at −80°C until further processing.
Tissue MDA assay
MDA levels were determined by a method based on a reaction with thiobarbituric acid, as described previously by Ohkawa et al. 19 The principle of the method is based on the spectrophotometric measurement of the color that occurs during the reaction of thiobarbituric acid with MDA. The concentrations were expressed as nmol/mg protein.
Tissue XO assay
The XO activity was determined spectrophotometrically by the method of Hashimoto based on the formation of uric acid from xanthine at 293 nm. 20
Tissue SOD assay
Total SOD activity was determined according to the method described by Sun et al. 21 The SOD activity was expressed as U/mg protein.
Tissue GSH-Px activities
The GSH-Px activity was determined by a photometric kinetic measurement using a GSH Assay Kit following the oxidation of nicotinamide adenine dinucleotide phosphate spectrophotometrically at 340 nm. 6 The GSH-Px concentration was calculated as U/g protein.
Determination of TNF-α, IL-1β, and IL-18 levels
TNF-α, IL-1β (E-Bioscience, Vienna, Austria), and IL-18 (Boster Biological Technology, Pleasanton, CA, USA) levels were determined by ELISA using specific kits and expressed as pg/mg protein.
Determination of the expression of Nrf2 and NLRP3 proteins by Western blot
The frozen spinal cord samples were processed and proteins were extracted (Applygen Technologies, Beijing, China). After determining protein concentration, equal amounts of protein per group were resolved by 10% sodium dodecyl sulfate– polyacrylamide gel electrophoresis and transferred to a PVDF membrane (Millipore, Billerica, MA, USA), which was blocked in 5% skim milk for 1 h at room temperature. PVDF membranes were subsequently incubated with primary antibodies overnight at 4°C: anti-Nrf2 (ab137550, 1:500; Abcam, Cambridge, MA, USA), anti-NLRP3 (ab214185, 1:1000; Abcam), and anti-β-actin (ab8227, 1:1000; Abcam). After washing in phosphate-buffered saline with 0.1% Tween-20 (PBST), membranes were incubated with a peroxidase-conjugated goat anti-rabbit IgG secondary antibody (1:1500; Millipore) for 2 h at room temperature. Detection of proteins was performed and the levels were analyzed by imaging software (Bio-Rad Co. Ltd., Hercules, CA, USA).
Statistical analysis
All data are expressed as the mean ± standard error of the mean for each group using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA). The significance of differences between the groups was assessed by one-way ANOVA followed by Tukey's test for multiple comparisons. P < .05 was considered significant.
Results
Effect of LU on hindlimb motor function of SCII rats
To evaluate the effect of LU on functional recovery in SCII rats, hindlimb motor function was recorded using the Tarlov scoring system (Fig. 1). Compared with before surgery, the scores obtained 24 and 48 h after reperfusion did not significantly differ (P > .05) in the sham group, but rats in the SCII group presented with severe neurological deficits, including paraplegia. Compared with the SCII group, the score of hindlimb function in the SCII+LU groups was higher 48 h after reperfusion than 24 h (P < .05). After 48 h, the hindlimb function of rats in the SCII+LU groups improved dose dependently compared with that of SCII group rats (both P < .05).

Neurological deficit scores of hindlimbs' motor function at different time points. The function of hindlimb recovery was assessed from before surgery to 48 h after SCII using the Tarlov scoring system. The scores in the SCII group markedly declined at 24 h after reperfusion. In SCII+LU groups, the hindlimb dysfunction was dose-dependently ameliorated not only at 24 h but also at 48 h after reperfusion. Values are mean ± SEM. *P < 0.05 versus Sham group, # P < 0.05 versus SCII group, & P < 0.05 versus SCII+L-LU group. LU, luteolin; SCII, ischemia–reperfusion injury of the spinal cord; SEM, standard error of the mean.
Effect of LU on neuron survival and cell apoptosis in the spinal cord of SCII rats
To observe a possible neuroprotective effect of LU, Nissl and TUNEL staining were performed (Fig. 2). In the sham group, positive cells in the Nissl-stained sections were abundantly detected and a large centrally located nucleus was found in the ventral horn of the spinal cord 48 h after reperfusion (Fig. 2A). In this group, the mean number of Nissl-positive neurons was 22.6 per section in the spinal cord (Fig. 2B). In contrast, the Nissl-positive neurons from the SCII group showed morphological deformation, including cytomembrane blurring, shrunken cell bodies, and pyknotic nuclei, which was significantly decreased compared with the sham group. Compared with the SCII group, the number of damaged neurons was markedly reduced in both SCII+LU groups. Hence, LU pretreatment resulted in significant protective effects on neuron survival in the spinal cord of SCII rats.
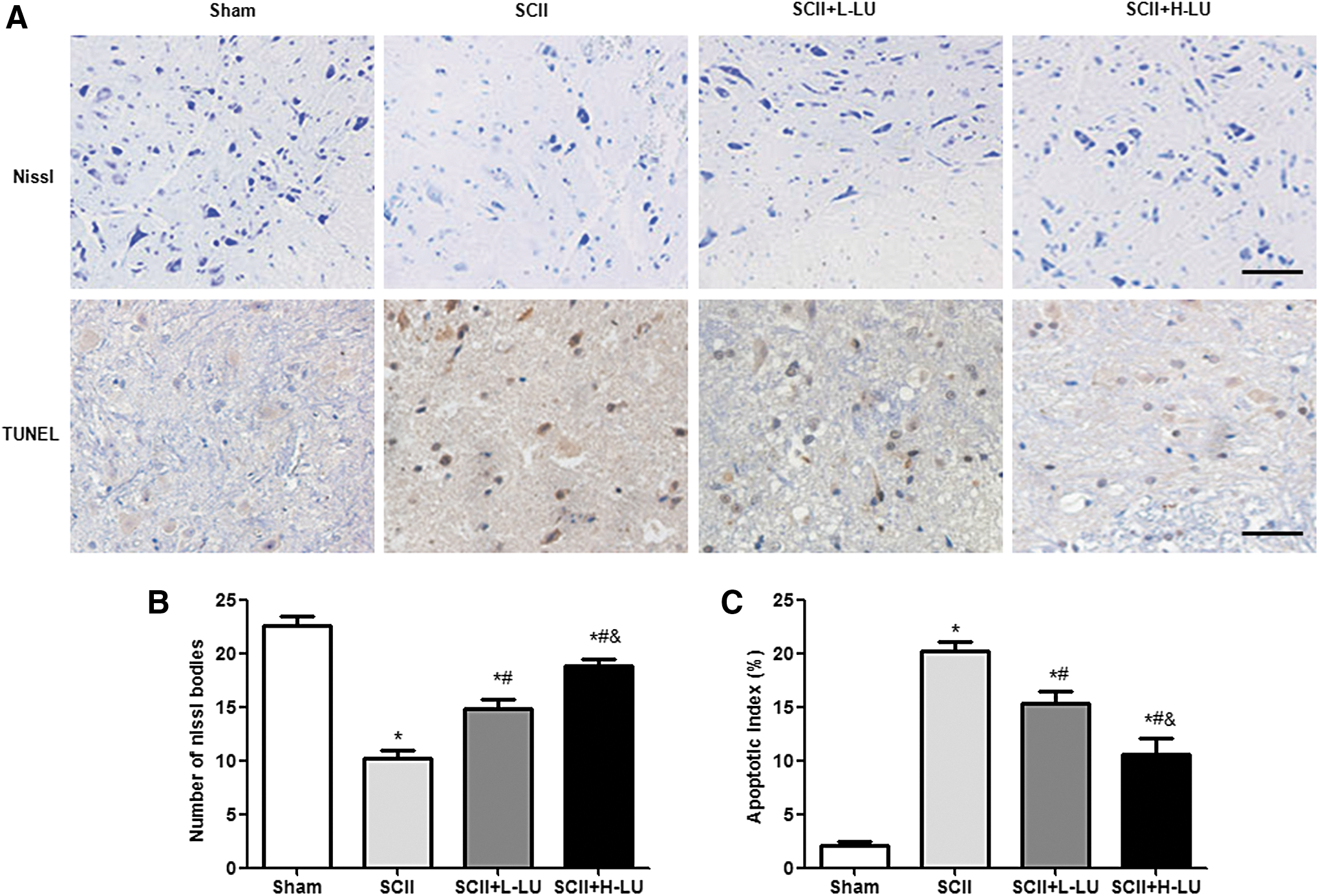
Effects of LU on neuron survival and cell apoptosis in the spinal cord after SCII.
As shown in Figure 2, there were almost no TUNEL-positive cells in the sham group, while numerous brown-stained TUNEL-positive cells were observed in the SCII group (P < .05; Fig. 2C). Moreover, LU pretreatment significantly and dose-dependently reduced the number of apoptotic cells in both groups of SCII rats.
Effect of LU on oxidative stress in the spinal cord of SCII rats
To evaluate how LU exerts neuroprotective effects, the oxidative (MDA and XO) levels and antioxidative (SOD and GSH-Px) activities in local spinal cord tissue were detected. In the sham group, MDA and XO levels in spinal cord homogenates were 4.15 nmol/mg protein and 3.65 mU/mg protein, respectively. In the SCII group, the MDA level was robustly increased by 11.78 nmol/mg protein; the XO level was 9.65 mU/mg protein. In the SCII+LU groups, the MDA and XO levels were significantly decreased in a dose-dependent manner relative to the SCII group (Fig. 3), thereby mitigating the oxidative stress resulting from SCII-induced injury.

Effects of LU on oxidative stress at 48 h after SCII. MDA
The highest SOD and GSH-Px activities were observed in the sham group. As expected, there was a significant decrease in the amount of SOD and GSH-Px in the SCII groups compared with the sham group, with a similar trend observed for SOD and GSH-Px activities. Interestingly, compared with the SCII group, the activities of SOD and GSH-Px were significantly higher in the SCII+LU groups, although there was no significant difference between the low dose and the high dose of LU pretreatment in SOD activity (P > .05).
Effect of LU on inflammatory responses in the spinal cord of SCII rats
Proinflammatory cytokines were measured to evaluate the anti-inflammatory effect of LU in SCII rats. As shown in Figure 4, the levels of TNF-α, IL-1β, and IL-18 in the spinal cord were relatively low in the sham group, whereas a marked increase was observed in the SCII group relative to the sham group. However, LU pretreatment dose-dependently prevented the secretion of these cytokines compared with the SCII group.

Effects of LU on proinflammatory cytokines at 48 h after SCII. The levels of TNF-α
Effects of LU on Nrf2 levels and NLRP3 inflammasome activation in the spinal cord 48 h after SCII
As shown in Figure 5, there was a significant downregulation of Nrf2 protein expression and an upregulation of NLRP3 protein expression in the SCII groups when compared with the sham group. Interestingly, LU pretreatment dose-dependently increased Nrf2 level and reduced NLRP3 level in the SCII +LU groups compared with the SCII group (P < .05).
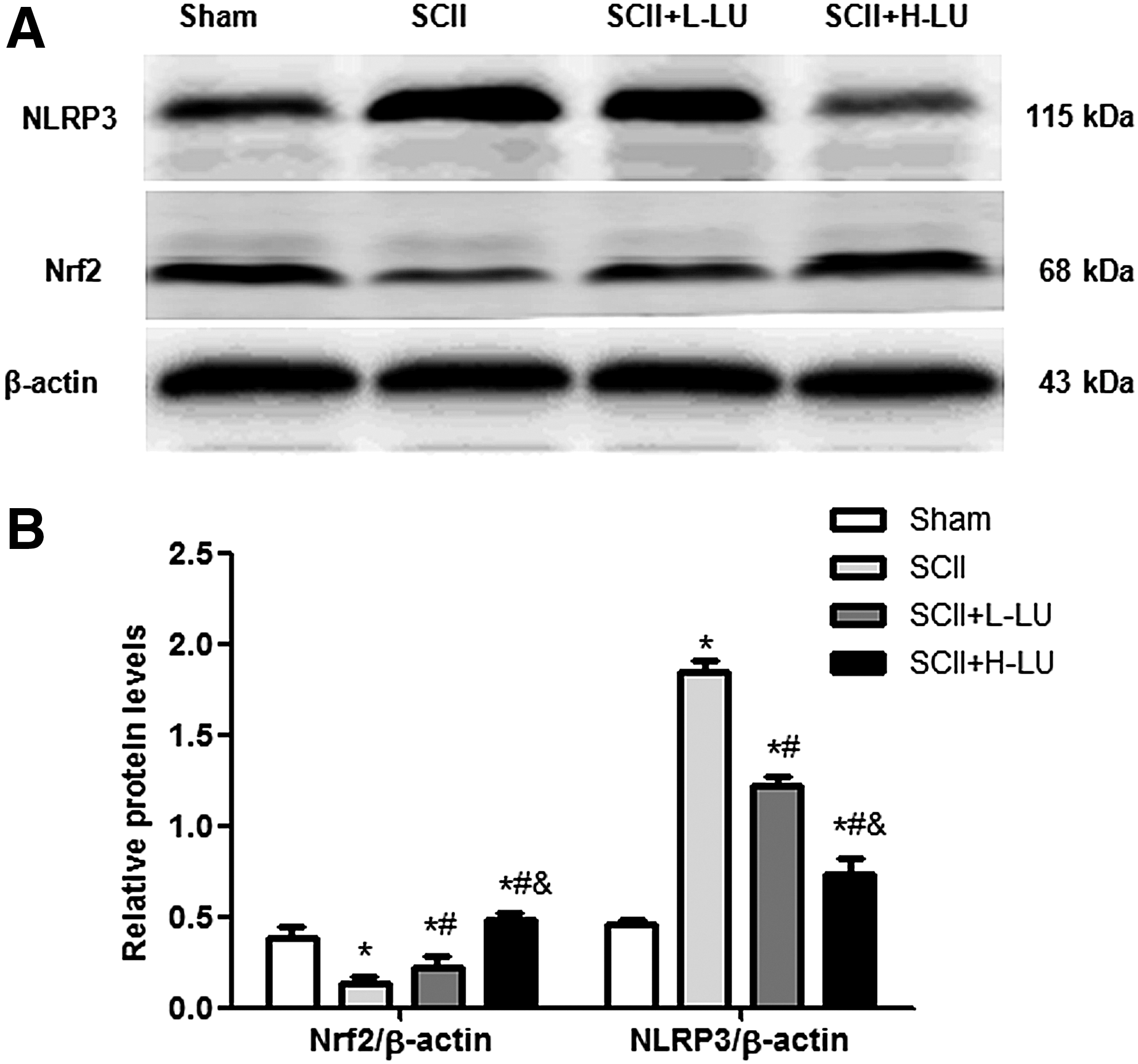
Effects of LU on Nrf2 and NLRP3 protein levels in the spinal cord 48 h after SCII.
Discussion
The spinal cord can easily suffer from ischemia damage since its blood supply is segmental and the collateral circulation is relatively poor. 9 SCII is mainly due to aortic cross-clamping during thoracoabdominal surgery and the subsequent recovery perfusion, which may further aggravate the damage of the spinal cord. Due to its complex pathophysiological mechanisms, oxidative stress and inflammatory processes with or without apoptosis and necrosis play a crucial role in SCII. 6,22 The cell membranes of spinal cord neurons, which are rich in lipids and unsaturated fatty acids, are more susceptible to oxygen-free radical attacks. 3,23 After the spinal cord suffered the secondary injury, the lipid peroxidation reaction was dramatically activated, whereas antioxidative activities markedly decreased. Decreasing MDA levels and increasing the SOD activity have been shown to markedly attenuate SCII. 3 In addition, an inflammatory response develops within several hours after injury and is characterized by the activation of proinflammatory cytokines and infiltration of neutrophils. 24 Numerous studies have confirmed that the proinflammatory cytokines (TNF-α, IL-1β, and IL-18) are major mediators of several central nervous system diseases. Increased expression of TNF-α, IL-1β, and IL-18 have been reported in animal models of traumatic spinal cord injury, and these cytokines also play a major role in the inflammatory response to traumatic spinal injury or in pathological mechanisms of secondary damage. 25 Therefore, the aforementioned factors may easily contribute to neuronal cell apoptosis or death during the reperfusion period. 26
LU, as a natural oxygen-free radical scavenger, has the advantage of lowering toxic adverse actions of nonnatural products. Shimoi et al. reported that LU can be absorbed after oral administration and it was absorbed more efficiently from the duodenojejunum than from the ileum. 27 Although most LU found in plasma is in the form of glucuronide and sulfate conjugates, low concentrations of free LU can be achieved in plasma after oral ingestion of this flavonoid. Furthermore, several animal experiments have shown that LU possesses a wide range of biological activities and several possible mechanisms of action in vivo, 28 which have protective effects against ischemia injury. 29 Several studies have shown that LU effectively decreased oxidative stress and inflammation in various diseases, such as hepatoma 15 and severe acute pancreatitis. 13 Zhen et al. also confirmed that pretreatment with LU rescues pentylenetetrazole-induced cognitive impairment in epileptic rats by decreasing oxidative stress and neuronal damage. 10 However, its neuroprotective effects with respect to SCII have never been investigated before. Therefore, it is necessary for us to further consider related studies on LU.
In this study, we observed that substantially fewer neurons survived in the area around the lesion in the SCII group at 48 h after reperfusion. Oxidative products (MDA and XO), proinflammatory cytokines (TNF-α, IL-1β, and IL-18), as well as cell apoptosis were dramatically elevated, which is consistent with previous reports. 6 Interestingly, the beneficial effects of LU treatment against SCII were associated with decreased levels of oxidative products (MDA and XO) and proinflammatory cytokines (TNF-α, IL-1β, and IL-18), increased activities of endogenous antioxidant enzymes (SOD and GSP-Px), as well as a reduction of spinal neuron apoptosis. Moreover, there was no significant difference in antioxidant enzymes (SOD) between doses of 50 and 100 mg/kg. The aforementioned results demonstrate that LU pretreatment is beneficial to SCII by reducing oxidative stress, inflammatory responses, and apoptosis. In addition, the pathological changes (Nissl staining and TUNEL staining) of the LU group were found to be significantly better than in the SCII group. Furthermore, LU pretreatment also significantly attenuated SCII-induced hindlimb neurological dysfunction at various time points after reperfusion. These results suggest that LU pretreatment might protect against SCII in rats.
To assess the LU effects on molecular changes associated with improved motor function of SCII, we evaluated Nrf2 and NLRP3 protein expression, which are important factors to prompt the expression of many antioxidant genes and inflammation. 5,30 Previous studies have shown that Nrf2 knockout mice had higher levels of spinal cord edema, NF-κB activation, and TNF-α production after spinal cord ischemia compared with wild-type mice. 31 Moreover, ROS upregulated NLRP3 inflammasome activation, an important proinflammatory mediator regulating the maturation and release of IL-1β and IL-18. 32 Prevention of NLRP3 inflammasome activation will lead to inhibition of ROS levels, reductions of IL-1β and IL-18 secretion, and less inflammation. 33 In this study, we also found that SCII caused Nrf2 inhibition and NLRP3 inflammasome activation in spinal cord tissue after SCII and LU pretreatment significantly increased Nrf2 expression and inhibited NLRP3 inflammasome activation. Nevertheless, the following limitations of this study should be taken into account. Due to the relatively small number of rats, we lack a dynamic observation for the aforementioned factors. Moreover, only two different doses of LU were chosen in the study.
In conclusion, these results indicated that LU may be a potentially neuroprotective therapeutic agent for SCII through the reduction of oxidative stress, inflammatory response, and cell apoptosis 48 h after SCII. Furthermore, neurological measurements supported these results. Importantly, the possible underlying molecular mechanism may be related to the activation of Nrf2 and inhibition of the NLRP3 inflammasome pathway. However, further studies are needed to show the therapeutic mechanisms and the correct dose of LU necessary for maximum benefit.
Footnotes
Acknowledgment
This work was supported by the Health Department Science Foundation of Hebei Province (grant no. 20170101) for data collection.
Author Disclosure Statement
No competing financial interests exist.