
Abstract
Chemotherapeutics, including 5-fluorouracil (5-FU), often damage the intestinal mucosal barrier and cause intestinal mucositis (IM). Supplementation with immunoregulatory polysaccharides from Crassostrea hongkongensis has been shown to positively influence the effectiveness and toxicity of 5-FU. Therefore, we studied the effects of oyster polysaccharides on 5-FU-induced intestinal mucosal damage in rats. The C30–60% ethanol-precipitated fraction of polysaccharides promoted IEC-6 cell proliferation and exhibited a maximal effect at a 0.0195 mg/mL concentration. Moreover, treatment with C30–60% polysaccharide-based nutrition formula (OPNF) partially prevented the 5-FU-induced degenerative changes in the histology and ultrastructure of small intestinal mucosa. In addition, the endotoxin level of rats fed with 5-FU and OPNF decreased to the normal control level. Furthermore, the 5-FU-induced increase of proinflammatory cytokine interleukin (IL)-2 and the decrease of anti-inflammatory cytokine IL-10 level in the peripheral blood were significantly attenuated by OPNF administration. In conclusion, Oyster C30–60% polysaccharides can ameliorate 5-FU-induced IM by partially preventing mucosal damage, reducing inflammation, and promoting immunity.
Introduction
T
However, clinical studies have shown that immunonutrition-enriched enteral nutrition formulas (NFs) can improve the prognosis and quality of life during cancer chemotherapy 5 by reducing the inflammatory response of gut mucosa, 6 reducing damage to the permeability of the intestinal mucosa, 7 improving immune function, and preventing postoperative complications. 8 –10 Therefore, it is essential to develop immunonutrition-based enteral nutrition support to improve the efficacy of antitumoral therapy and prevent patients from developing the unwanted side effects of chemotherapy.
Intestinal mucosal tissue contains the largest portion of the immune system present in the human body. The intestinal immune system must permit the absorption of nutrients, while protecting against pathogen invasion by repairing injuries to the intestinal mucosa. 11 A meta-analysis revealed that immunoenhancing enteral nutrition is the best available option to reduce the incidence of complications after gastrointestinal injury. 12 Studies have found that various polysaccharides can be used as immunomodulators, stimulating the immune system to more effectively protect the body against foreign invaders.
The gut mucosal immune and other bioactive functions of dietary polysaccharides have been reported. For example, dietary squid ink polysaccharide reduced small intestinal mucosal damage induced by cyclophosphamide by increasing the secretion of secretory immunoglobulin A, 13 enhancing immune function, 14 and increasing the mRNA level of tight junction proteins. 15 Chitosan, a polysaccharide extracted from crustacean shells, contributed to the maintenance of intestinal homeostasis by enhancing the microenvironment of T helper cell type 2 (Th2)/Th3 in the mucosa. 16 However, few studies pertaining to the intestinal mucosal function of polysaccharides in enteral nutrition have been reported. For example, an enteral diet supplemented with soy polysaccharide was reported to stimulate crypt cell proliferation for accelerating recovery from intestinal mucosal damage. 17
Our preliminary study revealed that supplementation with immunoregulatory polysaccharides from Crassostrea hongkongensis enhanced the inhibition of tumor growth and improved the leukocyte and lymphocyte index in mice subjected to 5-FU chemotherapy. 18 Indeed, oyster polysaccharides could be useful immune stimulants that contribute indirectly to antitumor activity by improving immunologic function. Thus, it is of interest to determine whether the beneficial influences of oyster polysaccharides on the toxicity and effectiveness of 5-FU chemotherapy are related to their protection of the intestinal mucosa. The aim of this study was to determine whether oyster polysaccharides promote the proliferation of intestinal epithelial cells and ameliorate intestinal mucosal damage by enhancing mucosal recovery, thereby reducing the inflammatory response and improving immunity in 5-FU-administered rats.
Materials and Methods
Materials
C. hongkongensis was obtained from an oyster culture farm in Hengqin, Zhuhai, China. The lyophilized voucher specimens (Voucher Specimen No. 20121014) were deposited in the South China Sea Institute of Oceanology, Chinese Academy of Sciences (SCSIO, CAS). 5-FU was obtained from Xudong Haipu Pharmaceutical Co., Ltd. (Shanghai, China). Rat small intestinal epithelial cells (IEC-6) were purchased from Yinzijing Pharmtech, Inc., Ltd. (Beijing, China).
Fractionation of polysaccharides by graded ethanol precipitation
The fractionation of oyster polysaccharides was achieved by graded ethanol precipitation as described by Cai et al. 18 Fresh oyster meat was homogenized, pretreated by ultrasound, and then hydrolyzed with alcalase. 19 The crude extract was centrifuged (1800 g, 15 min) and the supernatant was subsequently concentrated. Aliquots of ethanol were added to the hydrolysate with continuous stirring to a final concentration of 30% (v/v) ethanol, and the mixture was kept for 12 h at 4°C. The precipitate was recovered by centrifugation (1800 g, 15 min) and washed with anhydrous ethanol, acetone, and ether in turn. Removal of protein was performed by the Savage method (chloroform/butanol: 5/1), and the samples were then vacuum dried to obtain the C0–30% fraction. Then, the ethanol concentration of the supernatant was further increased to 60% (v/v) and 90% (v/v) in the same manner to obtain the C30–60% and C60–90% fractions, respectively.
Proliferative effects of the polysaccharides on IEC-6 cells
The cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen Co.) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (FBS; Thermo), penicillin-streptomycin (100 IU/mL to 100 μg/mL), 2-mM glutamine, and 10-mM 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid (HEPES) were fed every 2 days and incubated at 37°C in 5% CO2, and harvested by brief treatment with 0.02% ethylenediaminetetraacetic acid and 0.25% trypsin. The cells (1 × 106 cells/well, 100 μL/well) in the culture medium with different final concentrations of oyster polysaccharides were seeded into 96-well plates at 37°C and 5% CO2 for 24 h. Twenty microliters of 3-(4,5-dimethylthiazol-2-yl)-2,4-diphenyltetrazolium bromide (MTT) solution (5 mg/mL in phosphate-buffered saline) was added to each well 4 h before culture termination. Next, the supernatant was discarded and supplemented with 150 μL dimethylsulfoxide. After shaking to dissolve the formazan, the absorbance of each well was determined at 570 nm. Each experimental condition was tested in triplicate.
20
The proliferation rate was calculated using the following equation:
Protective effects of oyster polysaccharides in a model of 5-FU-induced intestinal mucosal damage
Male Sprague-Dawley rats (weighing 300–360 g, Approval No.: SCXK-2008-0002) were purchased from the Laboratory Animal Medical Center (Guangdong, China). Before the experiment, the animals were housed under constant temperature (23°C ± 1°C) and relative humidity (50% ± 5%) and were maintained on a 12-h light–12-h dark cycle (lights on at 07:00 AM) with free access to food and water for 1 week. All procedures regarding animal care and use were supervised and approved by the Institutional Animal Care and Use Committee (IACUC) and Ethics Committee of Sun Yat-Sen University. The permit number for the animal ethics is SCXK-2009-0020.
Eighty Sprague-Dawley rats were randomly divided into four groups and gavaged once a day at 9:00 AM, as described below.
Group 1: Control group, gavaged with normal saline for 13 consecutive days.
Group 2: 5-FU group, gavaged with normal saline for 13 consecutive days and intraperitoneally (i.p.) injected with 5-FU (50 mg/kg·day) from day 5 to 10. 21
Group 3: 5-FU and the oyster polysaccharide-based nutrition formula group (5-FU+OPNF: OPNF contained 1.3 g of C30–60% polysaccharide powder, 1.2 g of isolated soybean protein, 7.0 g of soybean peptide, and 8.0 g of oyster peptide), gavaged with OPNF (2.0 g/kg·day, 20 mL/kg) for 13 consecutive days and i.p. injected with 5-FU (50 mg/kg·day) from day 5 to 10. 21
Group 4: 5-FU and NF group (5-FU+NF: NF was the OPNF formula in which the C30–60% polysaccharide was replaced with equal-quality maltodextrin), gavaged with NF (2.0 g/kg·day, 20 mL/kg) for 13 consecutive days and i.p. injected with 5-FU (50 mg/kg·day) from day 5 to 10. 21
On day 14, rats were deeply anesthetized with ether and then killed by cervical dislocation. The small intestines were removed from the animals through a midline abdominal incision for light and electron microscopy analyses. In addition, serum samples were taken for endotoxin and cytokine (interleukin [IL]-4, IL-10, IL-2, and interferon-γ [IFN-γ]) analyses.
Intestinal mucosal morphology
A 4-cm segment of the jejunum was taken starting at 5 cm after the ligament of Treitz. The intestinal tissue was promptly rinsed with 0.1-M phosphate buffer. The jejunum samples (2 cm) were fixed in 3% glutaraldehyde solution and stored at 4°C overnight. Thin sections were cut, stained with lead citrate and uranyl acetate, and examined with a JEM100CXII (Japan) transmission electron microscope. 22
Separate 2-cm segments were fixed with 10% neutral buffered formalin, embedded in paraffin, sectioned (5 μm), and stained with eosin and hematoxylin. Measurements of villus width and height and mucosal thickness were performed by light microscopy (CX31; Olympus, Japan) using a medical micrograph analyzing system (BI2000; Chengdu Technology & Market Co., Ltd., China).
Determination of plasma endotoxin level
After clotting for 60 min on ice, vena cava blood samples were centrifuged at 2500 g at 4°C for 10 min. Sera were obtained and stored at −70°C. 23 The limulus test (Chinese Horseshoe Crab Reagent Manufactory, Co., Ltd., Xiamen) was used to detect the levels of lipopolysaccharide in serum according to the manufacturer's guidelines.
Enzyme-linked immunosorbent assay
The rats were deeply anesthetized, and the blood was collected from the posterior vena cava before being sacrificed. Serum was isolated from the blood, immediately frozen, and stored at −20°C. The serum concentration of IL-2, IL-4, IL-10, and IFN-γ were analyzed using ELISA kits (Shanghai Ruiqi Biological Technology Co., Ltd., China).
Statistical analysis
The data are presented as the mean ± standard deviation. The results were analyzed using SPSS 16.0 software. The level of significance was determined using hypothesis-testing methods, including one-way analysis of variance (ANOVA). A post hoc Bonferroni test was used to assess the differences between the individual groups. P-values < .05 were considered significant.
Results
Effect of oyster polysaccharides on IEC-6 cell proliferation
C30–60% polysaccharide at 0.0098 and 0.0195 mg/mL concentrations significantly increased IEC-6 cell proliferation in a dose-dependent manner (Fig. 1). At the concentration of 0.0195 mg/mL, C30–60% polysaccharide showed a maximum proliferation rate of 53.61% ± 5.70% (P < .01), but exceeding 0.0195 mg/mL, the proliferation rate decreased. In contrast, compared with the control group, C0–30% and C60–90% significantly increased IEC-6 cell proliferation in a dose-dependent manner (0.0098–0.0391 mg/mL). The variation trend of IEC-6 cell proliferation induced by C60–90% was the same as that observed with C30–60%, and at the higher concentration of 0.0391 mg/mL, IEC-6 cell proliferation was maximized at 50.00% ± 6.60% (P < .01). These results show that the C30–60% polysaccharide fraction produced the strongest proliferation activity in IEC-6 cells. Thus, the C30–60% fraction was added into the enteral NF for further animal experiments.

Proliferative effect of oyster polysaccharides on IEC-6 cells. Compared with the blank control, *P < .05, **P < .01 and ***P < .001. Values are expressed as the mean ± SD. IEC-6, rat small intestinal epithelial cells; SD, standard deviation.
Histological and morphological changes
The jejunum specimens showed atrophy of the villi with fewer goblet cells and a thick and edematous lamina propria. However, this effect was less severe in samples from the 5-FU+OPNF and 5-FU+NF groups than in those from the 5-FU group (Fig. 2). Similar observations were made between the 5-FU+OPNF and 5-FU+NF groups.
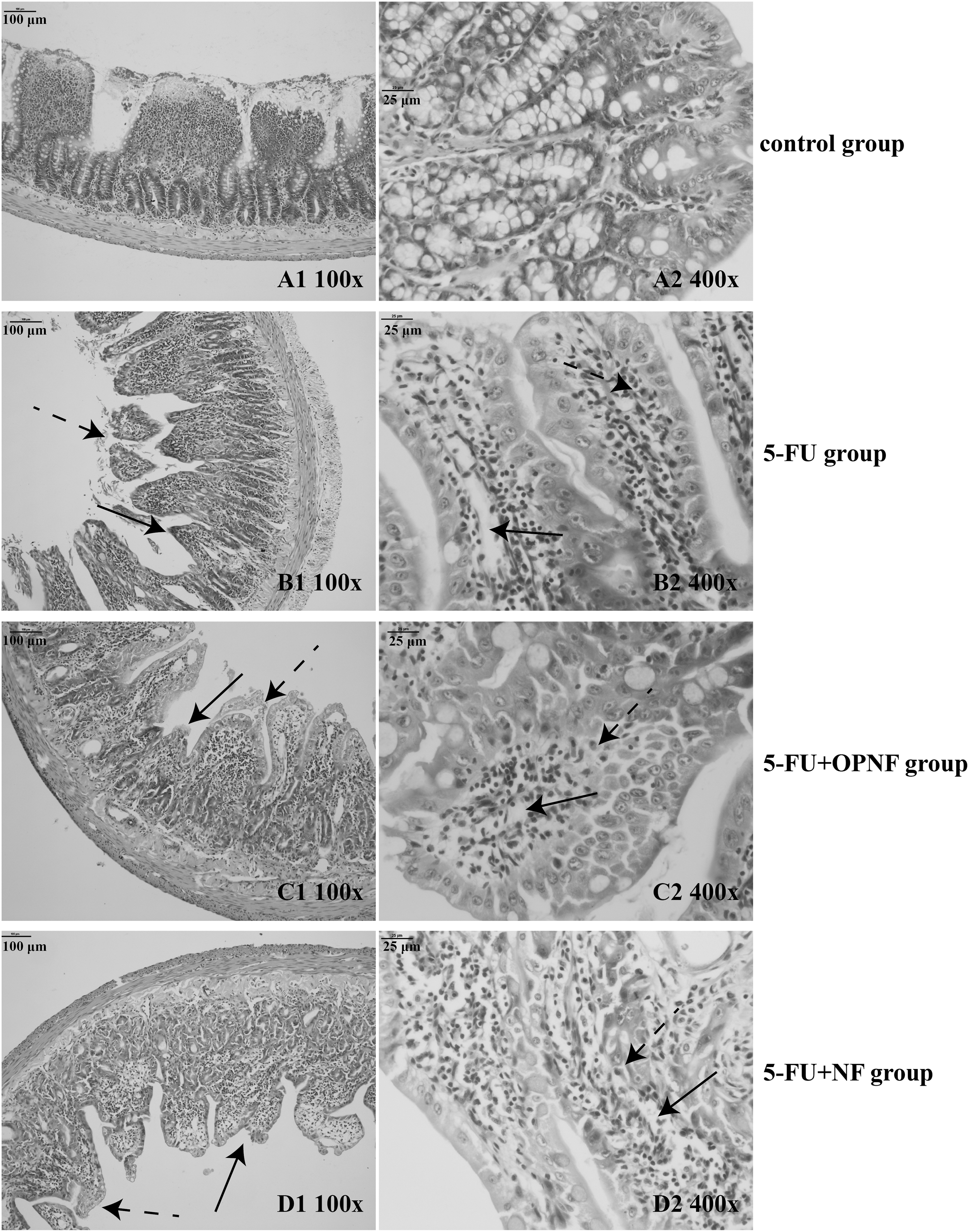
Histological features of the small intestinal mucosa. On day 14, rats were sacrificed, and the jejunum tissues were removed from 10 rats per group. Hematoxylin and eosin staining, original magnifications, 100 × and 400 × .
In the 5-FU group, an intense inflammatory process with ulceration reduced the size of mucosal villi and altered the structure of mucosa in all the studied segments, as shown in Figure 3. Significant reductions in the villus height, villus width, and mucosal thickness were shown in the 5-FU group (P < .001, P < .001, and P < .01, respectively), whereas the rats that received OPNF and NF showed taller villus, thicker mucosa, and an especially greater villus width (P < .01). Moreover, the villus height and width were slightly higher in the 5-FU+OPNF group than in the 5-FU+NF group, but these differences were not significant.
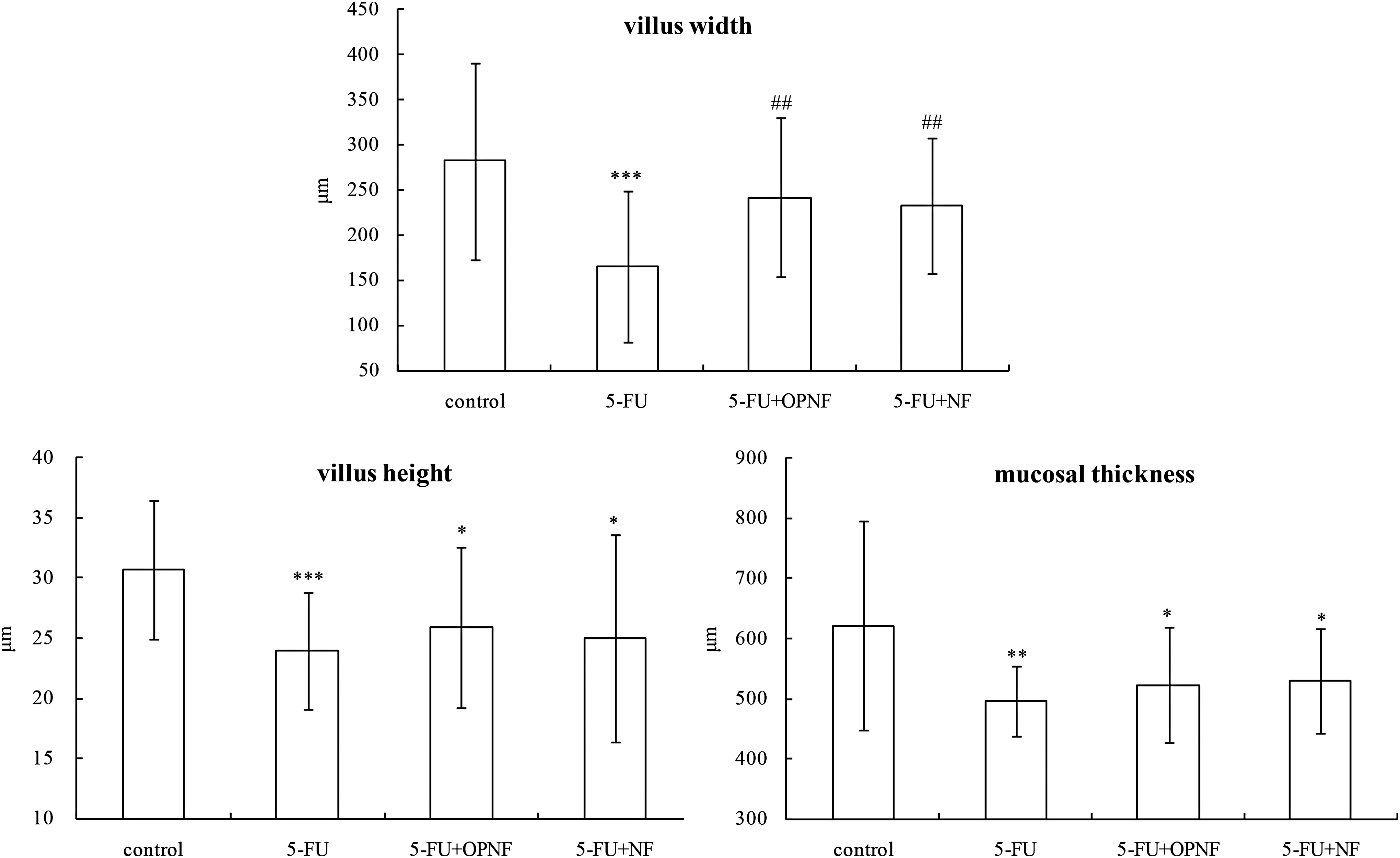
Mucosal morphology. Compared with the 5-FU group, ## P < .01, compared with the control group, *P < .05, **P < .01, and ***P < .001. Values are expressed as the mean ± SD.
Scanning electron microscopy revealed ultrastructural degenerative changes in the 5-FU group, which presented as rarefaction or disappearance of the microvilli, cytoplasmic vacuolations, and swollen mitochondria (Fig. 4). In contrast, these changes reversed in the 5-FU+OPNF group, with mildly exfoliated microvilli and dilated mitochondria, and the effects were also less severe than those in the 5-FU+NF group.

Scanning electron micrograph of the small intestinal mucosa.
In a word, treatment with OPNF and NF partially reduced the injury of intestinal mucosa induced by 5-FU, and the application of OPNF induced a greater reduction in injury.
Plasma endotoxin assays
Previous studies have suggested that 5-FU may induce injury in the intestinal mucosa following endotoxin translocation, which may initiate and exacerbate the inflammatory response and promote multiple organ dysfunction syndrome. 24 As shown in Figure 5, serum endotoxin levels in the 5-FU group were extremely elevated compared to the control group (P < .001). However, the endotoxin levels in the 5-FU+OPNF and 5-FU+NF groups were significantly lower than those in the 5-FU group (P < .001), and there was no significant difference between the normal control, 5-FU+OPNF, and 5-FU+NF groups.

Endotoxin levels in serum. Compared with the 5-FU group, ### P < .001, compared with the control group, ***P < .001. Values are expressed as the mean ± SD.
Changes in cytokine levels in plasma
An imbalance in proinflammatory and anti-inflammatory cytokines might cause intestinal mucosal injury. Proinflammatory cytokines are produced by Th1 cells and maintain inflammation, whereas anti-inflammatory cytokines are produced by Th2 cells and inhibit inflammation. 25 Th1 cytokines, namely, TNF-β (tumor necrosis factor-β), IFN-γ, and IL-2, promote cellular immunity, whereas Th2 cytokines, such as IL-4 and IL-10, are responsible for inducing humoral immunity.
Table 1 showed that, compared with the control group, administration of 5-FU elicited significantly higher increases in proinflammatory cytokine IL-2 (P < .01), but significantly greater decreases in anti-inflammatory cytokine IL-4 (P < .01) and IL-10 (P < .05) levels. In addition, the IFN-γ levels were markedly decreased in the 5-FU (P < .01) group. In contrast, the 5-FU-treated rats fed with OPNF showed significant increase of IFN-γ (P < .05) and IL-10 (P < .05) levels, and decrease of IL-2 levels (P < .001) compared with the 5-FU group. Furthermore, proinflammatory cytokine IL-2 was markedly increased in the 5-FU+NF group compared to that in the 5-FU+OPNF group (P < .001). In conclusion, these results show that OPNF enhanced the immunity and significantly offset the mucosal inflammation that occurred in the 5-FU-treated rats.
Compared with the 5-FU group, # P < .05 and ### P < .001.
Compared with the 5-FU+OPNF group, aaa P < .001.
5-FU, 5-fluorouracil; IFN-γ, interferon-γ; IL, interleukin; NF, nutrition formula; OPNF, oyster polysaccharide-based nutrition formula.
Discussion
The intestinal mucosa barrier, which includes the mechanical, biotic, and immunity barriers, is the first line of defense against a hostile environment. Approximately 40–100% of patients experiencing chemotherapy develop mucositis, 26 which is characterized by the increase of proinflammatory cytokines, increased epithelial cell apoptosis, and altered gastrointestinal microbiota, which causes dysfunction of the intestinal mucosal barrier. Therefore, an immunonutrition-based enteral nutrition should be used to inhibit the development of mucositis and facilitate the recovery process.
Intestinal barrier function depends on the strength of physiological renewal or the repair of pathological intestinal epithelia. Intestinal epithelial injury induced by chemotherapy agents may increase the rapid penetration of toxic substance absorption, immunogenic and inflammatory responses, and imbalance of the host's homeostasis. 20 Our results demonstrated that 5-FU administration in the surface epithelium resulted in separation and less shortening, and significant villi exfoliation and denudation. The cell construction of the villi was disrupted, with obvious widening of the intercellular spaces (Fig. 2). Similar results were previously found in the small intestine of rats treated with 5-FU chemotherapy. 27 Thus, the endotoxin levels were remarkably increased in the 5-FU group (Fig. 5), suggesting a disruption in the epithelial intestinal barrier. Rapid repairing of the epithelial surface barrier under injuries is indispensable to preserve normal homeostasis. In an MTT assay, exposure to oyster C30–60% polysaccharides for 24 h exerted dose-dependent effects on the IEC-6 cell proliferation. Maximal proliferation of IEC-6 cells was observed with the 0.0195 mg/mL concentration of C30–60% polysaccharide fraction. The proliferation of epithelial cells is necessary to supply the decreased cell pool. Moreover, the histological and morphological analyses revealed 5-FU-induced degenerative changes in the histology and ultrastructure of small intestinal mucosa. However, this effect was less severe in the 5-FU+OPNF group. In addition, the height, width, and thickness of the villi were clearly increased in the 5-FU+OPNF group, indicating cell renewal and migration to the villi. Moreover, the villi height and width of the rats fed with 5-FU and OPNF were slightly larger than in that of 5-FU+NF group. The oyster polysaccharide-enhanced NF induced partial mucosal recovery, possibly related to the proliferative effects of oyster polysaccharides on intestinal epithelial cells. However, without oyster polysaccharides, remission of damage to the intestinal mucosa was also observed. This finding may be related to the soybean peptide, which functions in the intestinal tract to enhance nutrient absorption and suppress cholesterol absorption. 28 In addition, soy protein promoted intestinal growth, which was related to effects on intestinal RNA and protein synthesis. 29
Significant endotoxemia in cancer patients is generally observed after chemotherapy, 30,31 and the endotoxin translocation could cause intestinal mucosal barrier dysfunction. In this study, the endotoxin translocation was caused by dramatic mucosal injury induced by 5-FU treatment. Furthermore, the increase in proinflammatory cytokine IL-2 in the plasma coincided with the elevated portal endotoxin levels. However, after administering OPNF and NF, the endotoxin levels in the peripheral blood of the 5-FU-treated rats recovered to control levels. This finding revealed that OPNF and NF ameliorate the intestinal epithelial injury and protect the structural integrity of the intestinal mucosal barrier. With stimulation from endotoxins, the immunocytes, including macrophages, monocytes, and lymphocytes, will secrete inflammatory cytokines. 32 Thus, the timing of the cytokine elevation correlated with the histological changes shown in the intestinal mucosa.
The small intestine immune system is anatomically connected to the systemic immune system by the lymphatic and blood circulation. It has been reported that exopolysaccharides protect the gut mucosal immunity and influence systemic immunity through the cytokines released to the circulating blood. 33 Proinflammatory and anti-inflammatory cytokines are important immunoregulatory factors contributing to chemotherapy-induced gastrointestinal mucositis. 34 In this article the results demonstrate that the cytokine profile shifted from Th1 to Th2 in the peripheral blood of OPNF- and NF-fed rats. The IL-10 level was significantly increased, and the IL-2 level decreased compared to those of 5-FU rats. The anti-inflammatory cytokine IL-10 is secreted by Th2 cells, and they can suppress Th1 cell proliferation, while promoting B cell proliferation, blocking harmful immunological responses and lowering proinflammatory factors from mononuclear macrophages. 35 However, the proinflammatory cytokine IL-2 levels in the 5-FU+OPNF group were significantly lower than those in the 5-FU+NF group. The oyster polysaccharide-enhanced formula induced a more obvious reduction in inflammation in the intestinal mucosa. Furthermore, the NF without oyster polysaccharides also upregulated anti-inflammatory cytokines, possibly because of immunoregulation by oyster peptides. 36 In addition, IFN-γ is secreted by activated natural killer cells and T lymphocytes and plays a major role in adaptive cell-mediated immune responses against viral and bacterial infections. 37,38 It is actively involved in mucosal proinflammatory pathways that are involved in disruption of epithelial apical junction structure, 39 but exerts protective effects against infectious agents to modify the epithelial barrier function. After administering OPNF, the IFN-γ level in the 5-FU-administered rats was obviously elevated in the peripheral blood. Thus, OPNF decreased the expression of tissue inflammatory cytokines and promoted cellular immunity to alleviate 5-FU-induced IM.
In this study, oyster polysaccharides were found to protect against some of the morphological, cellular, and molecular alterations in 5-FU-induced intestinal mucosal damage. Thus, the appropriate administration of C30–60% polysaccharides may decrease the severity of IM by reducing proinflammatory cytokine secretion, releasing anti-inflammatory cytokines, inhibiting inflammatory pathways, improving the barrier function, preventing epithelial cell apoptosis, and eliminating pathogenic bacteria. Building on our previous study of C30–60% polysaccharides, which showed that these polysaccharides can be used as an immunonutrition source to modulate the toxicity and effectiveness of 5-FU chemotherapy, this study provides a theoretical basis for developing oyster polysaccharide-based enteral NFs for cancer patients.
Footnotes
Acknowledgments
The authors thank Prof. Ziniu Yu (South China Sea Institute of Oceanology, Guangzhou, China) for contributing to the identification of C. hongkongensis; the experimental apparatus was provided by the Equipment Public Service Center, South China Sea Institute of Oceanology, Chinese Academy of Sciences; and thanks to Dr. Yongli Gao in the operation of equipment. The study was supported with the Project of Public Science and Technology Research Funds Projects of Ocean (No. 201305018), the National Natural Science Foundation of Guangdong, China (Nos. 2014A030310351 and 2014A030310338), the Innovative Development of Marine Economy Regional Demonstration Projects of Guangdong (Nos. SZHY2012-B01-004 and GD2013-B03-001), and the Science and Technology Planning Project of Guangdong Province, China (Nos. 2013B090800002 and 2015B090904003).
Author Disclosure Statement
No competing financial interests exist.