
Abstract
Panax ginseng (P. ginseng C.A. Meyer, Araliaceae) is used as a therapeutic agent for various diseases. P. ginseng saponins, known as ginsenosides, are the main bioactive compounds responsible for its pharmacological activities. In this work, we have developed a new method of P. ginseng root processing termed solid-state fermentation and examined its effects compared with nonfermented P. ginseng. Mice were fed a high-fat diet (HFD) to induce hyperlipidemia and then received 100 mg·kg bw−1·day−1 of fermented or nonfermented P. ginseng orally for 3 weeks. We assessed the activities of lipogenic pathways and lipid levels in the liver and plasma. The administration of either nonfermented or fermented P. ginseng improved hepatic lipid transfer protein profiles. Nonfermented P. ginseng exhibited significant effects on the regulation of lipid synthesis and oxidation. However, apolipoprotein A4 (apoA4) expression was increased by the administration of fermented P. ginseng. When ginsenosides were analyzed by high-performance liquid chromatography (HPLC), the amounts of the ginsenosides, Rg2, Rc, Rh1(S), Rh1(R), and Rd, were increased by fermentation, with Rd becoming a major constituent of fermented P. ginseng. These findings imply that nonfermented P. ginseng improves hypertriglycemia in HFD-fed mice through regulation of the hepatic lipogenic pathway. In contrast, the effects of fermented P. ginseng were mediated through increased apoA4, leading to decreased triglycerides. The HPLC profiles of ginsenosides suggest that the compositional changes in P. ginseng caused by fermentation processing could be useful in the development of novel triglyceride-lowering therapies.
Introduction
T
Panax ginseng has a long history in folk medicine, and based on ethnopharmacological evidence, the heat-processing method used to strengthen its efficacy is well known in Korea. 7 P. ginseng root can be processed to make white P. ginseng (in which the roots are air-dried after peeling) or red P. ginseng (in which the unpeeled roots are steamed at 98–100°C) to enhance their preservation and efficacy. Heat processing induces changes in the chemical constituents, which can enhance the biological activities of P. ginseng. 8 –10 Accordingly, red P. ginseng is more commonly used as an herbal medicine in Asian countries. In addition, a method of processing white P. ginseng by steaming it at a higher temperature than that applied to the preparation of red P. ginseng produces a novel heat-processed P. ginseng named Sun ginseng, which displays higher free radical scavenging activity than conventional P. ginseng. 5,6 The augmented biological activity of Sun ginseng may be attributed to specific ginsenosides, the main pharmacologically active components of P. ginseng, produced by chemical changes during heat processing, including Rg3, Rk1, and Rg5. 7,11,12 Solid-state fermentation is being increasingly applied to medicinal plants to enhance their therapeutic properties. Yeast, mycelium, and lactic acid bacteria are used as bioconversion microorganisms. 13
P. ginseng has been used as a therapeutic agent for various diseases, including hyperlipidemia. 14 –20 Disturbances in lipid metabolism are at the basis of many chronic disorders. Regulation of lipid metabolism is mainly coordinated by the liver, making it a key target organ for the pharmacological treatment of the abovementioned diseases. Numerous studies have documented the lipid-lowering effects of functional foods or nutrients, and the mechanisms by which they affect lipogenesis or fatty acid oxidation have been elucidated at the transcriptional level. However, mechanistic experiments involving solid-state fermented P. ginseng have not yet been reported.
In this study, we sought to investigate whether the ability of nonfermented P. ginseng to lower triglyceride levels could be augmented by solid-state fermentation. Furthermore, the mechanisms behind the hepatic triglyceride-lowering effects of nonfermented and solid-state fermented P. ginseng were compared in hyperlipidemic mice.
Materials and Methods
Materials
P. ginseng plants grown for 5 years were purchased from the Geumsan Ginseng Agricultural Cooperative Association (Geumsan, Korea) in July 2013. Protease inhibitor mixture solution was purchased from Wako Pure Chemical Industries, Ltd., (Osaka, Japan). Primary antibodies, including sterol regulatory element-binding protein-1 (SREBP-1; sc-13551) and peroxisome proliferator-activated receptor alpha (PPARα; sc-9000), polyclonal antibody against stearoyl-CoA desaturase-1 (SCD-1; sc-14720), apolipoprotein A4 (apoA4; sc-19036), microsomal triglyceride transfer protein (MTP; sc-33116), histone (sc-8030), and β-actin (sc-47778) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Goat anti-rabbit and goat anti-mouse IgG horseradish peroxidase-conjugated secondary antibodies were also purchased from Santa Cruz Biotechnology. Enhanced chemiluminescence (ECL) Western blotting detection reagents were purchased from GE Healthcare (Piscataway, NJ, USA). All other chemicals and reagents were purchased from Sigma-Aldrich.
Processing and extraction of P. ginseng root
Fresh P. ginseng roots were washed under running water, and the rhizome was removed. For solid-state fermentation, the body of P. ginseng was punctured before inoculation for easy penetration of mycelium. Four P. ginseng roots were placed in a 2-L flask and sterilized in an autoclave (HB506-121C; Hanbaek Sci., Gyeonggi, Korea) for 1 h, followed by aseptic inoculation with 20% (w/w) Phellinus linteus liquid spawn. The flask was immediately sealed with a silicone stopper and placed in an incubator at 27 ± 1°C for fermentation. During the first week of solid-state fermentation, inoculum samples were collected and tested for possible contamination. Solid-state fermentation was terminated after 30 days when the surface of P. ginseng was fully covered with the mycelium.
Nonfermented and fermented P. ginseng samples were boiled in 10 volumes of distilled water (w/w) for 150 min using an extractor (DW-290; Daewoong Bio Net., Chungbuk, Korea) designed for medicinal plant extraction. Hot water extraction was repeated twice, and the extracts were combined and filtered using filter paper No. 2 (Whatman, Inc., Clifton, NJ, USA). The extracts were concentrated using a rotary evaporator (Rotavator R-200; Büchi, Flawil, Switzerland) and then freeze-dried using a vacuum freeze dryer (SFDSM06; Samwon Freezing Engineering Co., Busan, Korea).
Analysis of ginsenosides in solid-state fermented P. ginseng and nonfermented P. ginseng
Freeze-dried nonfermented and fermented P. ginseng (10 mg) samples were dissolved in 10 mL of methanol by repeated vortexing and filtered through a Dismic-25 JP membrane filter (pore diameter: 0.5 μm; Advantec Toyo, Tokyo, Japan). High-performance liquid chromatography (HPLC) analysis was performed using an HPLC system (Waters Co., Milford, MA, USA) equipped with a gradient pump, autosampler, and an SPD-20A UV detector. A ZORBAX Eclipse Plus C18 reverse-phase column (4.6 × 250 mm; i.d., 5 μm) was used for separation. The mobile phase consisted of 0.1% formic acid (solvent A) and acetonitrile (solvent B). The gradient elution used was as follows: 0 min, 0% B; 10 min, 0% B; 15 min, 5% B; 54 min, 85% B; and 60 min, 85% B. The column temperature was kept constant at 40°C, and the flow rate was 0.25 mL/min. UV absorbance at 254 nm was monitored using an SPD-20A UV detector. Ginsenosides were assigned by performing coinjection tests with known standard samples and comparing them with UV spectral data. The linearity of the detector response was tested for all ginsenosides (5–100 μg/mL), and the coefficient of correlation was >0.99. The relative standard deviation value of intraday repeatability was lower than 3%, representing good precision.
Experimental animals and treatment
Animal experiments were approved by the Pusan National University Institutional Animal Care and Use Committee (Approval No. PNU-2013-0446). Three-week-old male ICR mice were purchased from DooYeol Biotech (Seoul, Korea). Mice were maintained under a 12-h light/12-h dark cycle and housed at a controlled temperature (23 ± 2°C) and humidity (∼50%). The mice were allowed free access to a laboratory chow diet (powdered CE-2 from CLEA Japan, Inc., Tokyo, Japan, comprising 24.0% protein, 3.5% lipids, and 60.5% carbohydrate) and water. After adaptation, the mice were fed a high-fat diet [HFD, powdered CE-2 containing 40% (w/w) lard] to induce hyperlipidemia. The blood triglyceride level and body weight were measured 4 weeks after the feeding, and to avoid any intergroup differences in these indices, the HFD-fed mice were randomly divided into three experimental groups: group 1 received water, and groups 2 and 3 orally received 100 mg·kg bw−1 ·day−1 of nonfermented or fermented P. ginseng, respectively, for 3 weeks (n = 10 each group). The administration dose and duration were determined based on preliminary experiments. Mice fed the powdered CE-2 diet were used as a normal control group (n = 6). After 3 weeks of treatment, the mice were fasted for 12 h and anesthetized by intraperitoneal administration of a mixture of zolazepam and tiletamine at a dose of 30 mg/kg (Zoletil 50; Virbac, Carros, France) and 10 mg/kg xylazine (Rompun; Bayer Korea, Seoul, Korea). Blood samples were obtained from the inferior vena cava into heparinized tubes. Subsequently, animals were perfused with ice-cold physiological saline and their livers were harvested, snap-frozen in liquid nitrogen, and stored at −80°C until further use.
Measurement of plasma parameters
Plasma triglyceride, nonesterified fatty acid (NEFA), and low-density lipoprotein (LDL) cholesterol concentrations were measured using commercial kits (triglyceride assay kit AM 157S-K [Asan Pharm. Co., Ltd., Seoul, Korea]; NEFA C-Test [Wako Pure Chemical Industries Ltd., Osaka, Japan]; and LDL Cholesterol Assay Kit #5607-01 [XpressBio, Frederick MD, USA], respectively). Hepatic functional parameter (alanine aminotransferase [ALT] and aspartate aminotransferase [AST]) assays were conducted using the Transaminase CII-Test kit (Wako Pure Chemical Industries, Ltd.).
Measurement of liver triglycerides
The livers were homogenized in ice-cold 0.9% NaCl. The homogenate was extracted with a mixture of chloroform and methanol (2:1, v/v) according to the method of Folch et al. 21 Determination of triglyceride content was performed using a commercial kit (Asan Pharm. Co., Ltd.).
Preparation of nuclear and postnuclear fractions
Nuclear protein and postnuclear fractions were prepared as described previously. 22 Briefly, the hepatic tissue was homogenized using lysis buffer, and then 0.1 M dithiothreitol and protease inhibitor mixture were added. After centrifugation (10,500 × g for 20 min at 4°C), the pellet was suspended in the extraction buffer. The mixture was incubated on ice for 30 min, and the nuclear fraction was prepared by centrifugation (20,500 × g for 5 min at 4°C). To extract the postnuclear fraction, hepatic tissue was homogenized in an ice-cold lysis buffer containing protease inhibitor mixture solution and then centrifuged at 2,000 × g for 10 min at 4°C. The protein concentration was determined using a Pierce bicinchoninic acid Protein Assay Kit (Thermo Fisher Scientific, Rockford, IL, USA).
Immunoblot analysis
Ten micrograms of total protein or postnuclear protein underwent sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Separated protein was transferred to a nitrocellulose membrane, blocked with a 5% (w/w) skimmed milk solution, and then incubated with primary antibodies at 4°C overnight. After the blots were washed, they were incubated with secondary antibody for 1.5 h at room temperature. Antigen–antibody complexes were visualized using ECL and detected by chemiluminescence using a Sensi-Q2000 Chemidoc (Lugen Sci Co., Ltd., Seoul, Korea). Band densities were measured using ATTO Densitograph software (ATTO Corporation, Tokyo, Japan), and protein amounts were normalized to histone or β-actin band density. The protein levels are expressed in relation to the expression level in normal mice, which was arbitrarily set to 1.
Statistical analysis
Data are expressed as the mean ± standard error of the mean (SEM). Significance was assessed by one-way analysis of variance, followed by Dunnett's multiple comparison test (SPSS 11.5.1 for Windows, 2002, SPSS, Inc., Chicago, IL, USA). Differences were considered significant if P < .05.
Results
Analysis of ginsenosides
The HPLC profiles are illustrated in Figure 1, and ginsenoside contents are listed in Table 1. Re, Rf, Rc, Rh1(S), Rh1(R), and Rd were the predominant ginsenosides in nonfermented P. ginseng. In fermented P. ginseng, Re and Rf were decreased and Rh1(S), Rh1(R), and Rc were increased. Rd was dramatically increased, and Rg2 was newly formed by fermentation.
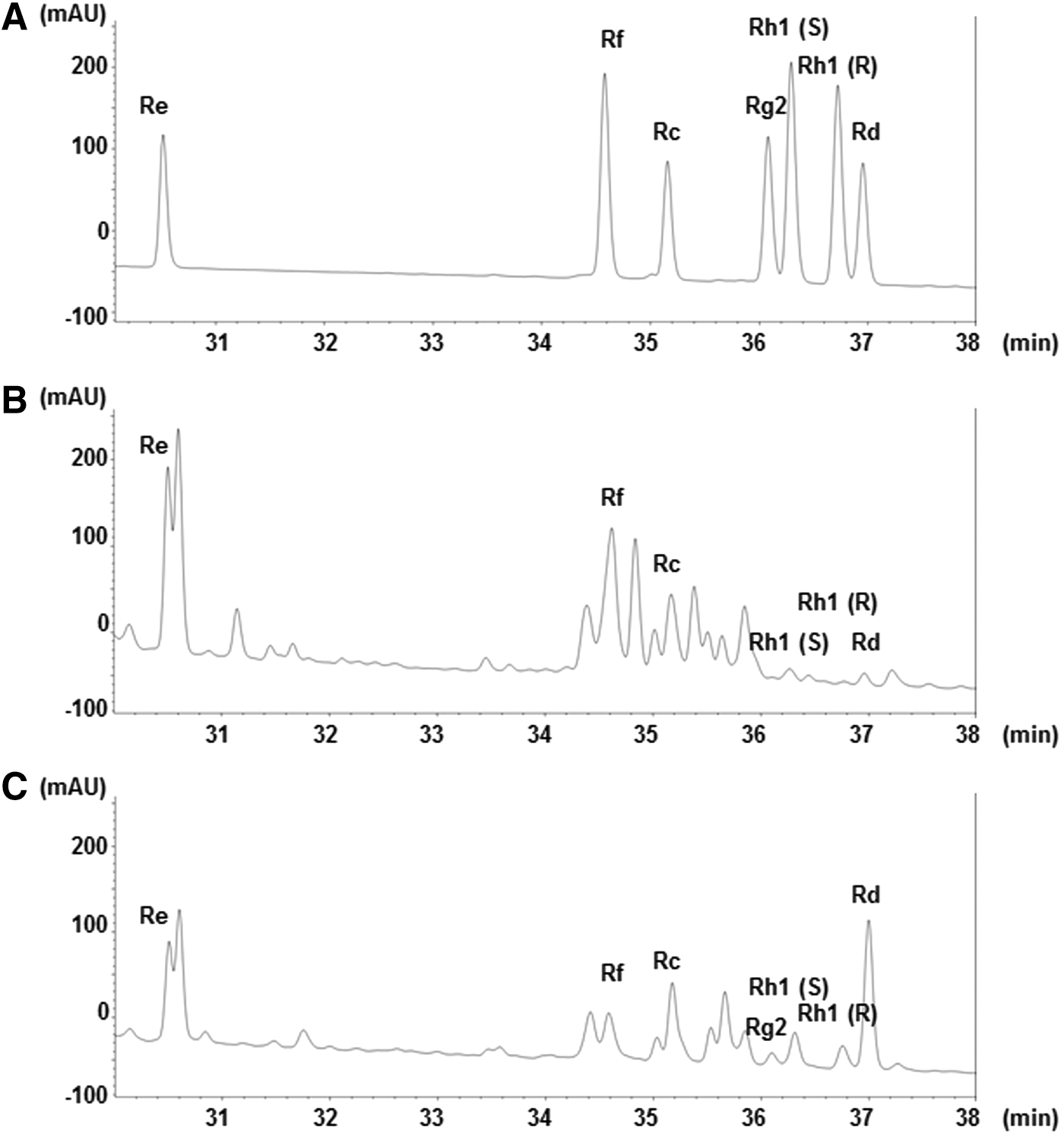
HPLC profiles of standard ginsenosides
Data are expressed as mean ± SEM.
SEM, standard error of the mean; P. ginseng, Panax ginseng.
Food intake and body weight gain
After 3 weeks, the food intake and body weight gain of HFD-fed mice were significantly higher than those of powdered CE-2 diet (normal diet)-fed mice. There were no differences in food intake or body weight gain among the HFD-fed mouse groups (data not shown).
Biochemical parameters of plasma
Plasma triglyceride, NEFA, and LDL cholesterol levels in HFD-fed mice were significantly higher than in normal diet-fed mice (Table 2). The elevated triglyceride level was significantly reduced when HFD-fed mice were administered either nonfermented or fermented P. ginseng. In addition, the elevated LDL cholesterol level of HFD-fed mice was significantly decreased by the administration of fermented P. ginseng, and the NEFA level decreased in the group administered nonfermented P. ginseng. Administration of fermented P. ginseng showed a tendency to decrease plasma ALT and AST levels compared with HFD-fed mice. The nonfermented P. ginseng group also exhibited significantly decreased ALT levels, but that of the AST level was not significant.
Data are expressed as mean ± SEM.
Significance: * P < .01, ** P < .001 versus vehicle-treated HFD-fed mice.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; HFD, high-fat diet; LDL, low-density lipoprotein; NEFA, nonesterified fatty acid.
Hepatic triglyceride content
HFD-fed mice showed a significant increase in hepatic triglyceride content compared with normal diet-fed mice (Fig. 2). This content was significantly decreased in mice administered with nonfermented and fermented P. ginseng.

Hepatic triglyceride content. Data are expressed as mean ± SEM. Significance: *P < .01, **P < .001 versus vehicle-treated HFD-fed mice. FG, fermented ginseng-treated HFD-fed mice; HFD, high-fat diet; N, normal mice; NFG, nonfermented ginseng-treated HFD-fed mice; SEM, standard error of the mean; Veh, vehicle-treated HFD-fed mice.
Hepatic SREBP-1 and PPARα protein expression
The protein expression of hepatic SREBP-1 was significantly higher in HFD-fed mice than in normal diet-fed mice. This increase was significantly reduced by nonfermented P. ginseng administration (Fig. 3A). Hepatic PPARα protein expression was decreased in HFD-fed mice compared with normal diet-fed mice; however, the group treated with nonfermented P. ginseng showed an increase in PPARα protein expression (Fig. 3B). In contrast, hepatic SREBP-1 and PPARα protein expression in HFD-fed mice was unchanged by fermented P. ginseng administration.
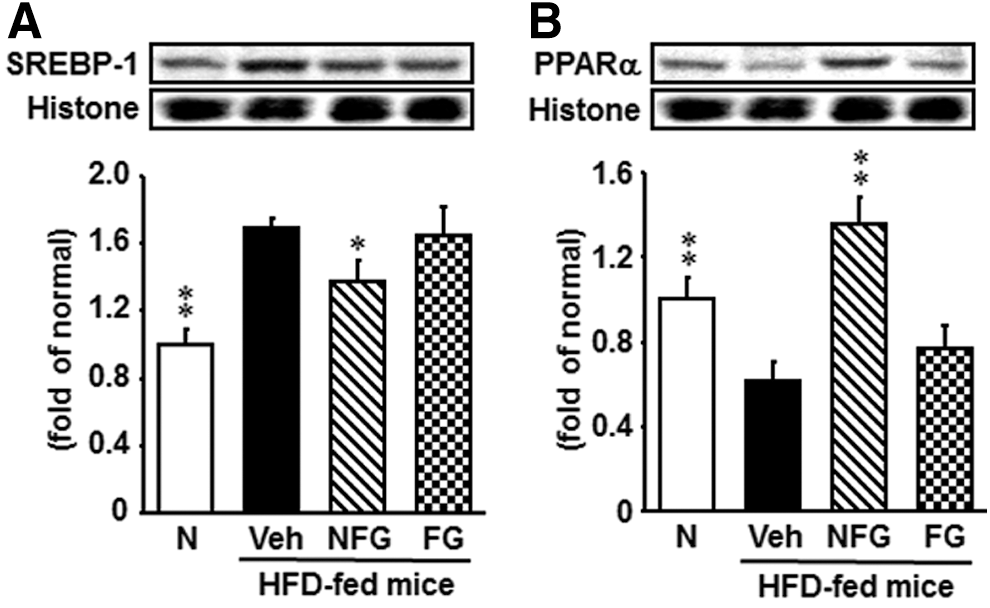
Hepatic SREBP-1
Hepatic SCD-1 protein expression
Overexpression of SCD-1, a protein involved in hepatic triglyceride synthesis, was observed in the hepatic tissue of the HFD-fed group compared with the normal diet-fed group (Fig. 4A). Mice treated with nonfermented P. ginseng had lower expression of SCD-1 than vehicle-treated mice; however, with administration of fermented P. ginseng, there was no change in SCD-1 expression.
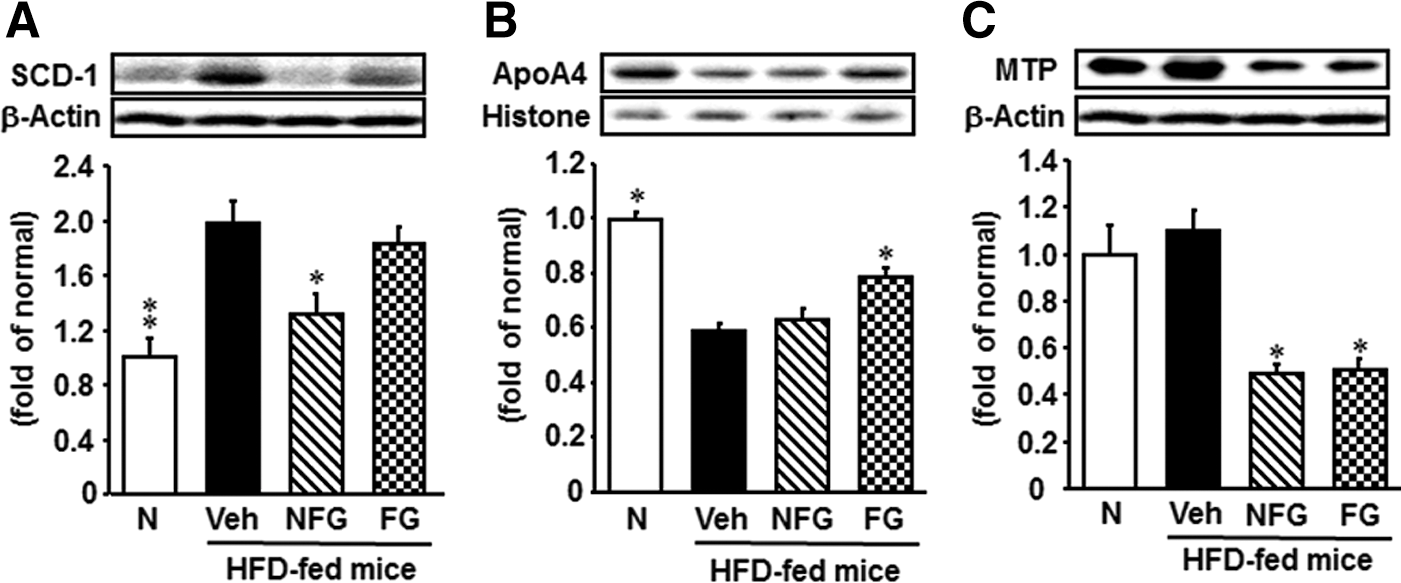
Hepatic SCD-1
Hepatic apoA4 protein expression
The expression level of apoA4 in HFD-fed mice was significantly lower than that of normal diet-fed mice (Fig. 4B). The expression of ApoA4 was significantly increased by fermented P. ginseng administration, but was unchanged after nonfermented P. ginseng administration.
Hepatic MTP protein expression
As shown in Figure 4C, the expression level of MTP was slightly increased (although not significantly) in HFD-fed mice compared with normal diet-fed mice; however, the administration of either nonfermented or fermented P. ginseng to HFD-fed mice decreased the expression of this protein.
Discussion
Regulation of lipid metabolism is mainly coordinated by the liver, which is therefore a key target organ for the pharmacological treatment of these metabolic diseases. Nuclear hormone receptors, particularly the PPARs, are an important class of ligand-activated transcription factors involved in regulation of hepatic lipid metabolism. 23 The PPAR group of nuclear receptors can be separated into three subtypes: PPARα, PPARβ/δ, and PPARγ. 24,25 PPARα is a key regulator of hepatic fatty acid metabolism. Furthermore, lipid homeostasis is regulated by a family of membrane-bound transcription factors called SREBPs. SREBP-1 is a key transcription factor that nutritionally regulates the hepatic gene expression of lipogenic enzymes and triglyceride deposition in the liver. 26 SREBP-1 increases the expression of genes involved in the fatty acid synthetic pathway, such as acetyl-CoA carboxylase alpha, fatty acid synthase, and SCD-1.
In this study, the increase in hepatic SREBP-1 in HFD-fed mice was attenuated by administration of nonfermented P. ginseng. This was probably related to inhibition of hepatic triglyceride accumulation. Furthermore, hepatic PPARα was decreased in HFD-fed mice; it was significantly increased by nonfermented P. ginseng administration. We also found that nonfermented P. ginseng exerted a significant effect on regulation of SCD-1. These results suggest that nonfermented P. ginseng can improve hypertriglyceridemia in HFD-fed mice through regulation of the hepatic lipogenic pathway as well as by downregulation of MTP. Fermented P. ginseng administration did not significantly reduce SREBP-1 or SCD-1 and elevated PPARα, but hepatic triglyceride and MTP levels in fermented P. ginseng-administered mice were significantly lower compared with nontreated mice. These findings imply that the hepatic triglyceride-lowering effects of fermented P. ginseng may be associated with a different mechanism of lipid metabolism.
Apolipoproteins bind lipids to form lipoproteins, allowing lipid transport through the lymphatic and circulatory systems. Apolipoproteins are of considerable physiological importance and are associated with various diseases, including hypertriglyceridemia. They serve as enzyme cofactors, receptor ligands, and lipid transfer carriers that regulate the metabolism of lipoproteins and their uptake in tissues. Among these, apoA4 has been ascribed a wide variety of functions in lipid metabolism and metabolic regulation. ApoA4 influences dietary fat absorption and chylomicron synthesis, 27 modulates the activation of LDL by apoC-II, 28 and activates lecithin–cholesterol acyltransferase. 29 It protects against atherosclerosis by inducing lipid transportation from tissues to the liver for elimination. 30 In our study, apoA4 decreased significantly in HFD-fed mice. However, apoA4 was significantly increased in fermented P. ginseng-administered mice, although nonfermented P. ginseng had no effect. This indicates that the triglyceride-lowering effect of fermented P. ginseng may be attributable to increased expression of apoA4.
According to the HPLC profile, typical ginsenosides of nonfermented fresh P. ginseng are Re, Rf, Rc, Rh1(S), Rh1(R), and Rd. In fermented P. ginseng, the contents of Re and Rf were decreased; Rh1(S), Rh1(R), Rc, and particularly Rd were increased; and Rg2 was newly formed. Recently, Rd and Rh1 demonstrated inhibitory effects on differentiation of the adipocyte-like 3T3-L1 cell line. 31 Rc also effectively inhibited adipogenesis in 3T3-L1 cells, as indicated by a significant reduction in intracellular triglyceride contents and lipid accumulation, without eliciting any apparent cytotoxicity. Furthermore, the suppressive effects of Rc were possibly mediated through decreased expression of major transcriptional activator proteins in the adipogenesis pathway. 32 Based on these reports, we hypothesize that these ginsenosides have potent triglyceride-lowering effects. Our data demonstrate that Rh1-, Rc-, and Rd-rich fermented P. ginseng significantly reduces hepatic triglyceride accumulation by increasing apoA4 expression, leading to decreased MTP. In contrast, nonfermented fresh P. ginseng administration influenced the expression of proteins that function in lipogenesis (Fig. 5). These results contribute a new understanding of how ginsenosides affect lipid metabolism and provide strong evidence for the value of further evaluation of the potential therapeutic roles of ginsenosides in fatty liver disease.

Possible mechanisms of nonfermented and fermented ginseng in the livers of HFD-fed mice.
Footnotes
Author Disclosure Statement
No competing financial interests exist.