
Abstract
Hyacinth bean, Dolichos lablab or Lablab purpureus, has been used for centuries in India and China as an edible pod and animal forage, as well as to treat diarrhea and other gastrointestinal disease in traditional Korean medicine. Recently, we have demonstrated that D. lablab extract (DLL-Ex) prevented free fatty acid-induced lipid accumulation in an in vitro cellular nonalcoholic fatty liver disease (NAFLD) model. In this study, we, thus, aimed at clarifying the hepatoprotective effects of DLL-Ex in a high-fat diet-induced in vivo animal NAFLD model, as well as at elucidating underlying mechanisms of identified effects. Sixty, 6-week-old, male C57BL/6J mice were randomly divided into six groups: a control group fed a low-fat diet, four high-fat diet (HFD) groups, three receiving daily oral supplementation of DLL-Ex (25, 50, and 100 mg/kg/day), and one HFD group receiving daily oral supplementation of MILK (100 mg/kg/day). Effects of DLL-Ex supplementation were evaluated by histopathological and histochemical assessments. DLL-Ex supplementation inhibited HFD-induced increases in body weight and body fat mass and ameliorated increases in body weight, manifested as decreased liver function tests, lower serum triglycerides and cholesterol levels, and increased serum adiponectin levels. The expression of hepatic genes involved in lipid droplet accumulation and in fatty acid uptake was also decreased. We provide evidence of a protective effect of DLL-Ex against HFD-induced fatty liver disease in an animal model.
Introduction
N
Generally, NAFLD results from an increased delivery and uptake of free fatty acid (FFA) into hepatocytes due to a number of underlying mechanisms, which can include: an excess dietary intake or release of FFAs from adipose tissue; increased de novo hepatic FFA and triglyceride (TG) synthesis; failure of very low-density lipoprotein (LDL) synthesis and TG export; and failure of FFA elimination due to impaired hepatic mitochondrial β-oxidation. 8 –10 Therefore, therapies that lower the accumulation of hepatic lipids may be beneficial to control the development and progression of NAFLD. Although it has been suggested that a reduction in overall body weight might reduce the risk of NAFLD, there is currently no evidence to support the effectiveness of pharmacological therapy for the treatment of NAFLD. 11 Current treatments are primarily directed toward improving metabolic parameters that contribute to the pathogenesis of NAFLD. 12
Traditional medicines and herbal products, in particular, have been widely used for generations in the treatment of liver diseases and may, therefore, provide novel pharmacological candidates for an efficient and safe treatment of NAFLD. In recent years, as traditional medicine is quickly becoming a popular alternative to western medicine, there has been increasing interest in identifying alternative approaches in medicine in general, and in the treatment of NAFLD specifically. 13,14 Multiple components of herbal extracts are considered to have diverse functions due to synergistic actions among the bioactive components, and they are also beneficial in preventing adverse effects. Our interest in this study was to evaluate the hepatoprotective effects of a novel Dolichos lablab extract (DLL-Ex). D. lablab has been reported to have antibacterial, antifungal, and hemagglutinating activity, as well as bioefficacy against causal agents of malaria, dracunculiasis, and amoebiasis, and antioxidant activities. 15 –17 D. lablab has also been used as an antipyretic and antispasmodic agent, as well as to treat stomach and phlegmatic disorders. Recently, we have demonstrated that DLL-Ex prevented FFA-induced lipid accumulation in an in vitro NAFLD model. 18 In the previous study, the results indicated that DLL-Ex inhibits cellular lipid accumulation, downregulates mRNA expression of fatty acid synthesis-associated genes, and reduces hepatic FFA uptake by inhibiting fatty acid translocase (FAT, CD36) expression. 18 Therefore, the aim of our study was to clarify the hepatoprotective effects of DLL-Ex in a high-fat diet-induced in vivo animal NAFLD model, as well as to elucidate underlying mechanisms of identified effects.
Materials and Methods
Preparation of DLL-Ex and milk thistle extracts
Dried D. lablab was purchased from a commercial supplier (Kwangmyung-Dang, Ulsan, Korea). All voucher specimens were deposited in the herbal bank of the Korea Institute of Oriental Medicine (KIOM112819). Dried D. lablab (total 3 kg) was pulverized and extracted with 10 L of distilled water for 3–4 h at 90–100°C. The crude extract solution was filtered, evaporated, and lyophilized in a freeze dryer, to yield 501.1 g (dry weight) of DLL-Ex (16.7% w/w yield). The DLL-Ex was suspended in sterilized distilled water at the appropriate concentrations. Since no reliable hepatoprotective drugs have been developed thus far, a large number of herbal extractions have become increasingly popular for the hepatoprotection. Among them, hepatoprotective effects of milk thistle extracts against a variety of toxins were reported and sold and taken as a herbal supplement. Therefore, we used milk thistle extracts as a positive control to compare experimental results. Lyophilized powder of milk thistle extracts was commercially obtained (Narae Trade, Incheon, Korea).
Animal and treatment
Male C57BL/6J mice were purchased from Samtako Bio, Inc. (Seoul, Korea). All mice were kept under controlled temperature (22°C ± 1°C) and humidity (55% ± 10%), on a 12-h light/dark cycle. For NAFLD studies, mice were randomly divided into 6 groups of 12 animals, with equal distribution of body weight: control group; high-fat diet (HFD) group; HFD plus 25 mg/kg/day DLL-Ex group; HFD plus 50 mg/kg/day DLL-Ex group; HFD plus 100 mg/kg/day DLL-Ex group; and HFD plus 100 mg/kg/day MILK group. Mice in the control group were fed a Purina diet with 10% of the calories derived from fat (Koatech, Seoul, Korea). Mice in the HFD groups were fed a pellet rodent diet, with 60% of the total calories derived from fat (Center Lab. Animal, Inc., Seoul, Korea). Animals were sacrificed for analysis after 9 weeks. Mice were fasted overnight before being sacrificed, with blood samples collected via cardiac puncture of anesthetized mice. The serum was separated from blood cells via centrifugation and was stored at −80°C until analysis. Livers and other tissues were excised, rinsed with phosphate-buffered saline to remove blood from the tissue, and snap-frozen in liquid nitrogen. Tissue samples were also stored at −80°C until analysis. All animal procedures were performed in accordance with the guidelines for the Care and Use of Laboratory Animals developed by the Institute of Laboratory Animal Resources of the National Research Council, and they were approved by the Institutional Animal Care and Use Committee of Daejeon University in Daejeon, Korea.
Histopathological examinations and Oil Red O staining
For histopathological examination, the abdomen was opened and the liver was quickly removed to obtain fresh liver tissue specimens that were fixed in 10% formalin and prepared into paraffin blocks. Sections from the blocks were stained with hematoxylin and eosin (H&E). To determine hepatic lipid droplet accumulation, frozen sections of liver tissue were stained with Oil Red O (Sigma, St. Louis, MO). The sections were visualized under an Olympus CKX41 microscope (Olympus, Tokyo, Japan), and digital images were captured at a magnification of 200× by using the Motic image Plus 2.0 program (Motic, Causeway Bay, Hong Kong). Adipose tissues were extracted by separating abdominal subcutaneous fat, epididymal adipose tissue, kidney adipose tissue, and visceral fat, with the weight of each tissue specimen weighed.
TG accumulation assay
The liver TG content was enzymatically measured by using a commercial kit (Asan Pharm. Co., Seoul, Korea), according to the manufacturer's instructions. The TG content of liver tissues (100 mg) was measured and expressed as mg/g liver.
Serum chemistry analysis
Samples of cardiac whole blood collected from anesthetized animals, after an overnight fast, were centrifuged at 2000 g for 15 min at 4°C to separate blood serum from blood cells. The serum biochemical concentrations of TGs, total cholesterol (TC), LDL cholesterol, high-density lipoproteins (HDL) cholesterol, glucose, creatinine, FFA, alanine aminotransferase (ALT), and aspartate aminotransferase (AST) were determined by using an automatic analyzer (Hitachi-720; Hitachi Medical, Japan) with reagents (Bio-Clinical System, Gyeonggi-do, Korea). Serum adiponectin and insulin-like growth factor 1 (IGF-1) levels were measured by immunoassay (ELISA) using commercially available kits (Linco Research, Inc., St. Charles, MO, USA). Blood serum was placed in wells of a microplate coated with diluted antibody and incubated overnight at 4°C overnight. Each well was washed three times with buffer, to which 100 μL of plasma (10-fold dilution) was distributed and incubated for 1 h at room temperature, followed by two subsequent washes with buffer. Then, 100 μL of streptavidin-horseradish peroxide conjugate was added and incubated for 1 h at room temperature, followed by a wash. A total 100 μL of tetramethylbenzidine substrate was added and then incubated in darkness for 30 min, followed by the addition of 50 μL of a stop solution. Absorbance at a wavelength of 450 nm was measured by using an ELISA reader.
RNA isolation and quantitative real-time polymerase chain reaction
RNA was isolated from 30 mg samples of liver tissue by using Trizol (Invitrogen), according to the manufacturer's instruction. Reverse-transcription and quantitative-polymerase chain reaction (qPCR) were performed to detect relative mRNA expression by using Taqman probes (ABI), as previously described. 18
Protein extraction and western blotting
The lysates of liver tissue prepared from 100 mg of the liver tissue of mice were centrifuged at 12,000 g for 10 min, and aliquots of the supernatant containing 20 μg of proteins were subjected to 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). After electrophoresis, the proteins were blotted onto polyvinylidene fluoride membranes. The membranes were incubated overnight at 4°C with specific antibodies for the genes of interest and β-actin. The blots were washed three times for 10 min each with TBS-T, after which the membranes were incubated for 2 h with anti-secondary antibodies. The immunoblots were visualized by using an ECL chemiluminescence detection system (Bio-Rad, Hercules, CA, USA), and the bands were visualized by using a chemiluminescence imaging system (ImageQuant LAS 4000 mini; GE Healthcare, UK). Primary antibody Adrp/Plin2C, Fsa27/Cidec, very-low-density lipoproteins (VLDLR), CD36, and Dgat1 were purchased from Abcam (Cambridge, UK) and Dgat2, UCP2, and β-actin were purchased from Cell Signaling Technology (Danvers, MA, USA). Secondary antibody, anti-rabbit, anti-mouse, and anti-goat were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Ultra-performance liquid chromatography quadrupole time-of-flight mass spectrometry analysis
To investigate the multiple component profiling of the DLL-Ex, a UPLC was performed by using an ACQUITY UPLCTM system (Waters Corporation, Milford, MA, USA) with a photo diode array detector and a sample manger coupled to a Micromass QTOF PremierTM mass spectrometer (Waters Corporation) equipped with an electrospray interface. Chromatographic separations were performed on a 2.1 × 100 mm, 1.7 μm ACQUITY BEH C18 (Waters Corporation) chromatography column. The column temperature was maintained at 35°C, and the mobile phases A and B were water with 0.1% formic acid and acetonitrile with 0.1% formic acid, respectively. The gradient conditions were as follows: 0 min, 10% B; 1–3 min, 10–15% B; 3–4.3 min, 15–30% B; 4.3–10 min, 30–70% B; and 10–12 min, 70–100% B; they were then held for 1.4 min before returning to the initial conditions. The flow rate was 0.4 mL/min, and the loading volume was 2.0 μL. The mass spectrometer was operated in negative ion mode. N2 was used as the desolvation gas. The desolvation temperature was set to 350°C at 400 L/h with a source temperature of 110°C. The capillary and cone voltages were set to 2350 and 50 V, respectively. The data were collected for each sample with a 0.25 sec scan time and a 0.01 sec interscan delay.
Statistical analysis
All experimental data are shown as mean values ± standard error of the mean. Between-group differences were evaluated by using a one-way analysis of variance, followed by Dunnett's test to identify significant differences between the HFD and CON groups. Significant P-values are reported as follows:* and #, P < .05; ** and ##, P < .001.
Results
Effect of DLL-Ex on fatty liver and body weight in HFD-induced NAFLD mouse model
To identify possible dose-related effects on the protective mechanisms of DLL-Ex against HFD-induced fatty liver disease in vivo, we administered three different DLL-Ex doses to different groups of HFD mice (25, 50, and 100 mg/kg/day), and one HFD group was administered milk thistle extract (MILK, 100 mg/kg/day) as a positive control for the 9-week period. Consistent with previous reports, a 9-week HFD diet resulted in an accumulation of fat in the animals' liver and the development of liver steatosis (Fig. 1A). On visual examination, the livers of mice in the HFD group were pale yellow in appearance and markedly enlarged, compared with those of mice in the control group. Liver histopathology revealed widespread lipid vacuole deposits in the HFD group, compared with the control group. DLL-Ex treatment reduced this microvesicular fat accumulation. Oil Red O-stained sections confirmed a significantly higher accumulation of lipid droplets in hepatocytes of mice in the HFD group, but the accumulation was obviously decreased in DLL-Ex-treated (25, 50, and 100 mg/kg/day) and MILK-treated (100 mg/kg/day) groups.

Effects of DLL-Ex on mice liver histology and body weight in an HFD-induced mice model. Histopathological examination by H&E and Oil Red O staining
Since an HFD induced not only fatty liver but also obesity, we evaluated the chronic effects of DLL-Ex on body weight. As shown in Figure 1B, the HFD-induced body weight gain was significantly reduced by a daily oral administration of DLL-Ex (25, 50, and 100 mg/kg/day) or MILK (100 mg/kg/day) supplement, indicating that DLL-Ex treatment inhibited the obesity caused by an HFD. However, daily food and water consumption were not significantly affected by DLL-Ex treatment.
The weights of the livers of the mice in the HFD group were 1.4-fold greater than the liver weight in mice in the control group. DLL-Ex treatment attenuated the effects of an HFD diet on liver weight, with restoration of the weight to normal control levels (Fig. 1C). Therefore, DLL-Ex treatment significantly prevented HFD-induced fatty liver and hepatomegaly.
We further confirmed that HFD induced hepatic lipid accumulation by quantitative analysis of hepatic TG content. Along with the hepatic steatosis phenotype, the hepatic TG levels in the HFD group of mice were substantially higher than those in the control group. Significantly lower hepatic TG levels were observed in mice receiving 50 and 100 mg/kg/day of DLL-Ex or MILK (Fig. 1D).
Total body fat, abdominal subcutaneous fat, epididymal fat, and intestinal fat were higher in the HFD groups compared with the control group. Total body fat, abdominal subcutaneous fat, and epididymal fat of mice treated with DLL-Ex (100 mg/kg/day) were significantly lower compared with the HFD groups (Fig. 2).
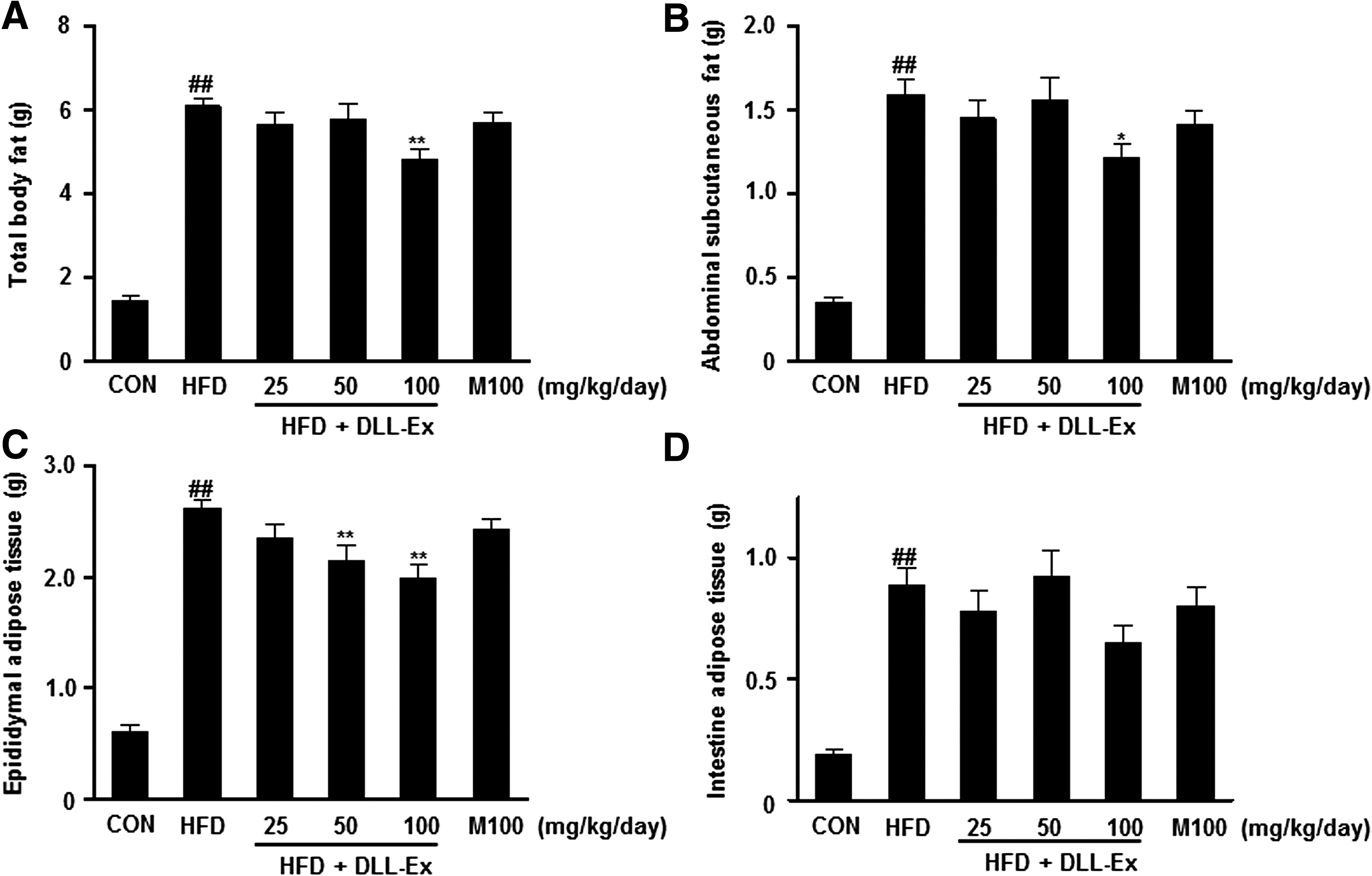
Effects of DLL-Ex on body weight gain and body fat mass in HFD mice showing:
Effect of DLL-Ex on serum liver marker enzymes
To address the effect of DLL-Ex on liver function and damage, serum enzyme activity of ALT and AST was evaluated. The serum ALT and AST levels were significantly higher in mice in the HFD groups, compared with those in the control group (Fig. 3). The HFD-induced increase in serum ALT and AST levels was dramatically suppressed by DLL-Ex treatment. In fact, a 100 mg/kg/day dose of DLL-Ex resulted in levels of ALT and AST that were near normal control levels (Fig. 3).
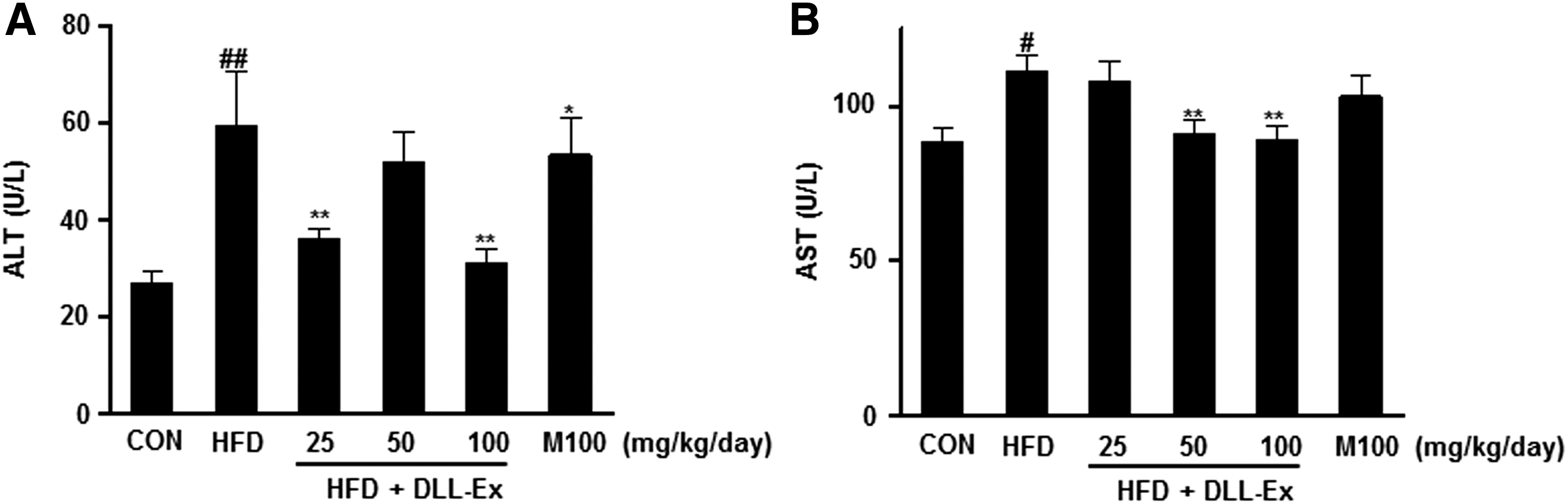
Effects of DLL-Ex on serum ALT and AST levels in HFD mice.
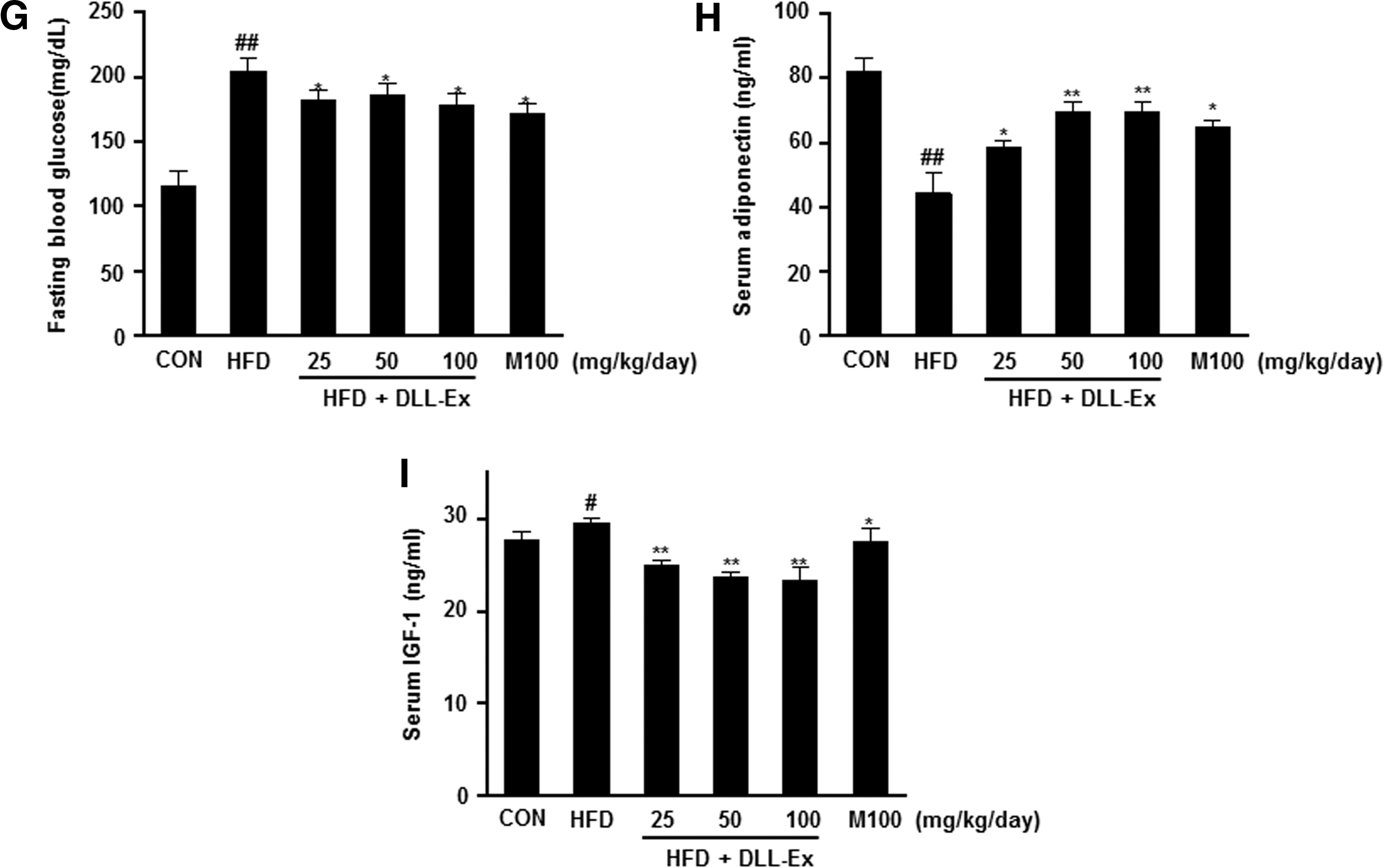
The effects of DLL-Ex on other serum parameters of TG, TC, LDL, HDL, FFA, creatinine, and blood glucose were also investigated. As shown in Figure 4A–G, the increased levels of TG, TC, LDL, FFA, and blood glucose were all significantly attenuated by DLL-Ex treatment in HFD mice. Further, suppressed serum adiponectin levels of HFD mice were dose dependently elevated by DLL-Ex (Fig. 4H). However, the HFD-related elevation of serum levels of IGF-1 was suppressed by DLL-Ex in HFD mice (Fig. 4I). Although serum HDL-cholesterol and creatinine levels were increased by an HFD, there were no significant differences in concentrations among all DLL-Ex (25, 50, and 100 mg/kg/day) groups and the MILK (100 mg/kg/day) group, compared with the HFD group (Fig. 4D, F).
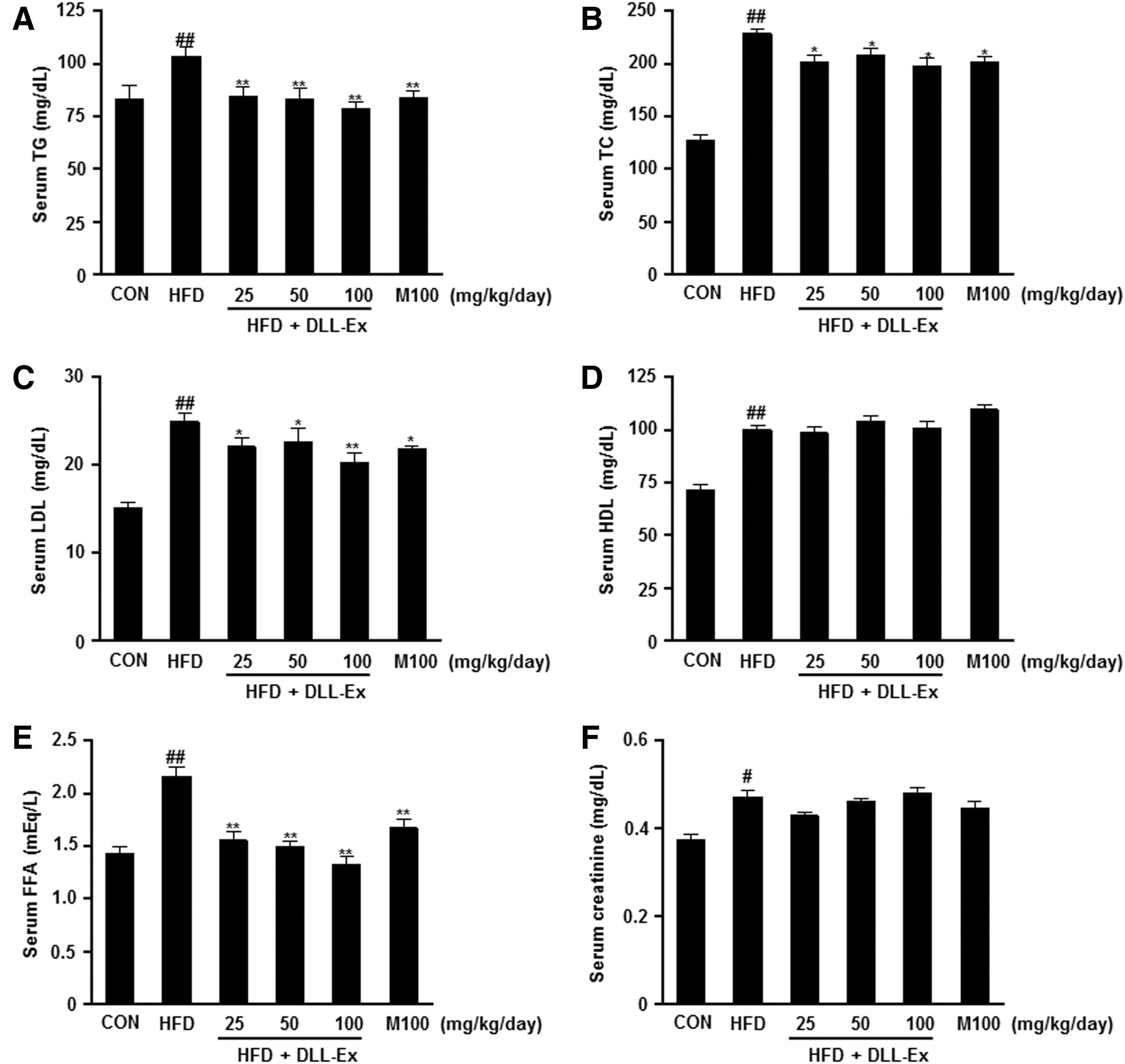
Effects of DLL-Ex on blood biochemistry parameters in HFD mice.
Effects of DLL-Ex on gene expression involved in lipid droplet accumulation and fatty acid uptake in liver
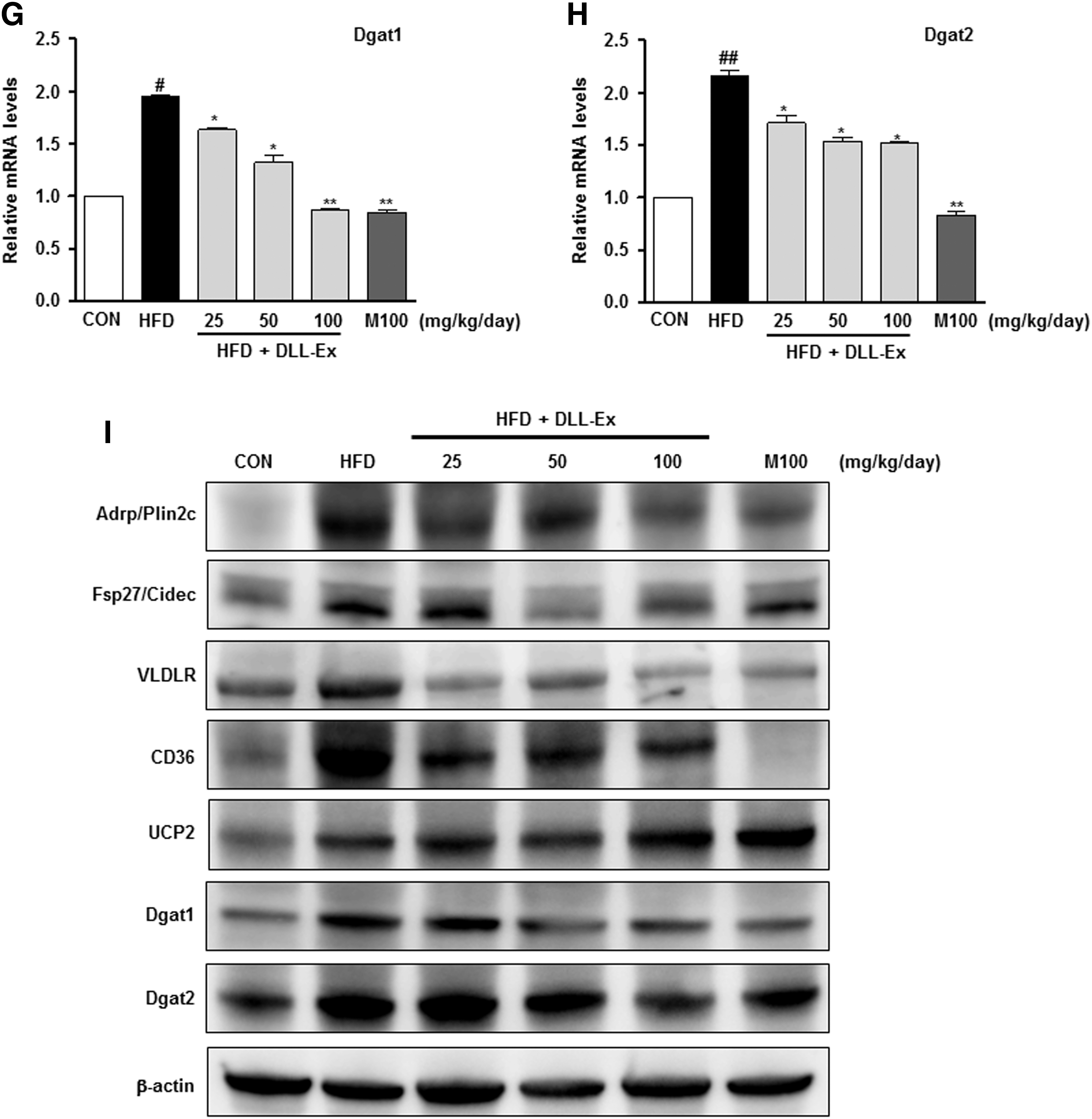
To further quantify the protective effects of DLL-Ex on an HFD-induced hepatic steatosis, we performed a lipid-associated pathway-specific quantitative real-time PCR analysis, using liver tissues from mice receiving an HFD, with or without DLL-Ex supplementation. Fat-inducing transcript 2 (Fit2), also known as fat storage-inducing transmembrane protein 2 (Fitm2), is involved in the microvesicular accumulation of lipid droplets. Levels of Fit2 were significantly higher in the HFD group, with a notable comparative decrease in the liver of mice treated with DLL-Ex (Fig. 5A). The expression of the genes for adipose differentiation-related protein (ADRP), also known as perilipin 2 (Plin2) and fat-specific protein 27 (Fsp27, also known as Cidec), involved in lipid droplet accumulation, was higher in the HFD group and lower in the HDF mice treated with DLL-Ex (Fig. 5B, C). HFD increased not only the expression of VLDLR but also the expression of CD36 genes involved in lipid delivery. DLL-Ex (25, 50, and 100 mg/kg/day) and MILK- (100 mg/kg/day) supplementation significantly inhibited HFD-elevated VLDLR and CD36 expression (Fig. 5D, E). Although the mRNA expression of mitochondrial uncoupling protein (UCP2) was not significantly affected by an HFD, supplementation with DLL-Ex (25, 50, and 100 mg/kg/day) or MILK (100 mg/kg/day) significantly elevated mRNA levels of UCP2 in the liver (Fig. 5F). Higher mRNA levels of diacylglycerol acyltransferase-1 and 2 (DGAT1 and DGAT2, respectively), which catalyze TG synthesis, were observed in the HFD group. In contrast, DLL-Ex or MILK treatment significantly suppressed DGAT1 and DGAT2 mRNA levels (Fig. 5G, H). Through this analysis, we identified that expression of genes involved in hepatic lipid import and lipid droplet accumulation was significantly ameliorated in the DLL-Ex- and MILK-treated group, compared with the HFD group (Fig. 5). Western blot analysis also confirmed that proteins (Adrp/Plin2c, Fsp27/Cidec, VLDLR, CD36, Dgat1, and Dgat2) were involved in lipid droplet accumulation and in lipid delivery, and expression was higher in the HFD group and lower in the HDF mice treated with DLL-Ex (Fig. 5I). Interestingly, HFD increased UCP2 protein levels and DLL-Ex and MILK treatment increased UCP2 protein more in the liver.
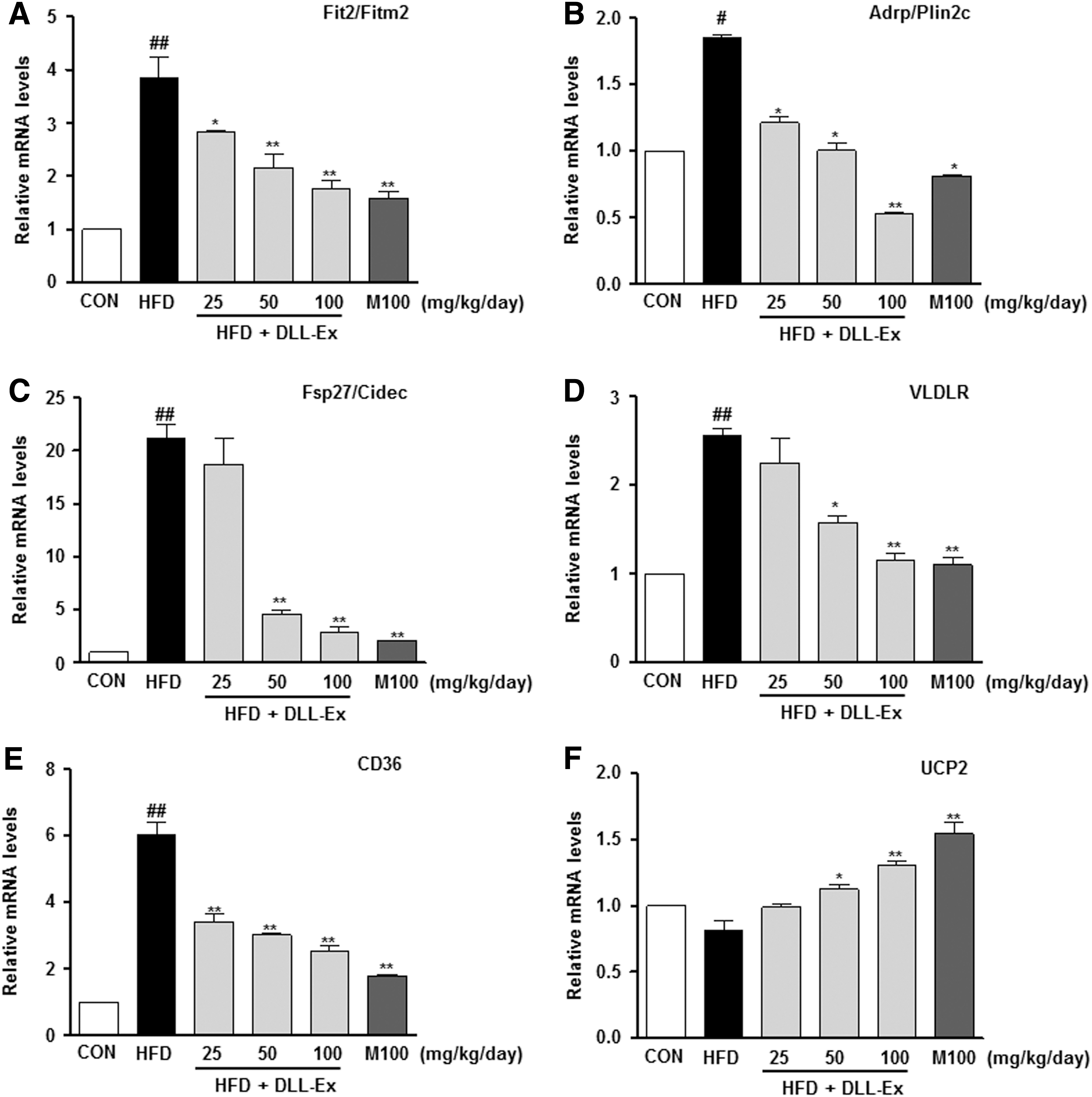
Real-time PCR analysis of liver mRNA and western blot analysis of liver protein isolated from mice treated with DLL-Ex in the HFD group. The expression of hepatic genes involved in lipid droplet accumulation was addressed by real-time quantitative PCR
Discussion
In this study, we report that DLL-Ex, an extract of medicinal bean, suppressed hepatic lipid accumulation and downregulated mRNA levels of genes that mediate fatty acid uptake and lipid accumulation in HFD mice. Our results provide an additional understanding of the pharmacological function of a novel decoction, and of the potential usefulness of DLL-Ex as a valuable therapeutic agent against NAFLD.
NAFLD encompasses a broad spectrum of liver diseases that can progress from simple fatty liver to NASH and, ultimately, to cirrhosis. 19 Insulin resistance, oxidative stress, cytokines, intestinal permeability, and obesity have been identified as the major factors involved in NAFLD pathogenesis. 4 NAFLD can develop as a primary outcome of metabolic stress or as a secondary outcome of primary hepatitis, and it promotes the progression of an inflammatory disease of the liver to fibrosis. 20
Accumulation of TG droplets within individual hepatocytes is a common feature of all stages of NAFLD, including steatosis, inflammation, necrosis, and, ultimately, fibrosis and cirrhosis. 10 High circulating levels of FFA can trigger a series of biological changes in hepatic lipid metabolism. Increased transport of FFAs into hepatocytes would increase levels of hepatic de novo lipogenesis, which would exceed the export capacity of hepatic FFA for β-oxidation and very LDL, leading to increased hepatic steatosis. 21 Current treatment of NAFLD focuses mainly on weight loss and management of cardiometabolic risk factors via the correction of elevated cholesterol, TGs, and blood sugar level. However, no truly effective therapy has been found to date. In our study, we clearly demonstrated a protective effect of DLL-Ex on FFA/HFD-induced fatty liver in an in vivo mouse model. We further identified DLL-Ex-specific mechanisms of inhibition of lipid uptake into the liver and hepatic lipid droplet accumulation.
Although traditional oriental herbal medicines have long been used to treat various conditions in humans, including liver disease, it is important that the cytotoxicity of herbal medicine should be considered before use. In this study, we used an in vitro cellular model of hepatic steatosis to investigate the potential hepatoprotective effect of DLL-Ex against FFA-overloaded steatotic cells.
The excess accumulation of lipid in the liver produces hepatomegaly and fatty liver disease. 22 Hepatic TG levels and liver weight were increased by an HFD. These effects were attenuated by DLL-Ex treatment. Therefore, DLL-Ex protects against long-term HFD-induced liver damage. Reducing body weight and fat mass is important for preventing fatty liver disease. Although weight gain increased in all mice fed an HFD, mice treated with DLL-Ex gained significantly less weight from week 3 to 9, compared with mice in the HFD control group. Our finding of a reduced body fat weight in HFD mice treated with DLL-Ex confirmed a direct effect of DLL-Ex on fat mass, rather than on muscle and bone mass. When we further consider the upregulation of UCP2 expression with DLL-Ex supplementation (Fig. 5F), we propose that DLL-Ex treatment induced hypercatabolism, which allowed the balance of energy storage to be restored and/or reduced the rate of carbohydrate and lipid absorption in the gastrointestinal tract. Although mice fed an HFD show increased UCP2 mRNA expression in the liver, 23 we could not detect any significant increase in UCP2 mRNA levels in HFD mice. However, DLL-EX and MILK treatment increased not only the UCP2 mRNA levels but also protein levels in the liver. These results suggested that the increased hepatic expression levels of UCP2 in DLL-Ex- or MILK-fed mice, possibly via PPARα, may enhance energy expenditure for the prevention of obesity. Moreover, it has been reported that silibinin, a natural plant flavonolignan, is the main active constituent found in milk thistle and is able to significantly increase the expression levels of UCP1, 2, and 3. 24
Elevation of serum levels of ALT is a surrogate biomarker for NAFLD and steatohepatitis in clinical practice, being used for diagnosis and to assess damage to the structural integrity of the liver. 25,26 We also observed high plasma levels of ALT in the HFD group, but significantly less in the DLL-Ex-treated group. Therefore, it is likely that DLL-Ex restored hepatocyte levels and normalized the integrity of the plasma membrane of hepatocytes, preventing leakage of the ALT enzyme into the blood.
Insulin resistance increases TG synthesis, fatty acid uptake, and peripheral lipolysis, resulting in fat accumulation in the liver. 27 TGs combine with special proteins to form tiny spheres, called lipoproteins, which are sent into the bloodstream to circulate among the cells of the body. 26 When the release or secretion of lipoproteins from the liver is interrupted or the flow of TGs to the liver is increased, fat accumulates in the cells of the liver. 28
Serum HDL and creatinine levels were increased in HFD mice compared with control animals. However, there are no apparent changes in the serum HDL and creatinine after DLL-Ex intervention. Most of the reduced serum cholesterol was LDL, a reduction in LDL would be advantageous clinically, and, in fact, it was clearly shown that the DLL-Ex improved the hypercholesterolemia induced by an HFD. In addition, there was no significant increase in the serum creatinine of DLL-Ex or MILK, indicating that the DLL-Ex or MILK was not nephrotoxic since the creatinine is a marker of kidney function.
Adiponectin is a hormone that enhances insulin sensitivity in the liver and increases FFA oxidation in several tissues. 29 Hepatic lipid homeostasis is regulated by multiple lipid-associated pathways, which include fatty acid import, lipogenesis, lipolysis, fatty acid oxidation, and lipid deposition. Development of hepatic steatosis is related to a dysregulation of one or more of these lipid-associated pathways. To further gain insights into the preventive effect of DLL-Ex against hepatic steatosis induced by HFD, we performed a lipid-associated pathway-specific quantitative real-time PCR analysis, using liver tissues from mice treated with or without DLL-Ex, together with HFD. Through this analysis, we identified that the expression of genes involved in hepatic lipid import, lipid droplet accumulation, and pro-inflammatory cytokines was significantly ameliorated in the DLL-Ex-treated group, compared with that in the HFD group (Fig. 5).
As previously reported, we observed markedly increased expression of the lipid droplet accumulation gene (Fsp27/Cidec) in mice receiving an HFD. 30 Fsp27/Cidec is an important regulator of energy homeostasis, with its physiological functions related to the development of metabolic disorders, including obesity, diabetes, and liver steatosis. 31 Moreover, in our study, an increase in Fsp27/Cidec expression was associated with suppressed mitochondrial β-oxidation and the accumulation of TG, as well as an increase in lipid droplets. Similar to our findings (Fig. 5E), previous studies using an FFA-induced in vitro cellular model of NAFLD identified a dose-dependent effect of DLL-Ex in inhibiting HFD-induced increases in CD36. 18 These results indicated that the protective effect of DLL-Ex against NAFLD may be mediated via a decrease in hepatic FFA uptake.
Recently, it has been widely recognized that parallel modulation of multiple biological targets can be beneficial for the treatment of disease. 32 Since common disorders, such as metabolic disorders and cancer, tend to result from multiple molecular abnormalities rather than a single defect, combination drugs that impact multiple targets simultaneously are better at controlling complex disease systems. 33 Moreover, traditional medicines, such as Korean medicine, traditional Chinese medicine, and Indian Ayurveda, are being scientifically evaluated, and are becoming important resources for the discovery of bioactive molecules with therapeutic effects, as well as for designing multi-target drugs. 34 Dietary supplementations of functional herbal medicines have received increasing attention as adjuvants to attenuate the effects of metabolic diseases, including NAFLD. DLL-Ex is extracted from D. lablab beans and has been used for centuries in India and China as an edible pod and animal forage, as well as for treating diarrhea and other gastrointestinal diseases in traditional Korean medicine. 35
Recently, we reported that DLL-Ex ameliorated FFA-induced hepatic lipid accumulation in HepG2 cells, via activation of the AMPK cellular signaling pathway and by suppressing of the expression of genes involved in lipid droplet accumulation in hepatocytes. 32,36 Based on this evidence, we hypothesized that DLL-Ex could be very useful in the prevention of NAFLD. Identification of bioactive compounds from herbal extracts is a very important step in the pharmaceutical discovery process. An ultra-performance liquid chromatography quadrupole time-of-flight mass spectrometry (UPLC Q-TOF-MS) is the ideal analysis for herbal extract complexes since it provides the sensitivity and data density needed to allow the entire composition to be screened in a single analysis. We tentatively identified Lablaboside E, Lablaboside C, Lablaboside B, Azukisaponin III, Licoricesaponin J2, Sandosaponin, Chikusetsusaponin Iva, Soyasaponin Bb, Saponin D, Prosapogenin, and Monoglucuronylglycyrrhetinic acid (Fig. 6). These results explain, in part, the multi-target activity of DLL-Ex against NAFLD, which might include synergistic or additive effects of multiple active components in DLL-Ex. This hypothesis is supported by a recent report that multiple components in herbal extracts show synergistic effects to treat symptoms and can even result in novel pharmacological activities. 37 In conclusion, we showed that a novel herbal medicine, DLL-Ex, ameliorates body weight gain without inhibiting appetite in HFD mice. Daily oral supplementation of DLL-Ex attenuated hepatic fat accumulation and lowered blood levels of TG, cholesterol, and FFA, via repression of mRNA levels for genes involved in lipid import and lipid droplet accumulation. The sustained anti-NAFLD effects of DLL-Ex included alleviation of hepatic lipid accumulation, anti-obesity activity, decreased hepatic hypertriglyceridemia, and reduced FFA uptake. Taken together, our results provide additional understanding of the pharmacological function of a novel decoction, and support for the potential usefulness of DLL-Ex as an effective novel anti-NAFLD drug.
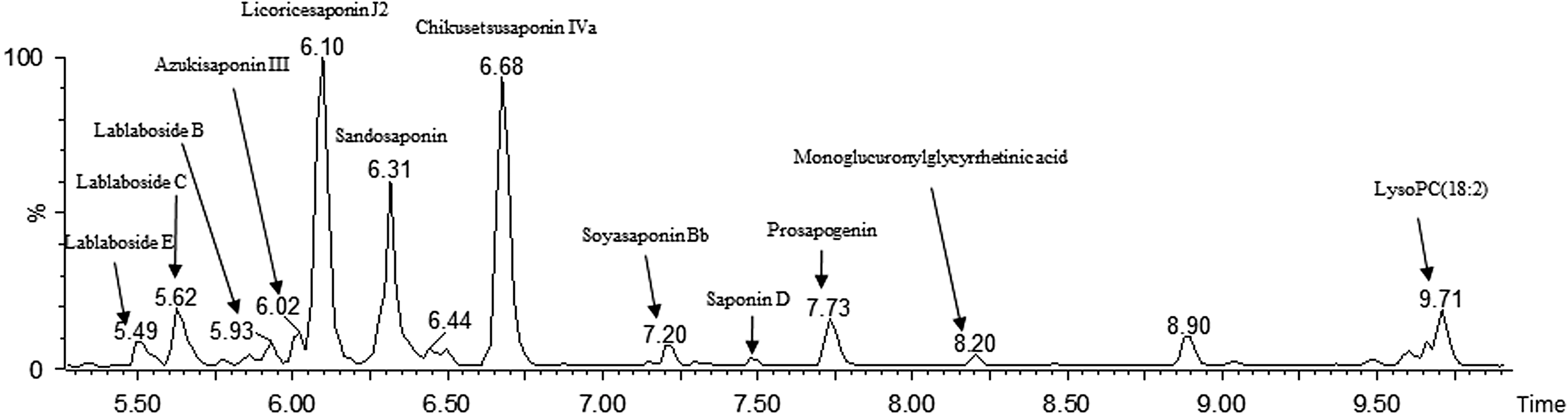
UPLC Q-TOF-MS chromatogram of DLL-Ex. UPLC Q-TOF-MS, ultra-performance liquid chromatography quadrupole time-of-flight mass spectrometry.
Ethics Approval and Consent to Participate
All experiments were performed in accordance with the guidelines for the Care and Use of Laboratory Animals developed by the Institute of Laboratory Animal Resources of the National Research Council, and they were approved by the Institutional Animal Care and Use Committee of Daejeon University in Daejeon, Korea.
Footnotes
Acknowlegments
This research was supported by the Development of Herbal Medicine for Aging Related NAFLD (K16302, K17300) funded by KM Convergence Research Division of Korea Institute of Oriental Medicine (KIOM). The author thanks Prof. Hoon Choi (Wonkwang University) and Dr. Hyuk Hwan Song (Korea National Food Cluster) for technical assistance with UPLC Q-TOF-MS analysis.
Author Disclosure Statement
No competing financial interests exist.