
Abstract
Licorice is extracted from the roots of plants in the Glycyrrhiza genus, especially Glycyrrhiza uralensis in China and Korea. It has several pharmacological activities, including neuro-protective, anti-fungal, and anti-cariogenic effects. Ischemia/reperfusion-induced brain injury is a leading cause of adult disability and death; thus, the identification of anti-apoptotic, neuro-protective therapeutic agents is viewed as an attractive drug development strategy. Infarct volumes and the expression of several apoptosis-related proteins, including Bcl-xL, Bcl-2, caspase-8, and caspase-9, were evaluated by western blotting in the brains of mice subjected to middle cerebral artery occlusion (MCAO). Three consecutive days of oral pretreatment with the methanol extract of licorice (GRex) significantly reduced infarct volumes 24 h after MCAO. In addition, GRex effectively inhibited the activation of caspase-9 by upregulating protein expression of Bcl-xL and Bcl-2. The neuro-protective effect of licorice was due to its regulation of apoptosis-related proteins. These data suggest that licorice could be a potential candidate for the treatment of ischemia-induced brain damage.
Introduction
S
Cerebral infarction, brain damage induced by reperfusion after ischemia, damages the local cerebral blood vessels and causes neuronal cell death and loss of brain tissue. 3,4 Animal models of cerebral infarction often involve cerebral arterial vasculature obstruction and embolization; the most commonly used is the middle cerebral artery occlusion (MCAO) model, in which the middle cerebral artery (MCA) is occluded with a suturing filament to block blood flow, causing ischemic stroke. 5,6 In the MCAO model, it is possible to either permanently or temporarily occlude blood flow to the MCA by controlling the duration of intra-luminal filament insertion. When the ischemic period is longer than 60 min, the boundary region of damaged brain tissue can be observed and quantified. Therefore, the MCAO model can be used to evaluate the effects and efficacies of neuroprotective drugs. 5,6
Licorice, the dried roots and rhizomes of Glycyrrhiza uralensis and Glycyrrhiza glabra, is a perennial plant belonging to the legume family, distributed in northeastern China, Siberia, and Mongolia. 7,8 In Chinese and Korean traditional medicine, licorice has been used to regulate the action of other medicines, and for various other purposes, including in food. 9 Among more than 20 species from the Glycyrrhiza genus, G. uralensis is used most commonly in Asian traditional medicine. 7 –11
Numerous studies have reported the different phytochemicals of the Glycyrrhiza species and their bioactivities. 10,12,13 Recently, some phytochemicals isolated from licorice were reported to have neuro-protective potential. 14 –16 In our preliminary study, a chemical from G. uralensis showed neuro-protective potential in a mouse MCAO model. Therefore, in this study, we confirmed the protective effects of the methanol extract of licorice from the dried roots and rhizomes of G. uralensis in the MCAO model of stroke.
Materials and Methods
Methanol extract of licorice
The licorice used in this study was purchased from a commercial pharmaceuticals company (Gwangmyoung Pharmaceuticals Co., Korea) available in markets under the medicinal name of Glycyrrhiza Radix. To validate the quality of the herbal material, fingerprinting data of the licorice imported from Kazakhstan were kindly provided by Gwangmyoung Co., and the data are shown as Figure 1. The quality and origin were clearly confirmed through external morphological observation, and the sample was stored at −4°C. To prepare the methanol extract, 200 g of licorice was placed in 2000 mL of methanol and incubated at room temperature for 5 days. Then, the supernatant liquid was removed, and 1000 mL of methanol was added to the filtered licorice residue and filtered again. The supernatant liquid was obtained two times and filtered through filter paper; it was then subjected to vacuum concentration and freeze-drying. The final weight of lyophilized methanol extract of licorice (GRex) was 17 g (yield: 8.5%). The GRex was stored in micro tubes at −20°C until use.
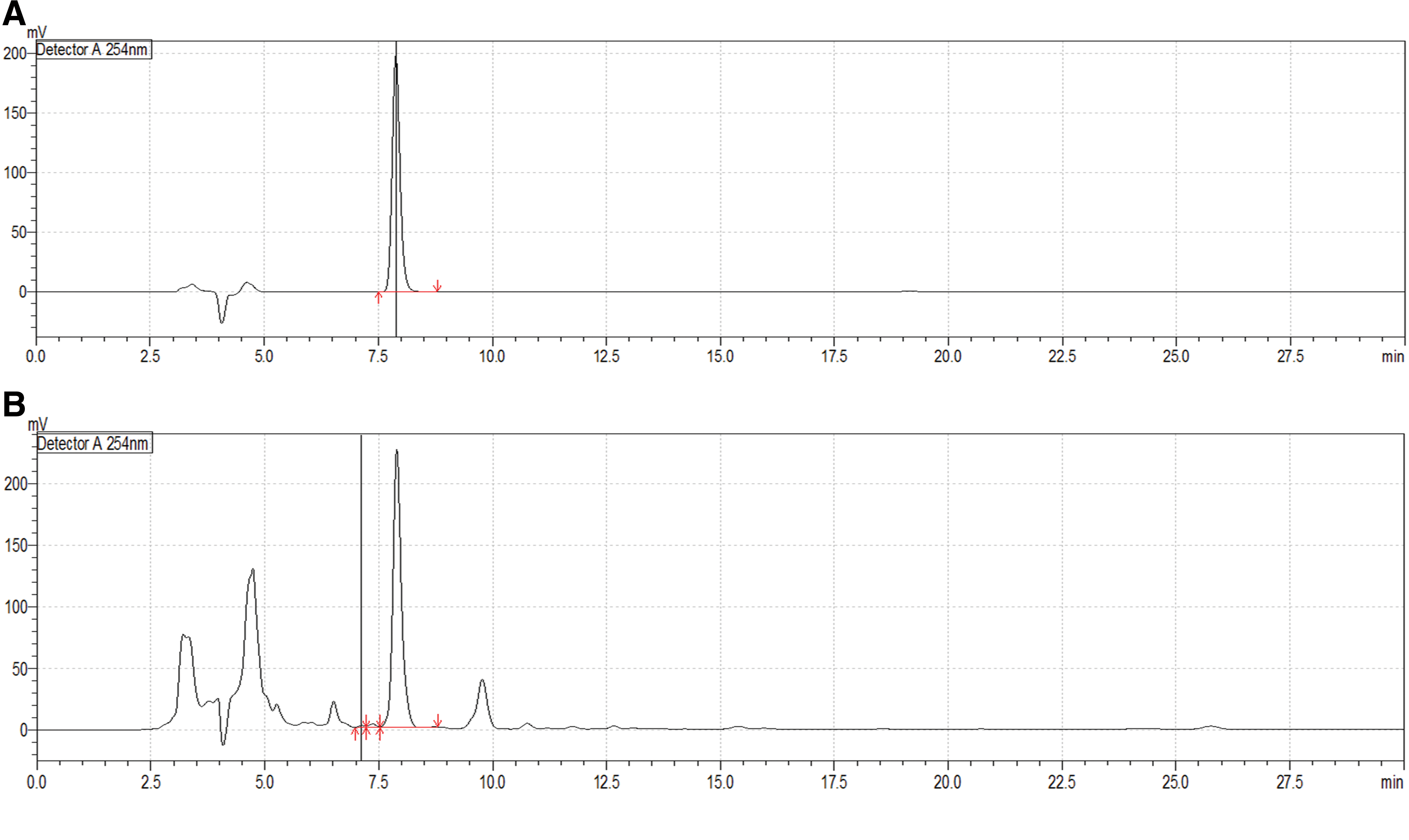
HPLC images of the licorice and its standard compound, glycyrrhizic acid.
Animals
Male C57BL/6 mice aged 6 weeks and weighing 22–25 g (BioLink, Korea) were adapted to the laboratory environment for 1 week before use. All animals had free access to standard chow and water and were housed in an environment with controlled temperature (22°C ± 1°C) and light cycle (12 h light/dark). The animal experiment protocol was approved by the ethics committee of Pusan National University (Approval No. PNU-2016-1087). The experimental groups included the sham-operated normal group, control group, and treatment groups with three concentrations of GRex, with each containing at least seven animals per experimental group. All procedures were approved by Pusan University Animal Experimental Ethics Committee, and each experimental procedure was conducted in compliance with related provisions.
Administration of GRex
GRex was dissolved in dimethyl sulfoxide (DMSO), diluted with 0.9% physiological saline, and finally filtered through a 0.45 μm syringe filter. The final volume of administration was adjusted to 5 mL/kg body weight. GRex was administered once a day for 3 days at doses of 50, 150, and 500 mg/kg body weight by oral gavage. The normal group and control group were orally administered equivalent amounts of DMSO and physiological saline.
Animal model
In the control group and GRex treatment groups, surgery for the MCAO model was performed as previously described, 17 with several modifications described later. Anesthesia was induced by inhalation of 2% isoflurane in 70% N2O and 30% O2, and body temperature was maintained at 36.5°C ± 0.5°C by using a body temperature-holding blanket to which a thermometer was connected. The hair on chest and neck were removed with clippers, and an incision was made in the center of the neck. The left common carotid artery (LCCA), external carotid artery, and branch of the internal carotid artery were carefully isolated from surrounding connective tissues. The external carotid artery and the common carotid artery were ligated with a surgical suture to block blood flow during the operation, and the blood flow in the internal carotid artery was temporarily blocked. In this state, a silicon-coated nylon suture (8-0 monofilament; Ethicon, Scotland) ∼11 mm in length was inserted slowly through the internal carotid artery to the origin of the left MCA. MCAO was confirmed by a decrease in relative cerebral blood flow (rCBF) in the MCA to 20% of normal. The blood flow was measured by a Laser Doppler Flowmeter (moorVMS-LDF; Moor Instrument, United Kingdom). In brief, an optic fiber was securely fixed with surgical glue onto the skull where the left MCA is located, and rCBF before LCCA ligation was considered as 100%.
The inserted filament was fixed to the blood vessel for 2 h during which the cerebral artery was occluded; then, the filament was carefully withdrawn to restore the blood flow for 22 h of reperfusion. The skin was sutured, and each mouse was awakened from anesthesia. In the normal group, a sham operation proceeding to the common carotid artery isolation was performed in which the common carotid artery was ligated and the incised muscle and the skin were sutured. Drug administration, induction of ischemia and perfusion, and sacrifice of laboratory animals are shown in Figure 2.
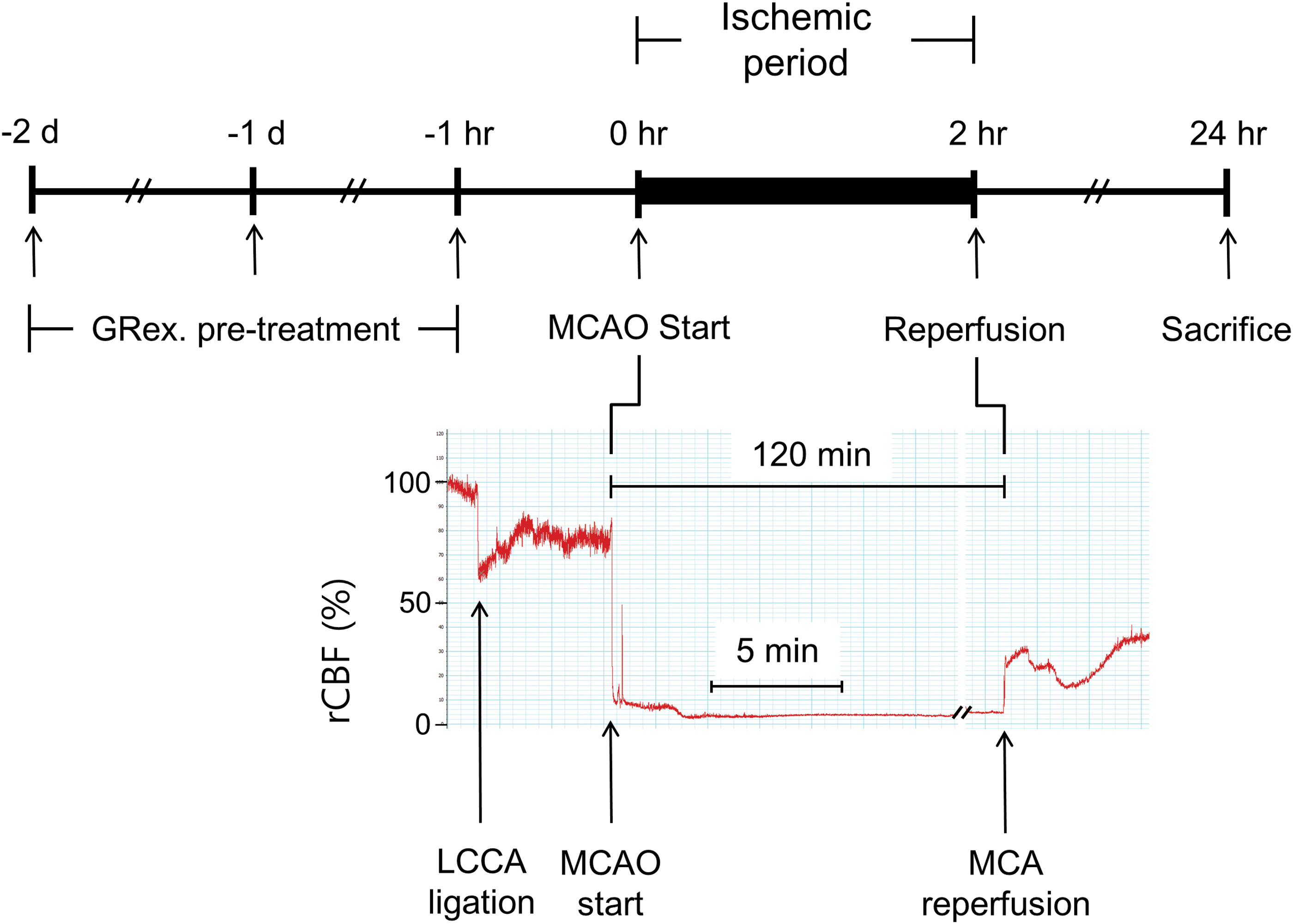
Establishment of MCAO model. Mice were pretreated for 2 days, 1 day, and 1 h before MCAO with three concentrations of GRex, and they were sacrificed 24 h after the start of MCAO. MCAO was maintained for 2 h. After mice were sacrificed, harvested brain slices were kept in a deep freezer for protein assay or stained with TTC solution for infarct measurement. GRex, methanol extract of licorice; LCCA, left common carotid artery; rCBF, relative cerebral blood flow. MCAO, middle cerebral artery occlusion; TTC, 2,3,5-triphenyltetrazolium chloride. Color images available online at
Analysis of damaged area of brain tissue
Twenty-four hours after the start of MCAO, the experimental animals were euthanized with CO2, and brains were removed and stored on ice. The excised brains were sectioned at a thickness of 1 mm by using a mouse brain matrix (Kent Scientific, USA), and they were then stained in a solution of 2% 2,3,5-triphenyltetrazolium chloride (TTC). Subsequently, the sections were fixed in 10% formalin for at least 2 h and photographed with a digital camera. TTC stains viable tissue red, whereas the necrotic area remains white. The cerebral infarct area of each section was quantified and analyzed by using ImageJ (NIH, USA).
Neurobehavioral evaluation
To confirm the extent of damage to motor ability in animal models of stroke, neurobehavioral function was evaluated 24 h after MCAO was applied, and the extent of injury was quantified as follows by using a 5-point scale.
Grade 0: There is no neurological abnormality.
Grade 1: Spontaneous exercise is performed smoothly, but forepaws cannot smoothly grasp surroundings.
Grade 2: Spontaneous exercise is performed relatively easily, but as mice were pulled by tail and body was placed to a flat bottom, the body shifted to the right.
Grade 3: Walking or circling to the right is done when stimulated, and animals respond sensitively to pain sensation when stimulating the tail.
Grade 4: Spontaneous exercise is not performed, and animals respond very sensitively to pain sensation when stimulating the tail.
Detection of proteins in brain tissue
Experimental animals were euthanized with CO2 24 h after MCAO was initiated, and the brains were removed and placed on ice. The left hemisphere of the excised brain, including the putative penumbra, was isolated and incubated in lysis buffer (1% Triton X-100, 0.32 M sucrose, 5 mM EDTA, 1 mM DTT, 10 mM Tris, pH 7.4). After removing the residue with a centrifugal separator, protein concentration was determined. Equal amounts of protein were dissolved in sample buffer and denatured by heating the proteins in lysis buffer with heating block for 5 min. Proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to nitrocellulose membrane (Whatman, United Kingdom). The membrane was blocked in 5% skim milk in tris-buffered saline with tween buffer at room temperature for 1 h. Then, membranes were incubated in primary antibodies for Bcl-xL, Bcl-2, caspase-8, and caspase-9 (1:1000 dilution) or β-actin (1:2000 dilution) at 4°C for 12 h. Next, the membranes were incubated with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG, pAb (1:5000) or HRP-conjugated goat anti-mouse IgG pAb (1:3000) for 2 h at room temperature. The membrane was developed with an ECL kit (GenDEPOT, USA) on film in a dark room. Protein concentration was quantified and compared with β-actin by densitometry using ImageJ software (NIH).
Statistical analysis
Experimental results were expressed as means ± standard deviation, and the statistical significance among the groups was determined by using one-way ANOVA test followed by Tukey's post hoc analysis using Sigmaplot v12.0 software. Statistical significance was determined when the P-value was <.05.
Results
Effects on damaged brain area and neurobehavioral changes
Ten 1 mm brain tissue sections were obtained from the brain, excluding the olfactory bulb and cerebellum. TTC staining was used to identify infarct volume and to obtain optical images of the damaged area (Fig. 3A). In the sham-operated normal group, no cerebral infarct was observed; in contrast, a relatively wide range of damaged area was observed in the control group. In the mice that received GRex, a significant reduction in damaged area was observed, whereas only mice that received 500 mg/kg GRex had a statistically significant reduction in damaged area (Fig. 3B).
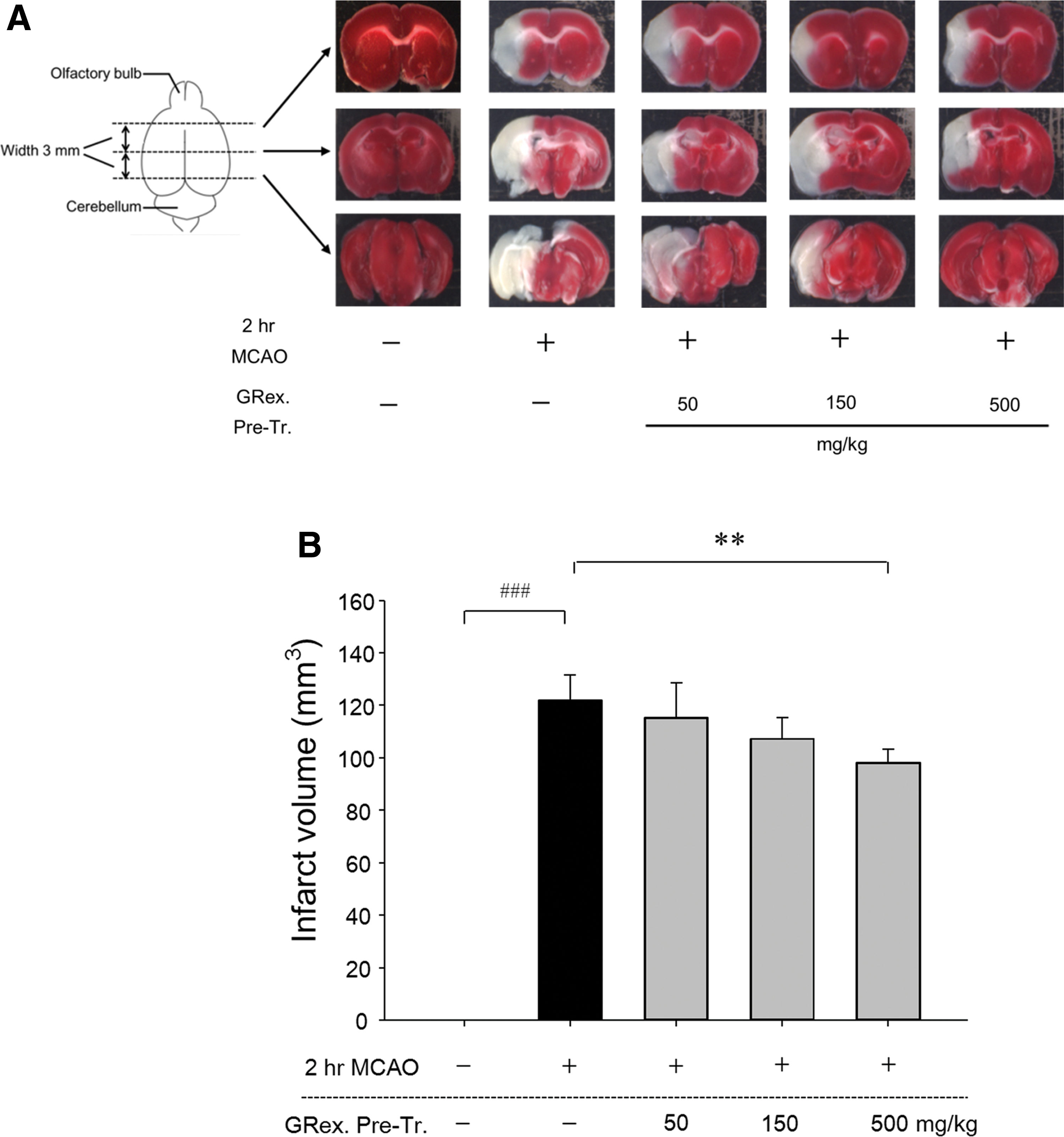
Effects of GRex on infarct volume, and representative images of each group. TTC staining was used to identify infarct volume and to obtain optical images of the damaged area. Pretreatment of 500 mg/kg GRex significantly reduced infarct volume (P < .01).
MCAO caused a significant increase in neuro-behavioral changes in the control group compared with the sham-operated normal group. However, GRex administration had no effect on neuro-behavioral changes compared with the control group (Fig. 4).
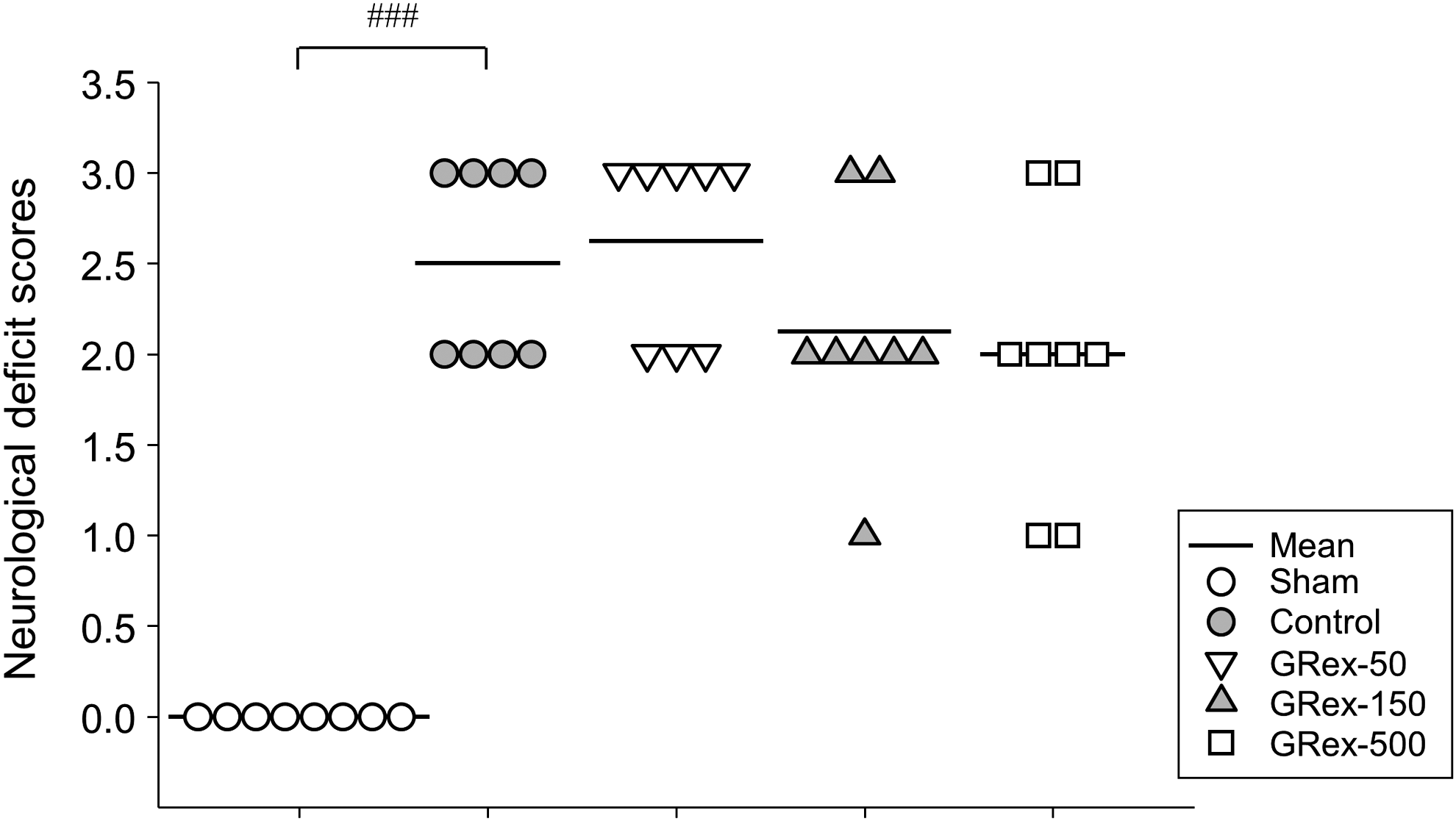
Effects of GRex on neurological deficit scores. Neurobehavioral function was evaluated 24 h after MCAO was applied to confirm the extent of damage to motor ability in animal models of stroke. GRex pretreatment showed no improvement on neuronal deficit scores. Data are presented as dot density plot and mean values (n = 7). ### P < .001 versus normal mice.
Influence of GRex on proteins related to brain tissue apoptosis
GRex was administered daily for 3 days at concentrations of 50, 150, and 500 mg/kg body weight, and then brain infarction was induced by MCAO. Mice were sacrificed 24 h after the onset of cerebral ischemia, and protein expression at the penumbra site was evaluated by western blotting to determine the effect of GRex on apoptosis induced by ischemia/reperfusion injury. We found a decrease in the expression of Bcl-xL in the control group that underwent MCAO compared with that in the normal group (Fig. 5A). Importantly, the expression of Bcl-xL increased in a concentration-dependent manner in mice that were pretreated with GRex. A statistically significant increase in Bcl-xL was observed in the groups administered GRex at concentrations of 150 and 500 mg/kg body weight (Fig. 5A).

Effects of GRex on Bcl-xL, caspase-9, and Bcl-2 expression in the brains of MCAO-induced mice. Expression of Bcl-xL, caspase-9
Expression of caspase-9 protein, one of the proteins involved in the pathways downstream of Bcl-xL, was increased by MCAO, showed a tendency to decrease in a concentration-dependent manner when GRex was administered, and reached statistical significance in the group that received 150 mg/kg GRex (Fig. 5B). The expression of Bcl-2 protein, a suppressor of apoptosis in a variety of cells, was decreased by MCAO and showed a tendency to increase in a concentration-dependent manner when GRex was administered (Fig. 5B).
Bcl-xL, Bcl-2, and caspase-9 belong to the intrinsic pathway of cell death, and caspase-8 is a protein belonging to the extrinsic pathway. In this study, several proteins in the intrinsic pathway of apoptosis were upregulated; however, there was no change in the expression of caspase-8 due to MCAO or GRex administration (Fig. 6).

Effects of GRex on caspase-8 expression in the brains of MCAO-induced mice.
Discussion
Stroke is a neuro-vascular disease caused by oxidative damage to the cells constituting the brain tissue and inflammation of the brain and nervous system, and it results in deficits to movement and language or death in severe cases. 18,19 With the increasing prevalence of metabolic diseases such as chronic hypertension, diabetes, and hyperlipidemia, which are major risk factors for stroke, the prevention and treatment of stroke are important areas of medical research. 20,21 Moreover, the deficits in language and movement due to stroke have a strong correlation with the degree of damage to brain tissue 22 and result in a poor quality of life for patients and their families; therefore, research on therapeutic agents that can prevent and treat stroke is critical. 19
It is important to use an animal model of stroke that involves the same pathological changes as human disease to study the efficacy of drugs for the treatment of cerebral infarct and damage due to ischemia. The MCAO model mimics thrombotic stroke by obstructing cerebral arterial vessels, is relatively reproducible, and is recognized as a relatively noninvasive research method; therefore, it is commonly used. 23 –26
Numerous studies have recently reported that licorice or its components have pharmacological activities, including antitumor, antimicrobial, and anti-inflammatory effects. 10 –12 Previous reports demonstrated that licorice and its components have pharmacological potential in ischemic brain injury 14 –16,27 ; however, its mechanism of action in an animal model was still unclear. In this study, we confirmed that 3 days of GRex pretreatment attenuated brain damage induced by 2 h of ischemic injury followed by reperfusion (Fig. 3).
During ischemic stress, blood is not supplied to the brain, resulting in cerebral infarction caused by cell death due to necrosis and apoptosis. In the penumbral region where apoptosis progresses, cells can die or survive depending on the compatibility of the treatment method. 18,20 –22,28 The Bcl-2 family, including Bcl-xL and Bcl-2, and multiple caspase proteins have important regulatory actions in the process of apoptosis; Bcl-xL and Bcl-2 proteins suppress apoptosis, and Bax and Bak proteins induce apoptosis. 29,30 In this study, we found that pretreatment with GRex significantly increased the expression of Bcl-xL and Bcl-2 in the penumbra area compared with that in the control group (Fig. 5). These results correlated with a reduction in the damaged area by GRex pretreatment (Fig. 3), indicating a possibility that GRex induces Bcl-xL/Bcl-2 expression, which leads to neuronal cell survival by suppressing the apoptotic pathway. To confirm this, the expression of caspase-8 and caspase-9 proteins was also measured.
Caspase-8 and caspase-9, the initiator caspases, facilitate apoptotic cell death by activating other proteins such as caspase-3, -6, and -7, which directly initiate apoptosis. 31 –33 Caspase-8 acts on the extrinsic pathway, whereas caspase-9 acts on the intrinsic pathway. 31,32 In this study, GRex administration had no effect on caspase-8 expression (Fig. 6), but caspase-9 expression was effectively regulated by GRex administration (Fig. 5A). This result indicates that the neuro-protective effects of GRex could be mediated by an anti-apoptotic mechanism via the intrinsic pathway (Fig. 7). Recently, numerous researchers have reported that brain tissue damage due to ischemia/reperfusion may be improved by inhibition of caspase-3. 33,34 Our study confirmed that GRex administration can suppress the damage to brain tissue by regulating signals upstream of caspase-3.

Schematic image of the anti-apoptotic action mechanism of GRex in MCAO mice model. Thickened red bars indicated the predicted action site of GRex pretreatment on ischemia/reperfusion-induced brain damage. Color images available online at
In summary, we have demonstrated that the neuro-protective effect of licorice is possibly related to the suppression of ischemia/reperfusion-induced brain damage by modulating the proteins involved in the intrinsic pathway of apoptosis.
In conclusion, to investigate the neuro-protective effects of licorice on stroke, GRex, a methanol extract of licorice, was administered to mice for three consecutive days; the MCA was occluded for 2 h to induce cerebral infarction; and finally, several biological indicators of stroke were evaluated. GRex pretreatment significantly reduced infarct volume but had no effect on neurobehavioral changes. GRex significantly inhibited the decrease in the expression of Bcl-xL and Bcl-2, and the changes affected the expression of caspase-9 protein. Thus, licorice appears to have an anti-apoptotic effect and may be neuro-protective in stroke.
Footnotes
Author Disclosure Statement
No competing financial interests exist.