
Abstract
The vascular tone plays an important role in blood pressure and flow. It is influenced by the contraction of vascular smooth muscle cells (VSMCs), which in turn is regulated by the balance between the myosin light chain kinase (MLCK) and the phosphorylated myosin light chain (p-MLC). Quercetin is a common flavonoid which is found in many fruits and red wine. Although quercetin has been widely reported to be involved in cell proliferation, migration, and apoptosis in VSMCs, it has not yet been demonstrated whether quercetin is related to vasocontraction, a function regulated by the AMP-activated protein kinase (AMPK) signaling pathway. Accordingly, the aim of this study is to investigate the molecular mechanism through which the quercetin-activated LKB1-AMPK signaling pathway regulates the contraction of VSMCs. In cultured VSMCs, quercetin activated AMPK in a dose- and time-dependent manner. Quercetin inhibited the phenylephrine (PE)-induced expression of MLCK and p-MLC through the LKB1-AMPK signaling pathway and decreased the mRNA level of MLCK. Adenovirus-AMPK DN α1 and AMPK DN α2-transduced VSMCs displayed higher p-MLC expression. Moreover, quercetin inhibited the PE-mediated contraction in rat aorta. These data suggest that the quercetin-activated LKB1-AMPK signaling pathway regulates VSMC contraction by inhibiting MLCK and p-MLC; hence, it may be a therapeutic intervention for the treatment of cardiovascular disorders such as atherosclerosis and hypertension.
Introduction
A
The contraction of smooth muscles is regulated by the balance between the activation of the myosin light chain kinase (MLCK) and the phosphorylated myosin light chain (p-MLC). 7 In vascular smooth muscle cells (VSMCs), Ca2+ is released in the interior of cells where it forms Ca2+-calmodulin complexes. 8 These complexes interact with MLCK, which then phosphorylate myosin light chains. Myosin and actin pull each other, hereby causing smooth muscle contraction.
Flavonoids have received attention as promising preventive and therapeutic agents for various diseases such as cardiovascular disorders and diabetes. 9,10 Quercetin is a common flavonoid which is found in many fruits, vegetables, red wine, and tea. 11 It is known that quercetin has antioxidative, anti-inflammatory, and anticancer activity. 11 –13 Previous studies reported that quercetin strongly inhibits cell proliferation and induces apoptosis through the AMPK signaling pathway in breast and bladder cancer cells. 14,15 In colon cancer cells, quercetin reportedly regulates the Sestrin 2-AMPK-mTOR signaling pathway and induces apoptosis by increasing intracellular ROS. 16 In addition, quercetin has been shown to protect from oxLDL-induced endothelial dysfunction. 17 It also exerts cardioprotective activity by inhibiting the infiltration of inflammatory cells to the site of cardiac infarction. 18
Although quercetin has been widely reported to be involved in cell proliferation, migration, and apoptosis in VSMCs, 19,20 it has not yet been demonstrated whether quercetin is related to vasocontraction, a function regulated by the AMPK signaling pathway in VSMCs. Therefore, we investigated whether quercetin can induce vasorelaxation through AMPK activation in VSMCs. Our results confirmed this induction and verified the involvement of quercetin in the vasorelaxation mechanism.
Materials and Methods
Reagents and antibodies
Quercetin, phenylephrine (PE), and anti-MLCK were purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies against LKB1, p-LKB1, AMPK, p-AMPK, p-acetyl CoA carboxylase (ACC), and p-MLC were purchased from Cell Signaling Technology (Beverly, MA, USA). Anti-CaMKKβ was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA) and anti-α-smooth muscle actin from Abcam (Cambridge, MA, USA). Compound C, an AMPK inhibitor, was obtained from Calbiochem (La Jolla, CA, USA). AMPK, LKB1, and control siRNA were purchased from Santa Cruz Biotechnology.
Cell culture
Sprague-Dawley rats were euthanized with 95% CO2. VSMCs were isolated from the thoracic aorta and cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 50% fetal bovine serum and 1% antibiotics in a CO2 incubator (5% CO2/95% air, 37°C). We used VSMCs from passages 5 to 8 at 70–90% confluence in 10 cm dishes, whose growth was arrested by incubation in serum-free DMEM for 24 h before use.
Aortic ring preparation and organ bath experiments
We euthanized 8-week-old male Sprague-Dawley rats with 95% CO2. The thoracic aorta was dissected rapidly, and 2–3 mm aortic rings were prepared. The endothelial lining was removed by gentle rubbing. The rings were suspended isometrically between two stainless steel hooks in organ chambers containing K-H solution at 37°C, and the top hook was connected to a force-displacement transducer connected to an amplifier (Biopac Systems, Inc., Goleta, CA, USA). The normal bathing solution contained the following (g/L): NaCl 6.9, KCl 0.35, MgSO4 0.14, NaHCO3 2.09, KH2PO4 0.16, glucose 1, and CaCl2 0.29 (pH 7.4). This solution was gassed constantly with 95% O2 and 5% CO2 and maintained at 37°C. The aortic rings were equilibrated for 30 min before the initiation of the experiment.
Western blot analysis
Whole cell extracts were prepared by lysis in a PRO-PREP protein extract buffer. The protein concentration was quantified with the protein assay reagent from Bio-Rad (Hercules, CA, USA). Equal amounts of protein were mixed with Laemmli Sample Buffer (Bio-Rad) and heated for 5 min at 100°C before loading. Total protein samples (30 μg) were subjected to 10% SDS-polyacrylamide gel electrophoresis for 90 min at 100–120 V. Separated proteins were electrophoretically transferred to a PVDF membrane for 80 min at 100 V using the SD Semi-dry Transfer Cell. Membranes were blocked with 5% nonfat milk in phosphate-buffered saline (PBS) containing 0.05% Tween 20 (PBS-T) for 1 h at room temperature and then incubated with the primary antibodies at a dilution of 1:1000 overnight at 4°C in PBS-T. Membranes were washed in four changes of wash buffer (PBS-T) and incubated for 1 h at room temperature in PBS containing anti-rabbit (Stress-gen, Ann Arbor, MI, USA) or anti-mouse IgG (Sigma-Aldrich). After three more rinses with PBS-T, membranes were exposed to enhanced chemiluminescence (ECL) and ECL Plus western blot analysis detection reagents.
Transfection of siRNA
Transfection of VSMCs with siRNA was performed using the Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. VSMCs were plated on six-well plates the day before transfection and grown to about 70% confluence. Cells were then transfected with 10 μM control, LKB1, or AMPK siRNA, and 100 pmol of Lipofectamine for 7 h in Opti-MEM I reduced-serum medium (Invitrogen). After an incubation period of 48 h, protein levels were measured by western blot analysis.
Adenoviral transduction
Adenoviruses expressing the control gene GFP, or a constitutively active form of AMPK (AMPK-CA), or the dominant-negative isoform of the α1 and α2 subunits of AMPK (AMPK DN α1 and α2) were amplified in AD293 cells using standard methodologies. The viruses were transduced to VSMCs in serum-free DMEM for 6 h.
RNA isolation and real-time polymerase chain reaction analysis
Total RNA was isolated from cultures using TRIzol reagent (Invitrogen), and reverse transcription was performed using the SuperScript III Reverse Transcriptase (Invitrogen). We then determined the MLCK mRNA levels in VSMCs using real-time polymerase chain reaction (PCR). The primers for the PCR analysis were synthesized by Macrogen (Daejeon, Korea) and were as follows: MLCK, forward 5′-AATGGTGTTGCTGGAGATCGAGGT-3′ and reverse 5′-GCTGGATCAAATTGCGGTGGTTCA-3′; Human GAPDH, forward 5′-ACAAGATGGTGAAGGTCGGTGTGA-3′ and reverse 5′-AGCTTCCCATTCTCAGCCTTGACT-3′. Real-time PCR was performed using the QuantiTect SYBR Green Kit (Applied Biosystems, Foster City, CA, USA) according to the manufacturer's instructions, and the total reaction volume was 20 μL. Reaction conditions were as follows: an initial denaturation step at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 sec, annealing at 57°C for 20 sec, and elongation at 72°C for 1 min. Fluorescence was detected using a Bio-Rad Real-Time PCR Detection System. The ratios of target gene to GAPDH mRNA were calculated.
Immunofluorescence analysis
VSMCs were seeded on coverslips in 35 mm dishes, fixed in 4% formaldehyde, and permeabilized with 0.2% Triton X-100. The MLCK primary antibody was used at 1:100 and incubated with cells overnight at 4°C. Mouse FITC secondary antibody (Invitrogen) was used at 1:1000 and incubated with cells for 1 h at room temperature. After fixation and immunofluorescent staining, cells were imaged under a fluorescence microscope (Olympus, Tokyo, Japan). The p-MLC primary antibody and α-smooth muscle actin were used at 1:50 and 1:100, respectively, and then incubated with cells overnight at 4°C. FITC secondary antibodies (Molecular Probes, Eugene, OR, USA) were used at 1:100 and incubated with cells for 1 h at room temperature. After fixation and immunofluorescent staining, cells were imaged under a confocal microscope (Leica, Bannockburn, IL, USA).
Statistical analysis
All data are expressed as mean ± standard error from at least three independent experiments. Differences between data sets were assessed by one-way analysis of variance of Bonferroni's t-test, with P values <.05 considered significant.
Results
Quercetin increased the phosphorylation of AMPK and LKB1 in VSMCs
Treatment of VSMCs for 6 h with different concentrations of quercetin (25, 50, and 100 μM) led to a dose-dependent increase in the phosphorylation of not only AMPK but also ACC, which is located downstream of AMPK (Fig. 1A, B). Moreover, compared with the control, quercetin (50 μM, 6 h) caused a significant increase in the phosphorylation levels of both AMPK and ACC (Fig. 1C, D). As previous studies have identified LKB1 and CaMKKβ as upstream elements of AMPK signaling, we next investigated whether quercetin affected these two proteins. Results showed that quercetin increased both LKB1 phosphorylation and CaMKKβ expression (Fig. 1E, F). Moreover, p-LKB1 and CaMKKβ increased significantly with increasing time of exposure to quercetin (Fig. 1G, H). The difference between LKB1 and CaMKKβ was that LKB1 phosphorylation could be observed at a lower concentration of quercetin than CaMKKβ expression. Moreover, the maximum increase of p-LKB1 was observed at an earlier time than CaMKKβ expression. These data suggested that quercetin dose- and time-dependently induced the activation of AMPK, ACC, LKB1, and CaMKKβ.
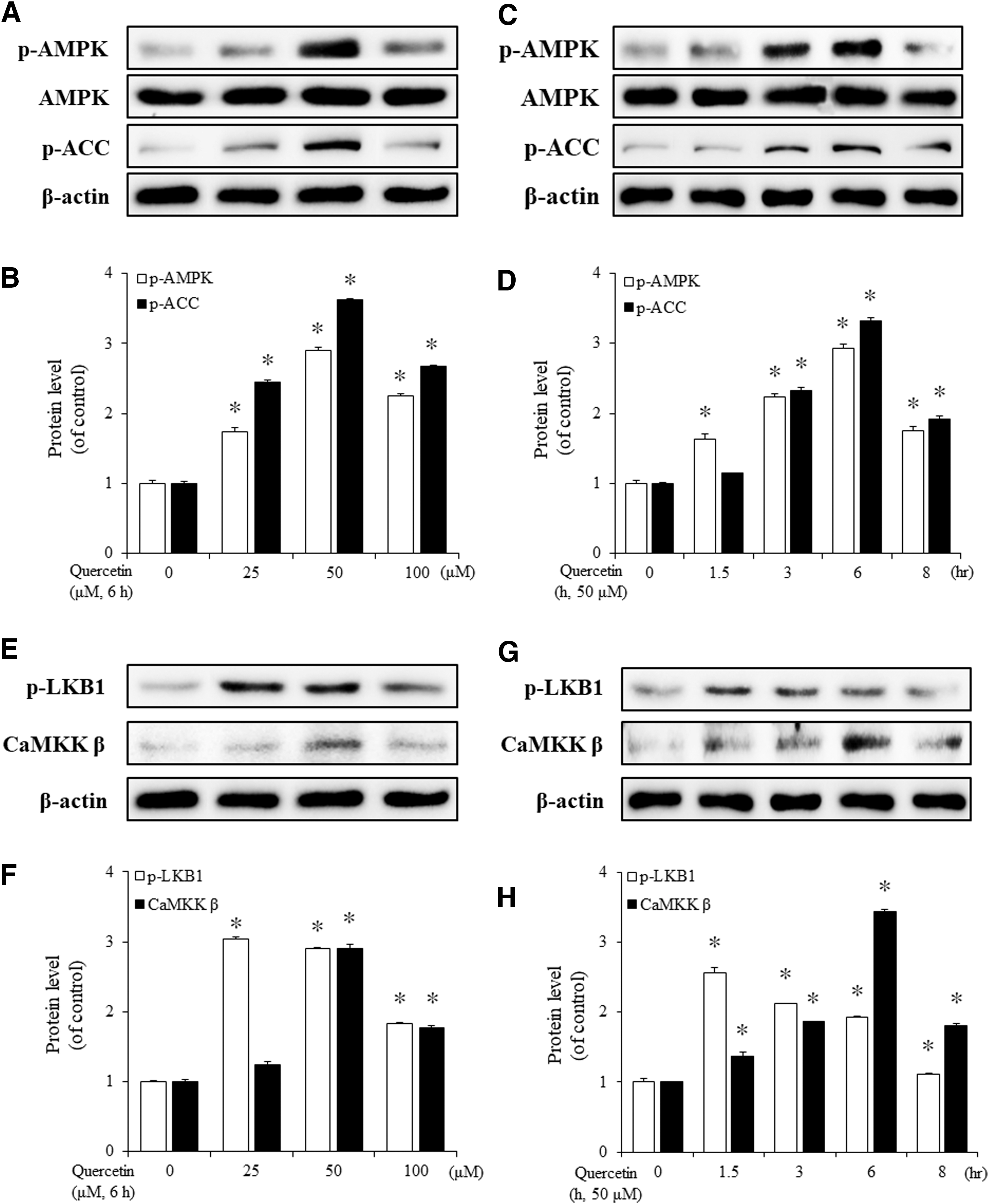
The effect of quercetin on AMPK and LKB1 phosphorylation in VSMCs. Cells were incubated with different concentrations of quercetin (25, 50, and 100 μM) for 6 h
Quercetin attenuates vascular contraction and inhibits the PE-induced increases in the expression of MLCK and p-MLC
We first investigated whether quercetin could regulate PE-induced vasocontraction in rat aorta. As shown in Figure 2A, quercetin inhibited vasocontraction, while pretreatment with compound C (10 μM) alleviated this effect. We next tested whether the AMPK signaling pathway was involved in the PE-induced expression of MLCK and p-MLC by treating rat aorta with PE and/or quercetin and then measuring the protein levels of MLCK and p-MLC using western blot analysis. Treatment with PE (1 μM) alone significantly increased the levels of MLCK and p-MLC. However, pretreatment with quercetin (50 μM) inhibited these effects (Fig. 2B, C). In contrast, treatment with quercetin increased not only AMPK phosphorylation but also activated LKB1 (Fig. 2D, F). Taken together, these results show that quercetin inhibited PE-induced vasocontraction through the LKB1-AMPK signaling pathway.
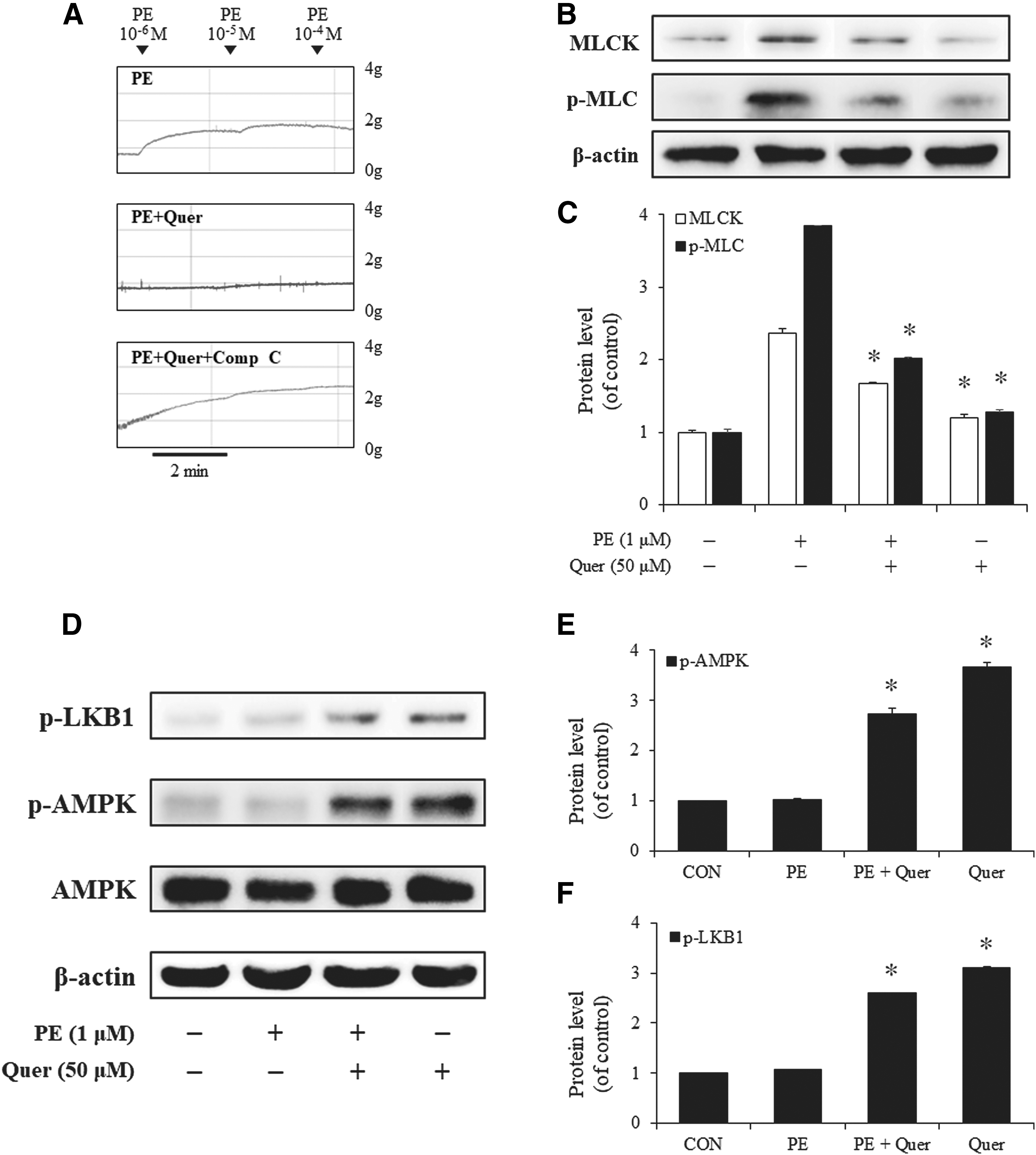
The effects of quercetin on the PE-induced contraction of rat aorta and the expression of MLCK and p-MLC.
Inhibition of AMPK activation restores the PE-induced increases in MLCK and p-MLC levels
To determine the potential effects of quercetin, we investigated whether compound C, a chemical inhibitor of AMPK, affected the PE-induced increases in the expression of MLCK and p-MLC. Although treatment with quercetin (50 μM) decreased the protein levels of MLCK and p-MLC, treatment with compound C (10 μM) restored MLCK and p-MLC to levels achieved by treatment with PE (1 μM) alone (Fig. 3A–C). We also checked the mRNA level of MLCK by q-PCR. As seen in Figure 3D, the result was consistent with the western blot analysis. Finally, we performed immunostaining with a specific antibody against MLCK. The results of the immunofluorescence experiment confirmed that MLCK expression was inhibited by quercetin but restored by compound C (Fig. 3E). Taken together, these results suggested that quercetin modulates the PE-induced MLCK and MLC phosphorylation through AMPK activation in VSMCs.
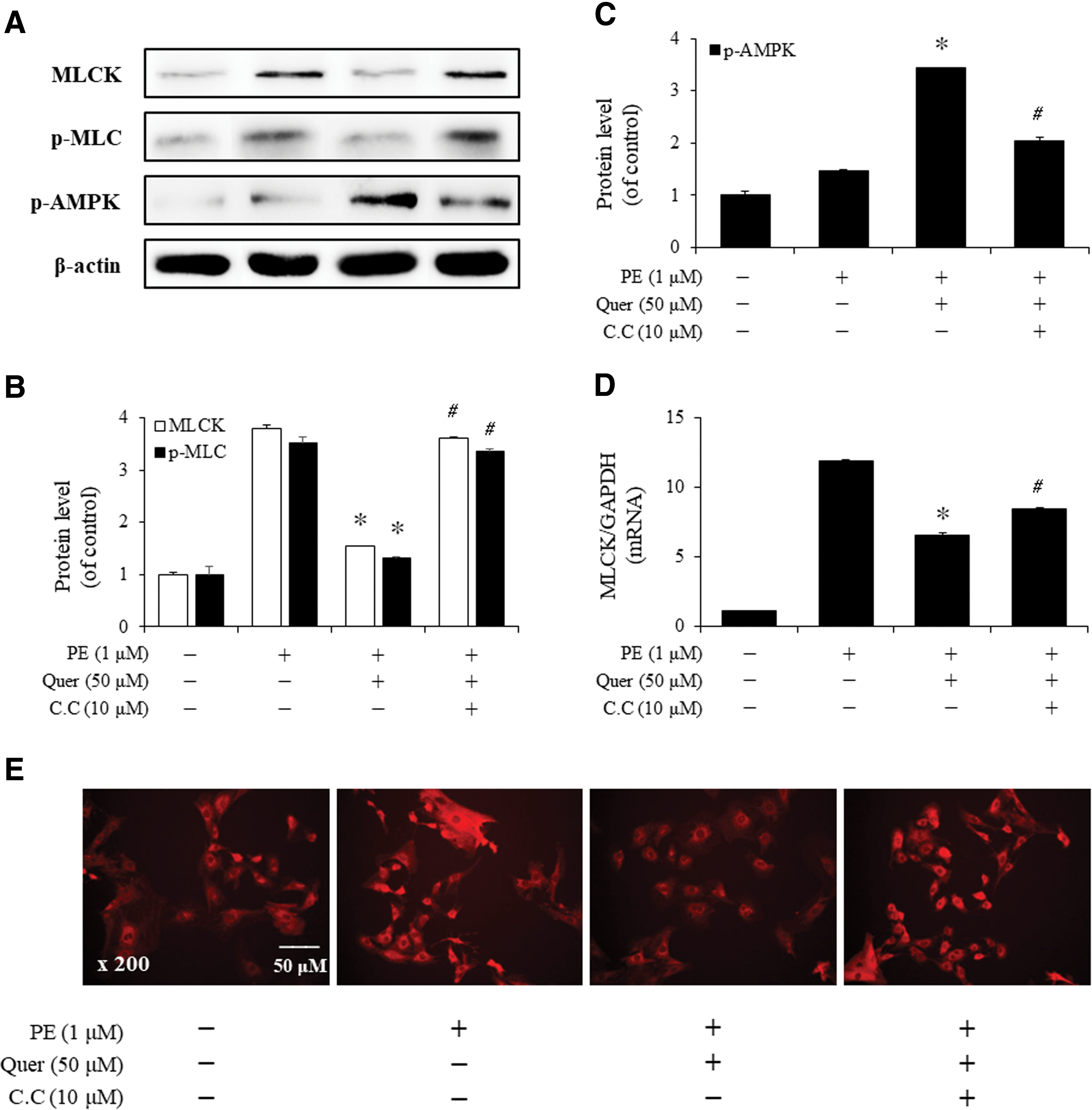
The inhibitory effect of quercetin-induced AMPK on the expression of MLCK and p-MLC in VSMCs. Cells were incubated in the presence or absence of PE (1 μM, 30 min) after pretreatment with quercetin (50 μM, 6 h) alone or quercetin + compound C (C.C; 10 μM, 1 h).
Quercetin regulates the expression of MLCK and p-MLC through the LKB1-AMPK signaling pathway
Since LKB1 phosphorylated AMPK, we determined whether quercetin regulates the expression of MLCK and p-MLC through the LKB1-AMPK signaling pathway in VSMCs. As shown in Figure 4A, B, transfection of VSMCs with LKB1 or AMPK siRNA resulted in increased expression of MLCK and p-MLC compared with control siRNA-transfected cells. q-PCR analysis of MLCK mRNA levels revealed consistent results (Fig. 4C). Furthermore, we transduced VSMCs with an adenovirus-mediated overexpression system and then investigated protein levels by western blot analysis. In contrast to the adenovirus-GFP control, the transduction of AMPK DN α1 and DN α2 increased the expression of MLCK and p-MLC by decreased p-AMPK (Fig. 4D, E). Moreover, as vascular contraction results from the interaction between myosin and actin in smooth muscle cells, VSMCs were transduced with adenovirus and then stained with Alexa Fluor 488-labeled anti-p-MLC (green) and Alexa Fluor 546-labeled anti-α-smooth muscle actin (red). Yellow staining indicated colocalization of p-MLC and α-smooth muscle actin. AMPK DN α1 and DN α2-transduced VSMCs displayed higher p-MLC expression compared to VSMCs transduced with the adenovirus-GFP control (Fig. 4F). These results suggested that quercetin affects vascular contraction by regulating MLCK expression and MLC phosphorylation through the LKB1-AMPK signaling pathway.

The regulation of quercetin through the LKB1-AMPK signaling pathway on expression of MLCK and p-MLC in VSMCs.
Discussion
The vascular tone, which plays an important role in the regulation of blood pressure and blood flow, 21 is known to be influenced by the contraction status of VSMCs in blood vessel walls. 22 The activation of MLCK by increased Ca2+-calmodulin complex concentration can regulate vasoactivity and blood flow. 23 During exercise and contraction, ATP is consumed in muscle, which in turn induces AMPK activation. 24 Then, AMPK activation inhibits the ATP-consuming pathways and activates metabolism of carbohydrate and fatty acid to restore ATP levels. 25 Furthermore, AMPK activation not only prevents vascular dysfunction 26 but also is involved in the regulation of vascular tone and remodeling. 27,28
It is clearly established that quercetin has many physiological effects, such as antioxidant and antihypertensive effects, and relates to improved vascular reactivity. 29,30 Previous studies demonstrated that quercetin induces the vasorelaxation and decreases the vascular resistance of arteries and potentially reduces blood pressure. 31,32 In addition, quercetin protects against diabetes-induced vascular contraction in rats. 33 Based on these data, we hypothesized that quercetin can induce the activation of AMPK signaling and that this can affect the vascular tone in aorta.
Similar to previous findings, our study indicated that quercetin affected PE-induced vascular contraction by inducing decreased expression of MLCK and p-MLC in rat aorta. This is the first study to report that vasorelaxation is affected by quercetin-induced LKB1-AMPK activation pathway in VSMCs.
In human leukemia cells, AMPK activation by quercetin reportedly induces apoptosis through an LKB1-independent pathway. 34 In contrast, our results showed that effects of quercetin in VSMCs were mediated by the LKB1-AMPK signaling pathway. Consistently, quercetin has been shown to activate LKB1 in bone marrow–derived macrophages, leading to the successive activation of both AMPK and ACC. 35
It is well established that quercetin interacts with Ca2+-calmodulin complexes in VSMCs, 36 but direct regulation of the Ca2+ concentration by quercetin has not been reported. According to the time course of AMPK activation in Figure 1, it can be assumed that quercetin-induced AMPK activation was influenced by upstream kinase LKB1. LKB1 phosphorylation precedes AMPK phosphorylation, whereas CaMKKβ expression increment is observed later than AMPK phosphorylation.
One of the most significant conclusions of our study is that LKB1-AMPK activation by quercetin regulates the expression of MLCK and p-MLC during VSMC contraction and was supported by several findings. First, quercetin regulated PE-induced vasocontraction through AMPK activation. Second, activation of the LKB1-AMPK signaling pathway by quercetin affected the expression of MLCK and p-MLC. Finally, the inhibition of LKB1-AMPK pathway restored the PE-induced increases in the expression of MLCK and p-MLC. Our results confirmed that quercetin directly affects the AMPK activation in VSMCs, resulting in enhanced vasorelaxation of aorta.
In conclusion, quercetin inhibits the PE-induced vascular contraction through the LKB1-AMPK signaling pathway, by decreasing the expressions of MLCK and p-MLC in VSMCs. Our data also support existing evidence that suggests a preventive or therapeutic role of quercetin against cardiovascular disorders such as atherosclerosis and hypertension.
Footnotes
Acknowledgments
This work was supported by the Medical Research Center Program (2015R1A5A2009124) through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning.
Author Disclosure Statement
No competing financial interests exist.