
Abstract
Korean red ginseng (KRG), a heat-processed Korean ginseng (Panax ginseng C.A. Meyer), has been used as a traditional medicine for its beneficial effects on hyperglycemia. This study aimed to investigate whether the antidiabetic action of KRG in an animal model of type 2 diabetes mellitus (DM) is partly mediated by prevention of mitochondrial dysfunction and intracellular inflammation. Four-week-old C57BL/KsJ db/db mice (a genetic animal model of obese type 2 DM) and C57BL/KsJ db/+ mice were divided into three groups: db/+ mice (normoglycemic control group, n = 8), db/db mice (untreated DM group, n = 8), and db/db mice with KRG administration (KRG-treated DM group, n = 8). After 12 weeks, metabolic parameters of fasting blood glucose concentrations, hemoglobin A1c (HbA1c) level, insulin level, lipid profile, and leukocyte count were determined using high-performance liquid chromatography. Mitochondrial DNA (mtDNA) copy number and inflammatory marker (interleukin-6, cyclooxygenase-2, and C-reactive protein) expression levels were measured in skeletal muscle tissue using quantitative real-time PCR analysis. After 12 weeks of KRG treatment at 100 mg/kg, the fasting glucose, HbA1c, insulin, and low-density lipoprotein cholesterol concentrations were lower, whereas mtDNA copy numbers were higher in the KRG-treated DM group than in the untreated DM group. Compared with the untreated DM group, the messenger RNA expression levels of mitochondrial biogenesis-related transcription factors and inflammatory markers were lower in the KRG-treated DM group. In conclusion, KRG had a beneficial effect on the metabolic profile by preserving mitochondrial function and protecting against intracellular inflammation.
Introduction
M
Mitochondrial dysfunction includes mitochondrial loss and overproduction of ROS, such as superoxides and other free radicals. Dysregulation of mitochondrial DNA (mtDNA) copy number and decreased mtDNA copy number have been found to be associated with type 2 DM. 3 –5 Furthermore, in type 2 DM, hyperglycemia induces overproduction of ROS and impairs the mitochondrial antioxidant defense mechanism. 6,7 Thus, mitochondrial dysfunction, oxidative stress, and chronic low-grade inflammation are closely interrelated with the pathogenesis of type 2 DM.
Ginseng, the root of Panax ginseng C.A Meyer, has been widely used as a botanical medicine in East Asia for >2000 years. Korean red ginseng (KRG) is produced by steaming raw ginseng root under high pressure and temperature. KRG contains a variety of bioactive compounds, including phenolic acid, flavonoids, and ginsenosides, all of which have been used for multiple pharmacological purposes such as treating cardiovascular and metabolic diseases. 8,9 KRG has been reported to exert antidiabetic activity in in vitro and in vivo studies, including studies of patients with type 2 DM 10,11 and animal models of type 2 DM. 12 Although the biological mechanisms by which KRG exerts its antidiabetic effects are not fully understood, most studies have focused on studying how KRG improves the insulin-secreting action of pancreatic β cells and glucose disposal of peripheral tissues. 13,14
The development and progression of type 2 DM are also mediated by mitochondrial dysfunction and chronic low-grade inflammation. Moreover, KRG is predicted to modulate mitochondrial function and intracellular inflammation levels. Thus, this study aimed to investigate whether the antidiabetic effects of KRG are also mediated through the prevention of mitochondrial dysfunction and intracellular inflammation in an animal model of type 2 DM.
Materials and Methods
Animals and experimental design
The research was conducted in accordance with the internationally accepted principles for laboratory animal use and care as found in the U.S. guidelines. Moreover, the study protocol and all related procedures were approved by the Institutional Animal Care and Use Committee at Yonsei Institute for Life Sciences, Yonsei University, Seoul, Korea (Approval No.: 2011-0291).
We adopted the C57BL/KsJ db/db mouse model for type 2 DM, which has an autosomal recessive mutation in the leptin receptor, ultimately leading to obesity, hyperinsulinemia, and hyperglycemia. 15 Previous studies have used C57BLKsJ-db/db mice as a hyperglycemic type 2 DM animal model and C57BLKsJ-db/+ mice as the normoglycemic control group. 16 Four-week-old C57BL/KsJ db/db and C57BL/KsJ db/+ male mice were purchased from Joongang Lab (Seoul, Republic of Korea). All animals were acclimatized to the laboratory environment for 2 weeks before the initiation of experiments. After acclimatization, the mice were divided into three groups: db/+ mice (normoglycemic control group, n = 8), db/db mice (untreated DM group, n = 8), and db/db mice that were administered KRG for 12 weeks (KRG-treated DM group, n = 8). Mice were allowed free access to drinking water and a chow diet under constant room temperature (22 ± 2°C) and humidity (50 ± 10%) with an automatic 12-h light and 12-h dark cycle. Food intake and body weight were recorded every day, and glucose concentration was recorded weekly for 12 weeks.
KRG powder was provided in dried form from the Korean Ginseng Cooperation (Daejeon, Korea). Our preliminary experiments revealed that the antihyperglycemic effect of KRG was dose dependent (data not shown); 100 mg/kg was subsequently chosen as the daily dose of KRG. KRG powder was dissolved in 1 cc of distilled water and injected once daily at a dose of 100 mg/kg body weight (∼0.2 mL per injection) into the intraperitoneal cavity of animals in the KRG-treated DM group. Animals in the untreated DM group received an equivalent saline injection, whereas animals in the normoglycemic control group did not receive any injection or treatment.
Biochemical analysis and tissue sampling
Body weight was measured at 11:00 AM. Fasting blood glucose level was measured by collecting blood samples from the tail vein on day 0 (before treatment) and then again every week thereafter (days 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, and 84). All samples were obtained after animals had been fasted for 4 h, starting at 7:00 AM. Glucose concentrations were determined in blood samples collected from the tail vein at 11:00 AM using a Glucose Analyzer (HemoCue AB, Angelholm, Sweden). After 12 weeks, all 24 mice were sacrificed under light ether anesthesia. Blood samples for the analysis of fasting plasma glucose, hemoglobin A1c (HbA1c), insulin, total cholesterol, triglyceride, high-density lipoprotein cholesterol, and low-density lipoprotein (LDL) cholesterol concentrations, and leukocyte counts were obtained by exsanguination of the heart. Parameters were measured using high-performance liquid chromatography (SIMADZU, Kyoto, Japan). Gastrocnemius muscle tissue samples were collected, quickly frozen in liquid nitrogen, and kept at −80°C until analysis.
mtDNA analysis
The mtDNA copy numbers were measured from gastrocnemius muscle tissue using quantitative real-time PCR analysis. Briefly, total DNA was extracted with a QIAamp DNA Extraction Kit (QIAGEN, Hilden, Germany). The ratio of mtDNA to nuclear DNA, which reflects the concentration of mtDNA in each tissue, was subsequently determined. Targeted genes were nuclear cystic fibrosis and mitochondrial nicotinamide adenine dinucleotide dehydrogenase-5 (ND5). For nuclear DNA quantification, 10 ng DNA was used as a template. Mouse-specific primers (sequences reported in Table 1) were selected using Primer Express Software (Biosystems, Barcelona, Spain).
CF, cystic fibrosis; COX-2, cyclooxygenase-2; CRP, C-reactive protein; IL-6 interleukin-6; ND5, nicotinamide adenine dinucleotide dehydrogenase-5; PGC1α, peroxisomal proliferator-activated receptor-γ coactivator-1α; T-fam, mitochondrial transcription factor A.
Quantitative real-time PCR analysis
Total RNA was isolated from liver tissue using TRIzol (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instruction. From each sample, 3 μg total RNA was reverse transcribed into complementary DNA using a Reverse Transcription System Kit (Thermo Fisher, Middlesex County, MA, USA). Briefly, quantitative RT-PCRs were performed with Power SYBR Green Master Mix (Biosystems) as described in the manufacturer's instructions. Thermo cycling was performed using an ABI 7500 Real-Time PCR system (Biosystems) with the following cycling parameters: stage 1, 50°C for 2 min; stage 2, 95°C for 10 min; stage 3, 40 cycles at 95°C for 15 sec; 60°C for 1 min; and stage 4, 95°C for 15 sec, 60°C for 15 sec, and 95°C for 15 sec. Primers were designed using Primer Express Software (Biosystems) (Table 1). Mouse target genes consisted of peroxisomal proliferator-activated receptor-γ coactivator-1α (PGC1α) and mitochondrial transcription factor A (T-fam) for markers of mitochondrial biogenesis and the inflammatory markers interleukin-6 (IL-6), cyclooxygenase-2 (COX-2), and C-reactive protein (CRP) (Table 1). The linearity of the dissociation curves was analyzed using ABI 7500 software. Data were analyzed by the comparative method (2−ΔΔCt) using an internal control. Each sample was analyzed in duplicate.
Statistical analysis
Results are presented as mean ± standard deviation. Data were compared using the analysis of variance test and Student's t-test as a post hoc analysis. All analyses were conducted using SAS statistical software, version 9.2 (SAS Institute, Inc., Cary, NC, USA). All statistical tests were two sided, and the threshold for statistical significance was set at P-value <0.05.
Results
During daily administration for 12 weeks, no skin irritation or any other adverse effect was observed. In our study, food intake was significantly higher in DM groups than in normoglycemic control group (12.9 g/day), but did not differ between untreated db/db group (18.4 g/day) and KRG-treated db/db group (17.9 g/day). There were no significant differences in body weight between the untreated DM group and the KRG-treated DM group throughout the duration of the experiment (Appendix 1).
Figure 1 shows the changes in fasting blood glucose concentrations during the 12 weeks. At the initial stage, blood glucose levels were 78.8 mg/dL for normoglycemic control group, 145.8 mg/dL for untreated DM group, and 172.3 mg/dL for KRG-treated DM group. However, after 1 week (at 5 weeks of age), the blood glucose concentrations increased steeply in the DM group: 192.0 mg/dL for normoglycemic control group versus 457.0 mg/dL for untreated DM group and 458.6 mg/dL for KRG-treated DM group, respectively. The fasting blood glucose concentrations of the untreated DM group and the KRG-treated DM group were not significantly different until 8 weeks. However, the KRG-treated DM group had a significantly lower mean fasting blood glucose concentration than the untreated DM group after 9 weeks. Table 2 presents the changes of metabolic parameters according to KRG treatment. Overall, a 12-week administration of KRG resulted in favorable effects on the metabolic profile. After 12 weeks of KRG treatment at 100 mg/kg, the fasting blood glucose, HbA1c, insulin, LDL cholesterol concentrations, and leukocyte counts were significantly lower in the KRG-treated DM group compared with the untreated DM group (all P < 0.05).
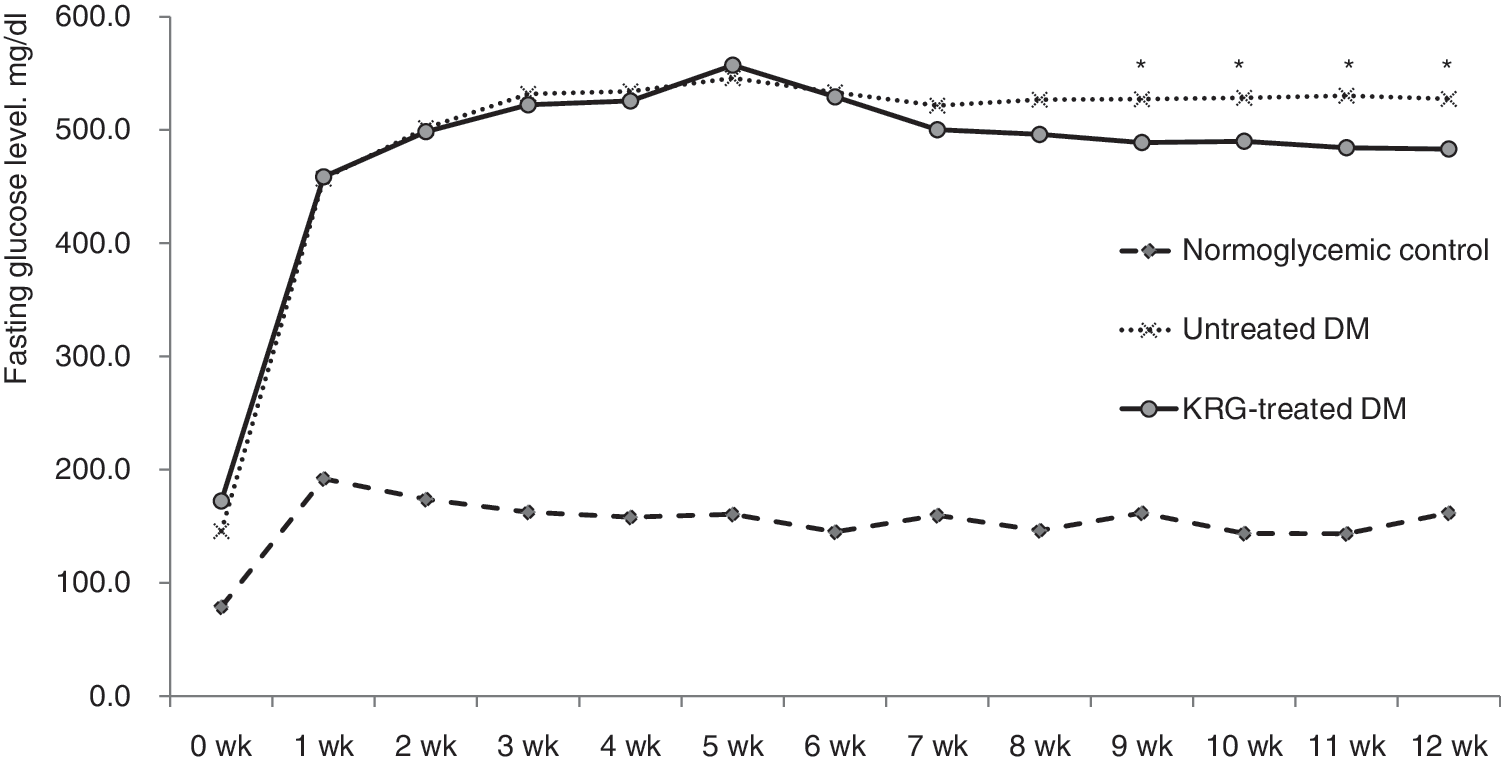
The changes in fasting blood glucose levels during 12 weeks. *P-value <0.05 vs. the untreated DM group. DM, diabetes mellitus; KRG, Korean red ginseng.
P-value <0.05 vs. normoglycemic control group.
P-value <0.05 vs. untreated DM group.
DM, diabetes mellitus; HbA1c, hemoglobin A1c; HDL, high-density lipoprotein; KRG, Korean red ginseng; LDL, low-density lipoprotein.
Compared with the untreated DM group, the mtDNA copy numbers and messenger RNA (mRNA) expression levels of mitochondrial biogenesis-related transcription factors (PGC1α and T-fam) were significantly higher in the KRG-treated DM group (Figs. 2 and 3). Moreover, the mRNA expression levels of inflammatory markers (IL-6, COX-2, and CRP) were significantly lower in the KRG-treated DM group (Fig. 4).
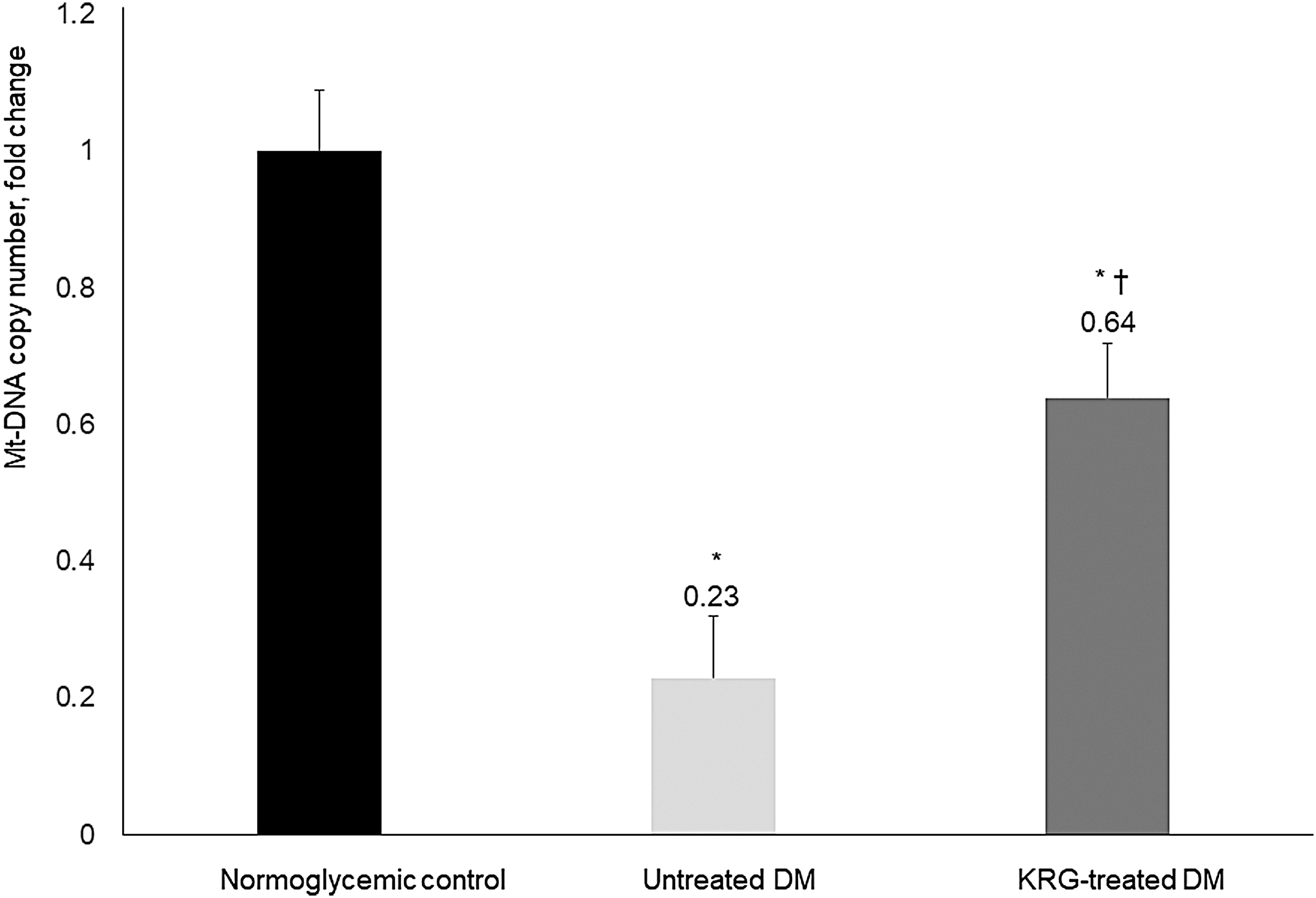
Effects of KRG on mtDNA copy number. Data are presented as mean ± SD. *P-value <0.05 vs. the normoglycemic control group, † P-value <0.05 vs. the untreated DM group. mtDNA, mitochondrial DNA; SD, standard deviation.
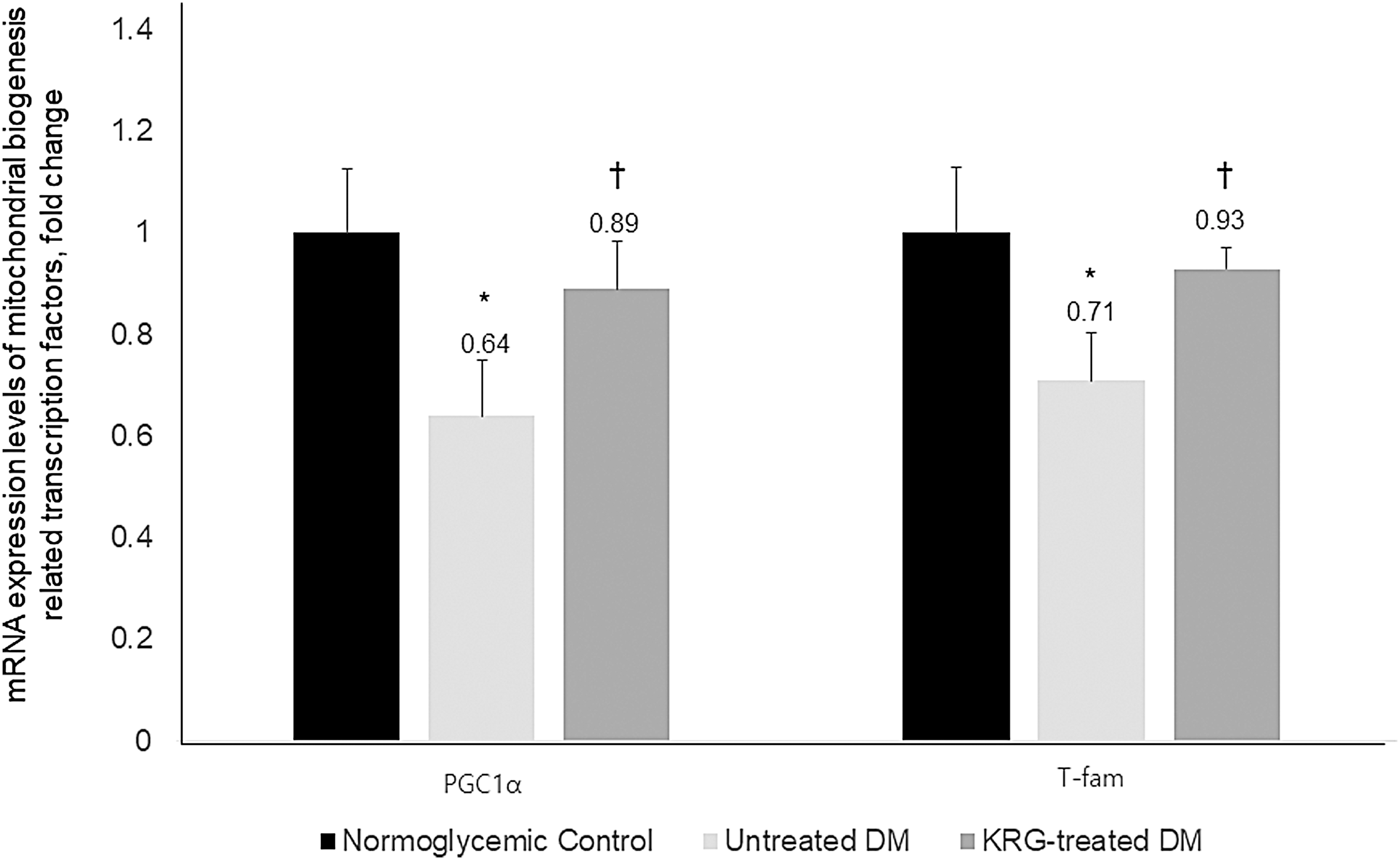
Effects of KRG on the mRNA expression levels of mitochondrial biogenesis-related transcription factors. Data are expressed as mean ± SD. *P-value <0.05 vs. the normoglycemic control group, † P-value <0.05 vs. the untreated DM group. mRNA, messenger RNA; PGC1α, peroxisomal proliferator-activated receptor-γ coactivator-1α; T-fam, mitochondrial transcription factor A.
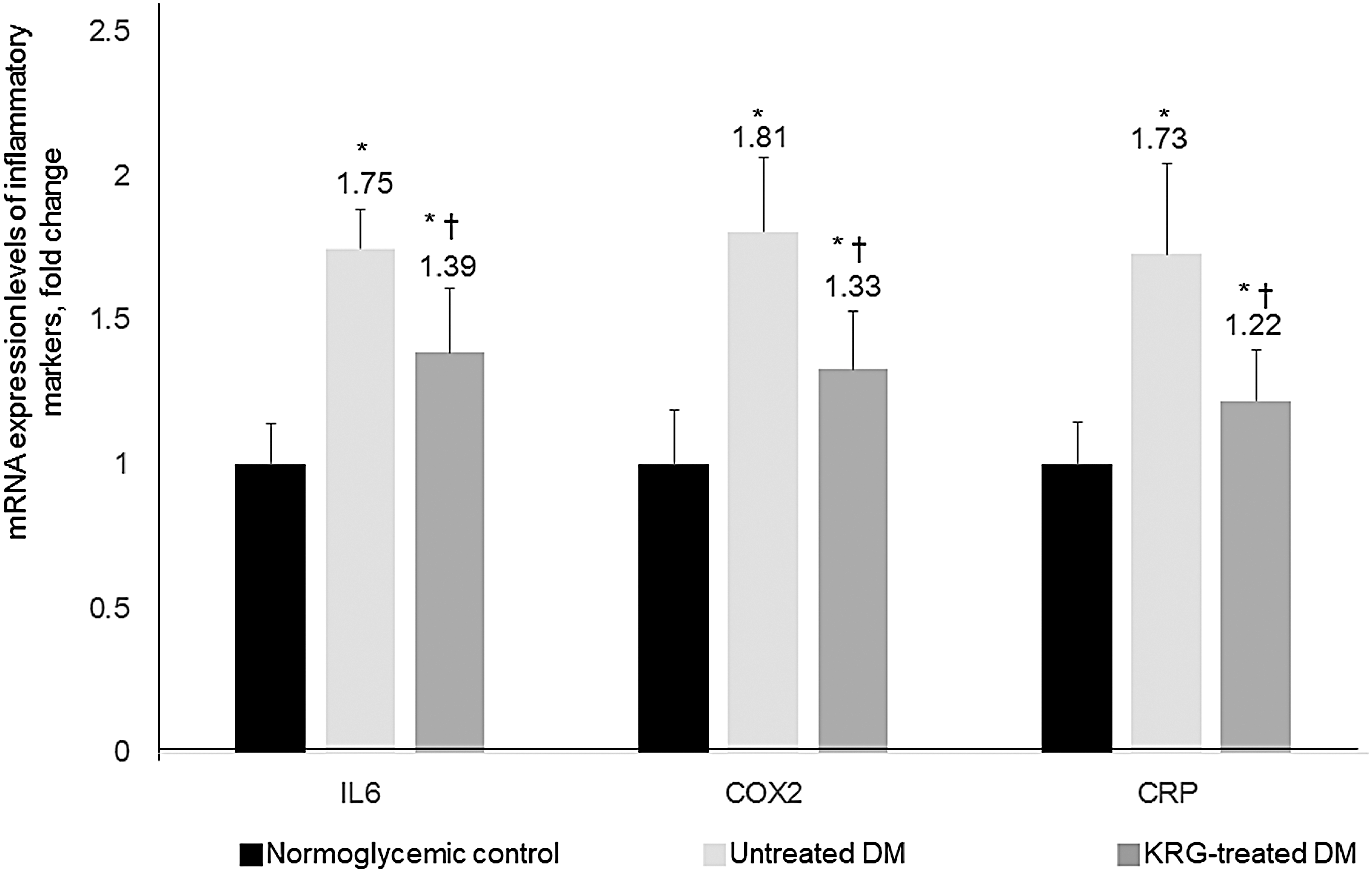
Effects of KRG on the mRNA expression levels of inflammatory markers. Data are expressed as mean ± SD. *P-value <0.05 vs. the normoglycemic control group, † P-value <0.05 vs. the untreated DM group. COX-2, cyclooxygenase-2; CRP, C-reactive protein; IL-6 interleukin-6.
Discussion
Using an animal model of type 2 DM, we tested the hypothesis that KRG has a beneficial effect on the metabolic profile by preserving mitochondrial function and protecting against intracellular inflammation. A 12-week course of KRG administration improved fasting blood glucose, HbA1c, insulin, and LDL cholesterol concentrations. We believe that this is the first study to determine that these favorable effects of KRG on the metabolic profile might be partly mediated through protection against mitochondrial damage and intracellular inflammation. Furthermore, it has been demonstrated that ginseng has an antihyperglycemic effect in diabetic animal models, but most of the results have shown only short-term effects. 17,18 In the present study, we demonstrated the long-term favorable metabolic effects.
Our results are in agreement with previous findings. Several clinical trials have documented that ginseng ameliorates hyperglycemia and elevated HbA1c in patients with type 2 DM. 10,11 Moreover, ginseng has been reported to attenuate the development of type 2 DM in animal models. 12 Although the exact mechanisms by which KRG exerts its hypoglycemic effects have not been elucidated, previous studies have suggested that KRG might be involved in stimulating insulin secretion and improving insulin resistance. For instance, KRG has been reported to stimulate glucose-induced insulin secretion from pancreatic β cells. 13 KRG has many biochemical constituents, of which ginsenosides are the main pharmacologically active components. 19 Since the biological activities of ginseng are enhanced by heat processing, 20 heat-processed KRG has been reported to have more potent pharmacological activity than nonprocessed ginseng. In the process of red ginseng production, major ginsenosides such as Rb1, Rb2, Rc, Rd, Rg1, and Re in fresh ginseng are denatured and converted into newly transformed ginsenosides with high concentrations. 21 Moreover, new bioactive compounds such as arginine–fructose–glucose and acidic polysaccharide can be produced by chemical reaction during red ginseng production. 8
A decrease in mtDNA copy number has been related to the development of type 2 DM. Moreover, a number of studies have reported downregulation of mitochondrial gene expression and lower protein expression of mitochondrial respiratory chain subunits in type 2 DM. 21 In the present study, the mRNA expression levels of PGC1α and T-fam were significantly higher in the KRG-treated DM group compared with the untreated DM group, suggesting KRG administration also enhanced PGC-1α and T-fam expression in the skeletal muscles of db/db mice. Interestingly, Santos et al. 22 reported that hyperglycemic insult stimulates mtDNA repair and biogenesis, thereby protecting the electron transport chain system and preventing the initiation of the vicious ROS cycle. In the present study, the mtDNA copy numbers were significantly higher in the KRG-treated DM group than in the untreated DM group, which suggests that KRG has a beneficial effect on type 2 DM by preserving or improving mitochondrial function. We used mtDNA copy number in skeletal muscle as a measurement of mitochondrial function. As one of the most common biomarkers for mitochondrial dysfunction, mtDNA copy number has been widely used in clinical and experimental studies. 23 The mtDNA copy number, which reflects the mtDNA content, is associated with the stability of mitochondrial genes and mitochondrial biogenesis. 24 Although mtDNA copy number can vary among organs, changes in mtDNA levels in muscle have been reported to reflect comparable processes in other tissues, such as blood leukocytes and hepatocytes. 25
Another noteworthy finding of the present study is that the levels of intracellular inflammatory markers were significantly decreased. A mounting body of evidence suggests that mitochondrial dysfunction induced by oxidative stress and chronic low-grade inflammation plays a key role in the development of insulin resistance and the pathogenesis of type 2 DM. Indeed, oxidative stress in the diabetic state can be induced by an increase in free radical production and a disturbance in free radical scavenging systems. In vitro and in vivo studies have suggested that KRG decreases oxidative stress and increases antioxidant enzyme activity. 26,27 Oxidative stress markers and inflammatory markers such as tumor necrosis factor-α, IL-6, CRP, and leukocyte count are elevated in patients with type 2 DM and insulin resistance. 23 Moreover, Lee et al. 28 reported that KRG exerted anti-inflammatory effects in a rat model of aging with high-fat diet-induced atherosclerosis. In our animal study, the mRNA expression levels of IL-6, COX-2, and CRP were also significantly decreased in db/db mice after treatment with KRG. Our results suggest that the antioxidant constituents of KRG prevent free radical chain reactions and attenuate intracellular inflammation, at least in an animal model of type 2 DM.
In summary, KRG administration improved blood glucose concentrations and insulin sensitivity, and these effects were associated with protection from mitochondrial damage and reduction of the inflammatory response. Our findings indicate that KRG is a potentially useful additive nutraceutical in type 2 DM.
Footnotes
Acknowledgment
This study was supported by a faculty research grant from Yonsei University College of Medicine in 2009 (Grant No. 6-2009-0097).
Authors' Contributions
J.K.P. analyzed and interpreted the data and wrote the article. J.Y.S., A.R.C., and M.R.C. assisted with data analysis and interpretation and revised the article. Y.J.L. designed the study, performed the experiment, analyzed and interpreted data, and revised the article. All authors read and approved the final article.
Author Disclosure Statement
The authors declare no potential conflicts of interests with respect to the research, authorship, and/or publication of this article.