
Abstract
Red pepper seed, a by-product of red pepper, has been reported to have antioxidant and antiobesity activities. However, its role in diabetes has not yet been highly investigated. Glucose homeostasis is mainly maintained by insulin, which suppresses glucose production in the liver and enhances glucose uptake in peripheral tissues. In this study, we investigated the underlying mechanisms through which red pepper seed extract (RPSE) affects glucose production in AML12 hepatocytes and glucose uptake in C2C12 myotubes. RPSE reduced glucose production in a dose-dependent manner in AML12 cells. The levels of glucose 6 phosphatase, phosphoenolpyruvate carboxykinase, and critical enzymes for hepatic gluconeogenesis were decreased by RPSE. Gluconeogenesis regulating proteins, Akt and forkhead box protein O1, were also activated by RPSE. In addition, RPSE increased glucose uptake in C2C12 via inducing translocation of glucose transporter type 4 from cytosol to plasma membrane. Analysis of the insulin-dependent pathway showed that the activities of insulin receptor substrate 1, phosphatidylinositol 3-kinase, and Akt were significantly stimulated by RPSE. In conclusion, RPSE might improve glucose homeostasis by reducing hepatic gluconeogenesis and increasing peripheral glucose uptake. Results obtained also suggest that RPSE can be a compelling antidiabetic nutraceutical.
Introduction
T
In treatment of diabetes mellitus, suppressing glucose production by the liver and enhancing glucose uptake in muscle is essential processes for therapeutic remedies. There are several glucose lowering drugs such as thiazolidinediones (pioglitazone and rosiglitazone), sulfonylureas, and metformin for the treatment of diabetes. 8 Despite their effectiveness, these drugs have side effects which vary in their extents. Thiazolidinediones and sulfonylureas are associated with hypoglycemia and weight gain, 9 –11 yet metformin can induce lactic acidosis 12 . Due to the noted and reported side effects, the demand for new antidiabetic substances is continuously increasing.
In an effort to discover advantages of red pepper seed, several studies have been conducted to reveal its physiological effects. 13,14 Red pepper (Capsicum annuum L.) is widely used in food and its physiological functions have been reported in several studies. 15,16 Apart from the skin and core, red pepper seed is seldom utilized for food material. Red pepper seed extract (RPSE) reduced mutagenicity induced by 2-nitrofluorene, sodium azide, and 2-aminoanthracene and increased nitric oxide and 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) scavenging activities, suggesting its antimutagenic and antioxidant effects. 15 In addition, RPSE was found to inhibit lipid accumulation via lowering expression of CCAAT-enhancer-binding protein alpha (C/EBPα), C/EBPβ, Peroxisome proliferator-activated receptor gamma (PPARγ), and adipogenic transcription factors in 3T3-L1 adipocyte. 16 Nonetheless, the antidiabetic activity of RPSE remains to be elucidated. This article presents research on the hypoglycemic effects of RPSE in vitro as well as an evaluation of its effects on the hepatic glucose production and muscle glucose uptake.
Materials and Methods
Preparation of RPSE
RPSE was obtained from NOVAREX, Inc. (Seoul, Republic of Korea). The company purchased the red pepper seed cultivated in Yeong-yang from Yeong Yang Red Pepper Trade Corp. (Republic of Korea). The red pepper seed was blended with water (solid:water, 1:9) at 50°C for 18–24 h followed by filtration of the extracts (90 mm diameter, 5 μm pore size) (Hyundai Micro Co., Ltd, Seoul, Korea). The filtered extracts were concentrated using a vacuum evaporator at 40–50°C and then spray-dried at 180–210°C by spray dryer (MSD-60-N; Niro Korea, Inc., Cheonan, Republic of Korea).
Cell line and culture
AML12 hepatocytes (American Type Culture Collection, Rockville, MD, USA) were cultured in DMEM/F12 supplemented with 10% fetal bovine serum (FBS), 0.005 mg/mL insulin, 0.005 mg/mL transferrin, 5 ng/mL selenium, 40 ng/mL dexamethasone, and 1 × penicillin streptomycin. C2C12 myocytes (American Type Culture Collection) were cultured in DMEM supplemented with 10% FBS and 1 × penicillin streptomycin. For differentiation into myotubes, myocytes were cultured in DMEM supplemented with 2% horse serum for 3–5 days after reaching 100% confluence. Both AML12 and C2C12 cells were maintained in an incubator at 37°C with humidified air and CO2 (5%).
MTT assay
The effect of RPSE on cell viability was measured using the MTT assay. AML12 hepatocytes were plated into 96-well plates at a density of 1.0 × 104/well. After 70% confluence, cells were starved for 18 h in serum-free media and then treated with RPSE at concentrations of 50, 100, 150, and 200 μg/mL. Differentiated C2C12 myotubes were also starved and treated as described above. After 48 h of incubation, 5 mg/mL of MTT reagent in media was administered for an additional 2 h and the formazan was dissolved in DMSO for 10 min. Absorbance of dissolved formazan was determined by use of a spectrophotometer (BioTek Instruments, Inc., VT, USA) at 540 nm.
Glucose production assay in hepatocytes
AML12 hepatocytes were seeded at 1.0 × 104/well in 96-well plates and after the confluency of cells was 70–80%, cells were treated with RPSE at concentrations of 50, 100, 150, and 200 μg/mL in serum-free media for 48 h. Cells were washed twice with Dulbecco's phosphate-buffered saline (DPBS) to remove glucose and then incubated in glucose and phenol red-free DMEM containing 2 mM sodium pyruvate and 20 mM sodium lactate for 3 h. The glucose concentrations of media were measured using an Amplex Red Glucose/Glucose Oxidase Assay Kit (Invitrogen, CA, USA). The media were reacted with reagents in kit, and absorbance was measured at 585 nm using spectrophotometer (BioTek Instruments, Inc.).
Glucose uptake assay in myotubes
C2C12 myocytes were cultured in black, clear bottom 96-well plates. After differentiated into myotubes, cells were starved for 18 h in serum-free media. Cells were treated with RPSE at concentrations of 50, 100, 150, and 200 μg/mL and 30 μM 2-deoxy-2-[(7-nitro-2,1,3-benzoxadiazol-4-yl) amino]-D-glucose (2-NBDG), fluorescently labeled deoxyglucose analog in glucose-free media without or with 100 nM insulin for 48 h. At the end of incubation, cells were washed twice with ice-cold DPBS to remove unabsorbed 2-NBDG. The 2-NBDG taken up by cells was detected by a spectrophotometer (BioTek Instruments, Inc.) designed to detect fluorescein (excitation/emission = 485/528 nm).
Cell membrane protein isolation
For detection of GLUT4 translocation, the plasma membrane was isolated from cell lysates by using a pierce cell surface protein isolation kit (Thermo Scientific, MA, USA). Cells were labeled with a thiol-cleavable amine-reactive biotinylation reagent (Thermo Scientific™ EZ-Link™ Sulfo-NHS-SS-Biotin) followed by a quench reaction. Cells were harvested and lysed with lysis buffer containing protease inhibitor. The labeled proteins were isolated from cell lysates by NeutrAvidin™ Agarose using column and eluted with SDS-PAGE buffer containing 50 mM DTT.
Immunoblotting
For the evaluation of glucose production-related proteins, AML12 were treated with RPSE at concentrations of 50, 100, 150, and 200 μg/mL in serum-free media for 48 h. To verify the insulin signaling activity by RPSE, C2C12 myotubes were treated with RPSE in differentiation media for 48 h and then further treated with 100 nM insulin for 15 min. After treatment, harvested AML12 and C2C12 cells were lysed with RIPA buffer containing protease inhibitor and phosphatase inhibitor (Cell Signaling Technology, Inc., MA, USA). Total protein was measured by Bradford assay, 17 using bovine serum albumin (BSA) as a standard. The proteins were separated on 20% SDS-PAGE, followed by electrophoretic transfer onto PVDF membranes. The PVDF membranes were blocked with 5% BSA in TBST for 50 min. Primary antibodies were applied to the blot overnight and washed with TBST for 10 min three times. The blot was incubated with appropriate HRP-conjugated secondary antibodies (Bio-Rad Laboratories, Inc., CA, USA) for 1 h and washed with TBST for 15 min four times. The blot was quantified with Bio-Rad Image Lab (ChemiDoc; Bio-Rad Laboratories, Inc.). β-actin and Na+/K+-ATPase α were used as the reference control. Antibodies against G6Pase, PEPCK, and Na+/K+-ATPase α were from Santa Cruz Biotechnology (TX, USA). Antibody against IRS-pY612 was from Abcam (Cambridge, United Kingdom). Antibodies against Akt, Akt-p473, Akt-pT308, forkhead box protein O1 (FoxO1), FoxO1-pS256, IRS1, phosphatidylinositol 3-kinase (PI3K), PI3K-pY199, GLUT4, and β-actin were from Cell Signaling Technology, Inc.
Statistical analysis
All data are presented as the mean ± standard error of three independent experiments. Statistical analyses were performed using the PRISM 6 program (GraphPad Software, Inc., CA, USA). Data were analyzed by one-way ANOVA followed by Dunnett's multiple comparisons test with statistical significance defined as P < .05.
Results
Cell viability
Cells were exposed to RPSE at concentrations of 50, 100, 150, and 200 μg/mL for 48 h before the MTT assay. Figure 1A and B show that treatment for 48 h did not change the cell viability in AML12 hepatocytes and C2C12 myotubes.
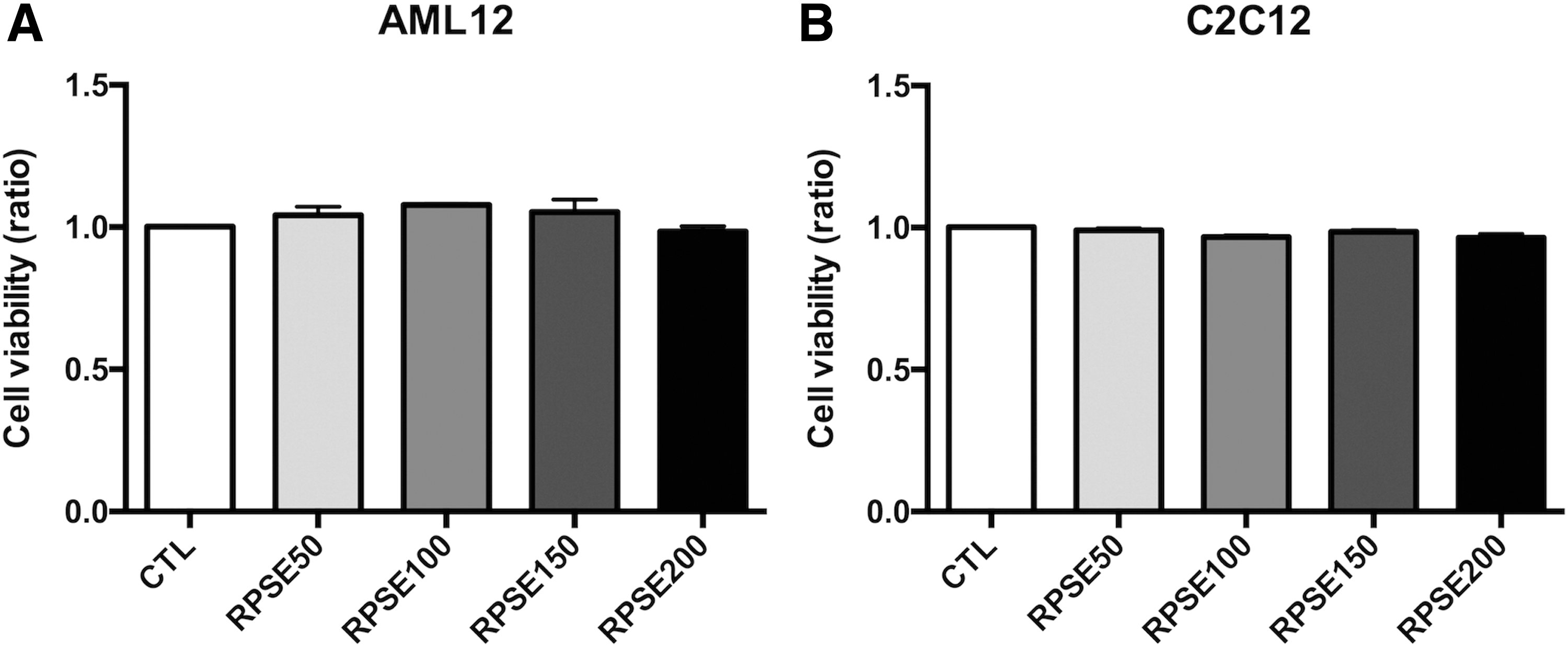
The effect of RPSE on cell viability in AML12 hepatocytes
Suppression of hepatic glucose production
Figure 2 shows that RPSE reduced glucose production in a concentration-dependent manner. The mean glucose concentration of controls was 30.86 ± 5.49 μM. After treatment with 50, 100, 150, and 200 μg/mL RPSE, glucose production was lowered by 8.83%, 25.22%, 24.45%, and 29.25%, respectively (P < .01), in comparison with the control. To examine the mechanism, Figure 3 shows the expressions of G6Pase and PEPCK and their relative intensity. The levels of G6Pase and PEPCK were lowered by incubation with RPSE, decreasing by about 41.50% and 19.63% (P < .001 and P < .01, respectively) compared with the control at a concentration of 200 μg/mL. Furthermore, as shown in Figure 4, the levels of Akt-pS473/Akt and FoxO1-pS256/FoxO1 were significantly upregulated by RPSE, increasing approximately threefold against the control at a concentration of 200 μg/mL (P < .001).
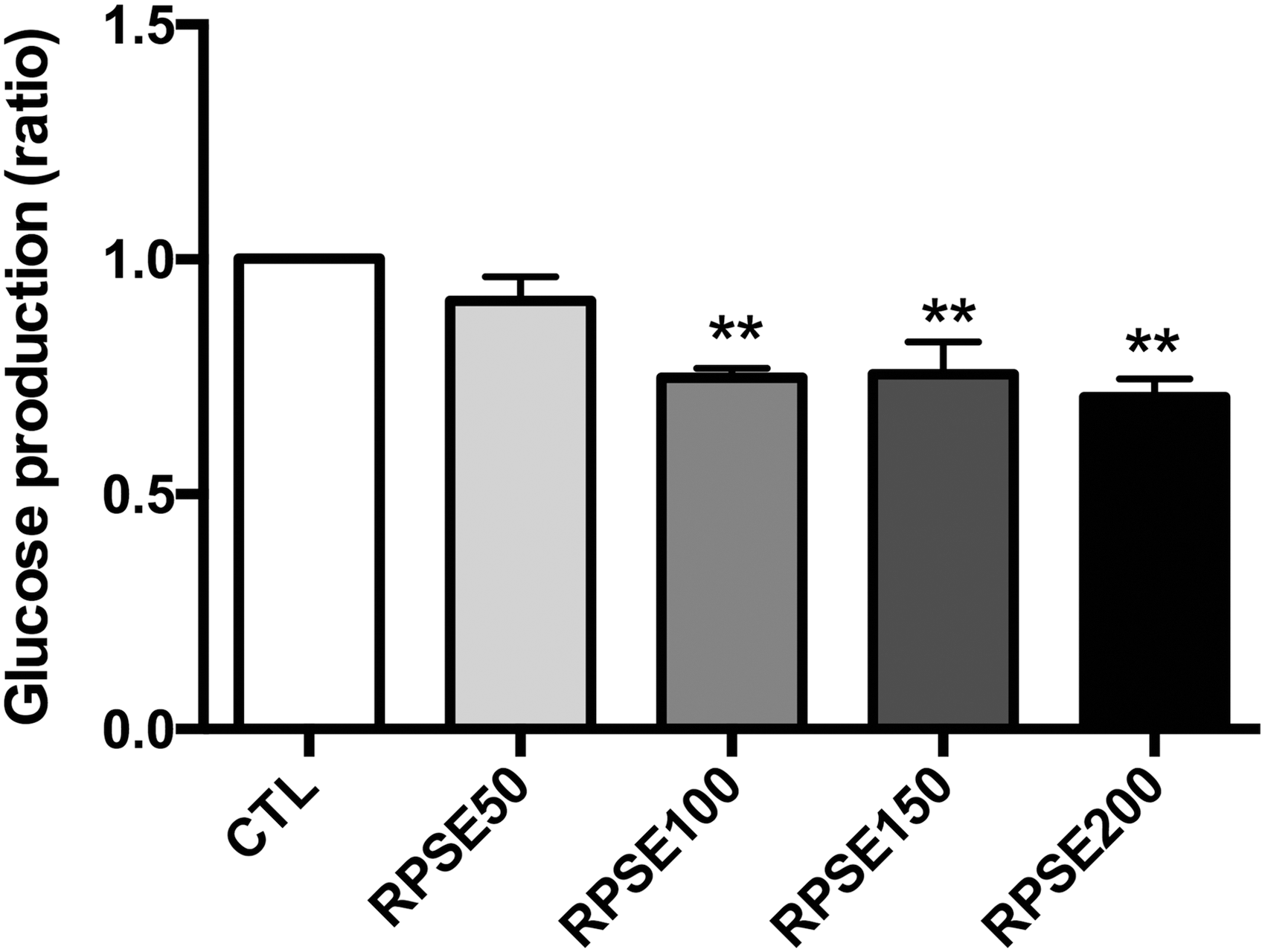
The effect of RPSE on gluconeogenesis in AML12 hepatocytes. AML12 hepatocytes were incubated for 48 h with RPSE in serum-free media supplemented with RPSE at concentrations of 50, 100, 150, and 200 μg/mL. Cells were further incubated for 3 h in glucose- and phenol red-free DMEM containing 2 mM sodium pyruvate and 20 mM sodium lactate. Values are expressed as means ± standard errors of three independent experiments. **P < .01 compared to CTL. CTL, control.
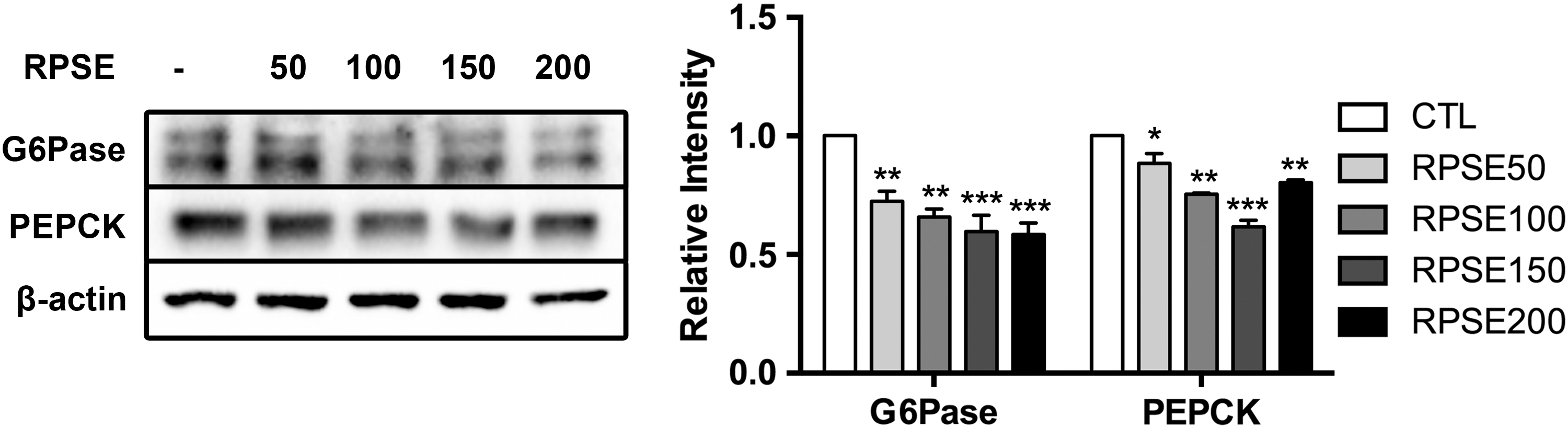
The effect of RPSE on G6Pase and PEPCK in AML12 hepatocytes. AML12 hepatocytes were incubated for 48 h in serum-free media supplemented with RPSE at concentrations of 50, 100, 150, and 200 μg/mL. The bands of representative experiments and relative intensity of G6Pase and PEPCK are presented with β-actin used as the reference control. Values are expressed as means ± standard errors of three independent experiments. *P < .05, **P < .01, ***P < .001 compared to CTL. G6Pase, glucose 6 phosphatase; PEPCK, phosphoenolpyruvate carboxykinase.

The effect of RPSE on Akt and FoxO1 in AML12 hepatocytes. AML12 hepatocytes were incubated for 48 h in serum-free media supplemented with RPSE at concentrations of 50, 100, 150, and 200 μg/mL. The bands of representative experiment and relative intensity of Akt-pS473/Akt and FoxO1-pS256/FoxO1 were presented with β-actin used as the reference control. Values are expressed as means ± standard errors of three independent experiments. *P < .05, ***P < .001 compared to CTL. Fox, forkhead box.
Enhancement of muscle glucose uptake
Figure 5 shows that RPSE increased 2-NBDG uptake. RPSE at a concentration of 150 and 200 μg/mL increased the glucose uptake by 32.71% and 48.71% compared with the control. It was also illustrated that the level of GLUT4 in plasma membranes was significantly enhanced by RPSE in Figure 6.
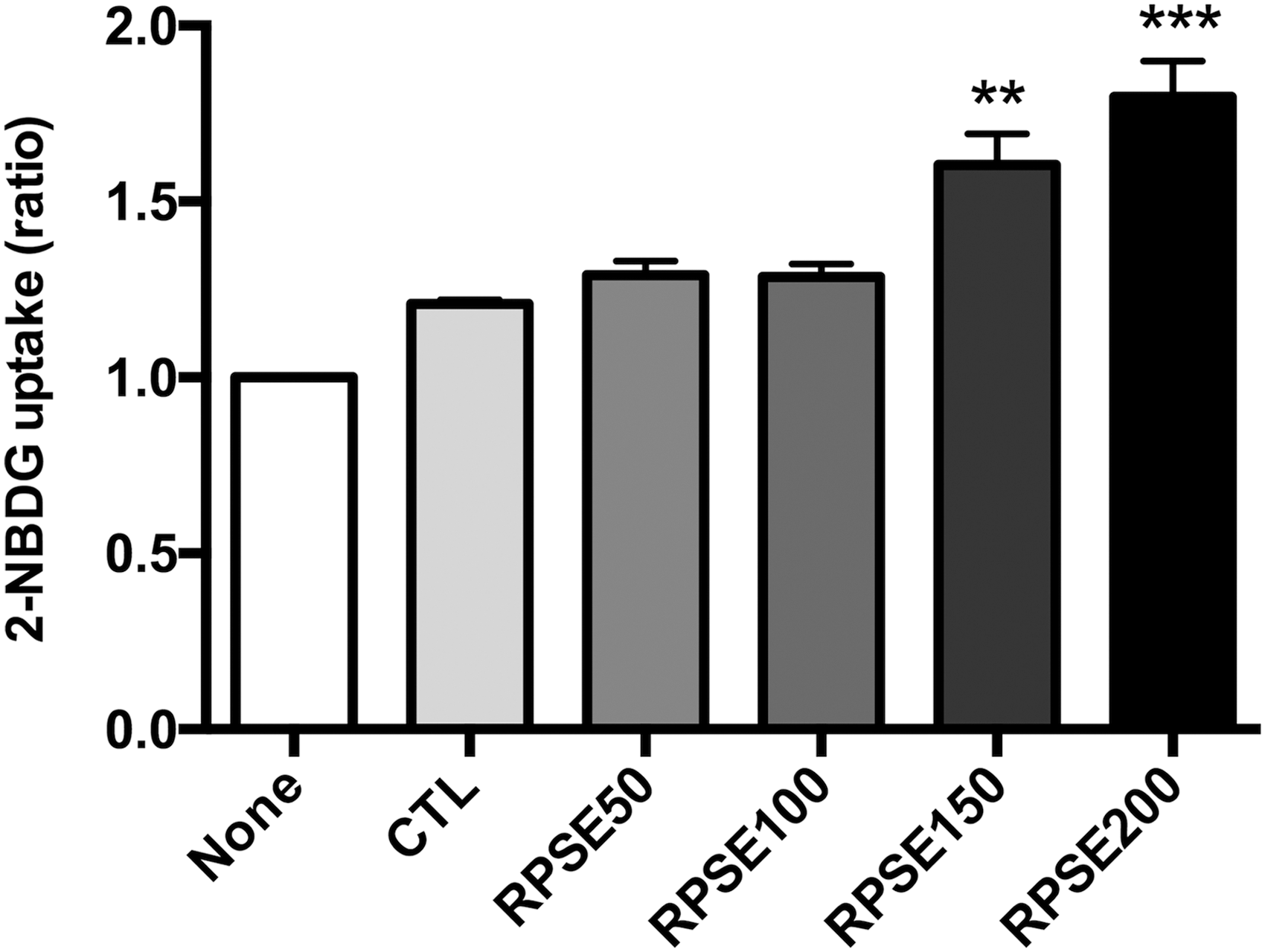
The effect of RPSE on glucose uptake in C2C12 myotubes. C2C12 myotubes were treated RPSE at concentrations of 50, 100, 150, and 200 μg/mL and 2-NBDG in glucose-free media without 100 nM insulin (None) or with 100 nM insulin (CTL, RPSE50, RPSE100, RPSE150, and RPSE200) for 48 h. Values are expressed as means ± standard errors of three independent experiments. **P < .01, ***P < .001 compared to CTL. 2-NBDG, 2-deoxy-2-[(7-nitro-2,1,3-benzoxadiazol-4-yl) amino]-D-glucose.

The effect of RPSE on GLUT4 translocation in C2C12 myotubes. C2C12 myotubes were treated with RPSE at 200 μg/mL in differentiation media for 48 h. The plasma membrane was isolated from total cell lysates. The bands of representative experiment and relative intensity of GLUT4 and Na+/K+-ATPase α were presented. Na+/K+-ATPase α was used as the reference control. Values are expressed as means ± standard errors of three independent experiments. *P < .05 compared to CTL. GLUT, glucose transporter.
The insulin signaling pathway was analyzed, accordingly. As illustrated in Figure 7, RPSE increased the expression of IRS1 (64.89%) as well as tyrosine phosphorylated IRS1, IRS1-pY612 (58.23%, P < .01) at 200 μg/mL. The level of tyrosine phosphorylated PI3K, downstream of IRS1 was also increased about twofold compared to the control with a treatment of RPSE at 200 μg/mL (P < .01), while the expression of PI3K was not changed. Furthermore, the threonine phosphorylated Akt, Akt-pT308, was stimulated with a treatment of RPSE at 200 μg/mL (68.02%, P < .01). There was not a change in expression of Akt.
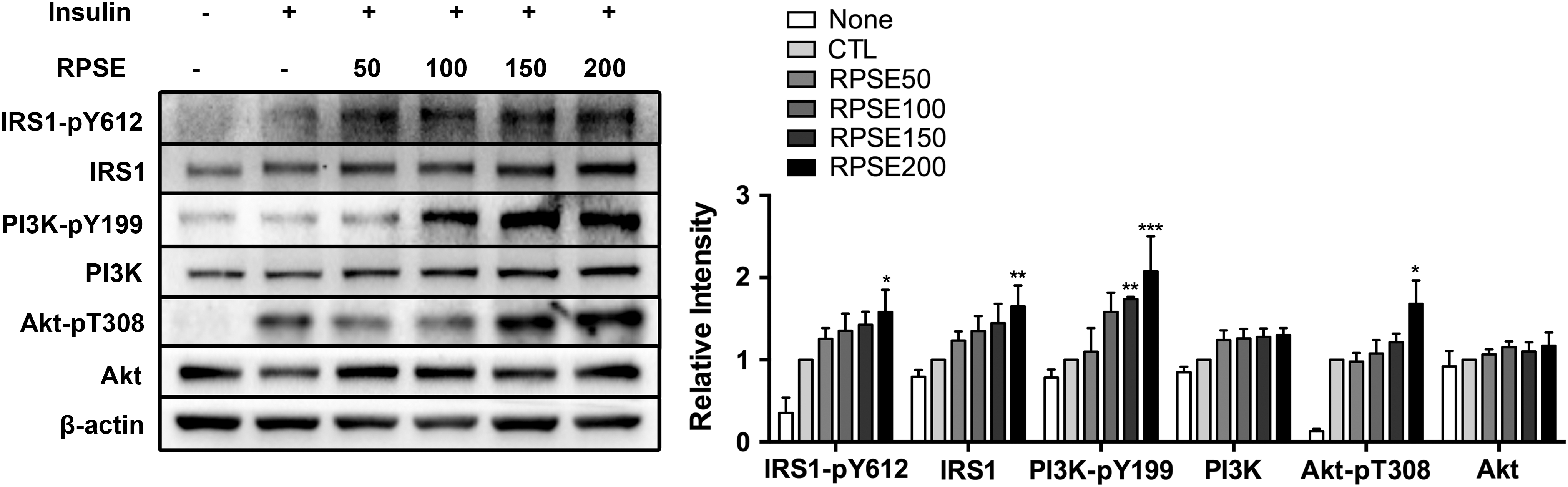
The effect of RPSE on insulin signaling related proteins in C2C12 myotubes. C2C12 myotubes were treated RPSE at concentrations of 50, 100, 150, and 200 μg/mL in differentiation media for 48 h. Cells were further incubated in the presence (CTL, RPSE50, RPSE100, RPSE150, and RPSE200) or absence (None) of 100 nM insulin for 15 min. The bands of representative experiment and relative intensity of IRS-pY612, IRS, PI3K-pY199, PI3K, Akt-pT308, and Akt were presented. β-actin was used as the reference control. Values are expressed as means ± standard errors of three independent experiments. *P < .05, **P < .01, ***P < .001 compared to CTL. IRS, insulin receptor substrate; PI3K, phosphatidylinositol 3-kinase.
Discussion
The incidence of type 2 diabetes mellitus has more than doubled over the past three decades. 18 As medical treatment of type 2 diabetes mellitus is limited, the need for physiologically effective compounds from especially natural sources is increasing. There are a number of compounds that have been known to have antidiabetic effects. 15,16 Several substances such as fucoxanthin, acacia polyphenol, and Panax ginseng berry extract have been known for having both antiobesity and antidiabetic effects. 19 –21 The antiobesity effect of RPSE was revealed already. Accordingly, we anticipated the RPSE also has antidiabetic effect.
Liver and muscle are major sites for blood glucose regulation. Glucose homeostasis is maintained by hormones and regulatory enzymes. In fed state, insulin regulates glucose metabolism, suppressing hepatic gluconeogenesis and enhancing glucose uptake in muscle. Therefore, we examined the antidiabetic effect of RPSE in AML12 hepatocytes and C2C12 myotubes. Although one of the most common liver cell-line is HepG2, HepG2 is a cancer cell-line, in which glycolysis is hyperactivated, thus raising concerns for its use in metabolism studies. It is emerging that AML12, normal murine hepatocytes, are a better alternative for gluconeogenesis experiments. 22
Cell viability test is required to evaluate physiological effect of certain substance to exclude the effect of toxicity to cells. We first examined the effect of different RPSE concentrations on cell viability in murine hepatocytes (AML12) and murine myotubes (C2C12) by MTT assay. RPSE had no toxicity at concentrations ranging from 50 to 200 μg/mL and thus based on those observations, RPSE was used at concentrations less than 200 μg/mL. Results obtained were consistent with other researches which reported that methanol RPSE did not alter cell viability in 3T3-L1 preadipocytes up to 200 μg/mL as this study. 14
To determine whether RPSE has an effect on hepatic gluconeogenesis suppression, the glucose production via gluconeogenesis was measured in AML12 cells as presented in the method above. Glycogenolysis is not the dominant source of glucose production in prolonged starvation. In addition, hepatic glycogenolysis is caused by various physiological factors, including glucagon and cortisol, which were also excluded in this study. Therefore, it was assumed that the output of glucose in AML12 was predominantly provided from gluconeogenesis. We observed the significant decrease in glucose production with RPSE. Hepatic gluconeogenesis is regulated by major enzymes responsible for gluconeogenesis such as G6Pase and PEPCK. To verify the effect of RPSE on suppression of gluconeogenesis in hepatocytes, expression of these enzymes was analyzed through immunoblotting. As presented in the results, RPSE inhibits G6Pase and PEPCK activities. Furthermore, gluconeogenic enzymes are regulated by Akt and FoxO1. The phosphorylation of Akt is followed by phosphorylation of FoxO1, which is a transcription factor regulating glucose metabolism that leads to restraint of PEPCK and G6Pase activities. The levels of phosphorylated Akt and FoxO1 were dramatically increased by RPSE. This result explains how RPSE inhibits G6Pase and PEPCK enzyme activities. Consistent with RPSE, aqueous extract of the seed of Tamarindus indica, an Indian traditional medicine, was found to have antidiabetic effects via lowered G6Pase activity in streptozotocin-induced diabetic rats. 23 Similarly, it was revealed that the levels of G6Pase and PEPCK in HepG2 human hepatocytes were lowered by treatment with cocoa flavonoids and Ginsenoside Rg1, hence decreasing glucose production. 18 This suggests that the above mentioned compounds may be helpful in type 2 diabetes. 15
Insulin resistance is ameliorated by elevated glucose uptake by skeletal muscle. The glucose uptake of myotubes was measured by 2-NBDG, fluorescent glucose analog. When cells were put in glucose-free situation, cells absorb 2-NBDG as glucose. The glucose uptake was increased by RPSE. Glucose uptake is achieved by GLUT4 translocation from cytosol to plasma membrane. To determine whether RPSE induces GLUT4 translocation, plasma membrane of C2C12 myotubes were isolated from total cell lysates. In this experiment, Na+/K+-ATPase was used as an internal control. The Na+/K+-ATPase is a universal enzyme found in cytoplasmic membrane of all eukaryotic cells. It is a well-known internal control to compare cytoplasmic membrane protein in various studies. 16,24 It was shown that RPSE stimulated GLUT4 translocation. Extract from Rooibos (Aspalathus linearis), South African herbal tea, increased glucose uptake through increased GLUT4 expression in C2C12 myotubes. 16 It was also demonstrated that upregulated glucose uptake in C2C12 treated with Tricin (5,7,4′-trihydroxy-3′,5′-dimethoxyflavone), cereal flavone was mediated by GLUT4 translocation. 19 These studies present the likelihood that RPSE stimulates GLUT4 translocation, which in turn increases glucose uptake.
It is known that insulin stimulates IRS/PI3K/Akt signaling pathway. Secreted insulin binds to IR in the membrane of muscle tissue, which stimulates phosphorylation of several intracellular substrates, including IRS proteins. 20 Phosphorylated IRS1 then triggers downstream PI3K pathway that subsequently phosphorylates Akt. Akt is stimulated through phosphorylation then induces translocation of GLUT4 from cytosol to plasma membrane in muscle cells. 21,25 The insulin signaling pathway involved proteins such as IRS1, PI3K, and Akt, and they were analyzed by immunoblotting. The results show that IRS1, PI3K, and Akt were activated by RPSE. In a similar study, it was reported that silibinin, a mixture of flavonolignans extracted from Silybum marianum seed, improved insulin sensitivity through IRS1/PI3K/Akt pathway in C2C12 myotubes. 26 Gingenoside Rg1 also improved the insulin signaling pathway in 3T3-L1. 27
In conclusion, RPSE treatment of hepatocytes results in suppression of hepatic gluconeogenesis via reduced activities of G6Pase and PEPCK. Furthermore, RPSE treatment of myotubes enhanced glucose uptake via insulin-dependent IRS/PI3K/Akt pathway. Results of this research suggest that RPSE has antidiabetic effects and can be effective if utilized in the treatment of type 2 diabetes as summarized in Figure 8.

The schematic overview showing the RPSE-induced hypoglycemic signaling pathways analyzed in AML12 hepatocytes and C2C12 myotubes. Arrows indicate positive inputs (activation) and perpendicular lines show negative inputs (inhibition).
Footnotes
Acknowledgments
This work was supported by the Korea Institute of Evaluation for Technology in Food, Agriculture, Forestry, and Fisheries (IPET) through High Value-added Food Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (115015-03-1-HD020).
Author Disclosure Statement
No competing financial interests exist.