
Abstract
Black raspberry extract (BRE) has been widely used for treating prostate and urinary diseases and hyperlipidemia in Asia due to its significant lipid-lowering effects. The aim of this investigation was to evaluate the antihypercholesterolemia activity of BRE and the potential molecular mechanisms responsible for its antihypercholesterolemia activity by regulation of proprotein convertase subtilisin/kexin type 9 (PCSK9) expression in the human liver cell line HepG2. Reporter-based functional assay was used to identify herbal extracts that suppress PCSK9 expression in the HepG2 cells. Quantitative real-time polymerase chain reaction, Western blot analysis, and immunofluorescence staining were used to evaluate whether BRE modulates low-density lipoprotein receptor (LDLR) expression by repressing the hepatic expression of PCSK9. The LDLR activity of the HepG2 cells was determined using an LDL uptake assay. Our finding revealed that BRE modulates LDLR expression by suppressing the hepatic expression of PCSK9. We found that the combination of simvastatin and BRE caused the synergic induction of LDLR expression and LDL-C uptake, whereas simvastatin alone increased the expression of PCSK9 in the HepG2 cells. These results clearly demonstrated that the BRE from black raspberry suppressed simvastatin-induced PCSK9 expression and improved LDL-C uptake by hepatocytes through the induction of LDLR expression. These results suggest that the suppression of PCSK9 expression by BRE may potentiate the hypolipidemic effect of statins.
Introduction
P
Statins belong to a class of drugs used to lower cholesterol levels by inhibiting the enzyme HMG-CoA reductase, which plays a central role in the biosynthesis of cholesterol in hepatocytes; surprisingly, statins also increase PCSK9 expression, which decreases their efficiency. 12 Potential PCSK9 inhibitors could be combined with statins to amplify their effect on LDLR activity, to overcome this unfortunate side effect of statins. Recently, monoclonal antibodies for PCSK9, alirocumab and evolocumab, have been approved by the European Commission and the FDA. 13 Thus, identifying a new antihypercholesterolemia medicine targeting PCSK9 expression is an important priority in the field of antihypercholesterolemia research.
Since inhibiting PCSK9 is a promising strategy for lowering LDL, we aimed to identify a herbal medicine that inhibits PCSK9 transcription to further enhance the plasma cholesterol-lowering actions of statins through their effect on LDLR transcription. Complementary and alternative treatments for regulating cholesterol levels and improving cardiac health have been used, and herbal medicines that induce LDLR expression could be considered an effective antihypercholesterolemia strategy. Several herbal medicines or natural products have been shown to increase LDLR expression. 14 –16
Black raspberry extract (BRE) has been widely used for treating prostate and urinary diseases and hyperlipidemia in Asia due to its lipid-lowering effects. 9,17,18 BRE contains diverse natural bioactive components, including flavonoids, phenolic acids, organic acids, and resveratrol, which regulate oxidized LDL uptake and cholesterol efflux in murine macrophages. 19,20 Although the serum cholesterol-lowering effects of BRE have been investigated, it remains unknown whether BRE treatment can influence PCSK9 expression. We measured PCSK9 and LDLR expressions in a human hepatocyte cell line, HepG2, to determine the antihypercholesterolemic effects of BRE on the regulation of PCSK9 expression. In this study, we report that BRE modulates LDLR expression by repressing the hepatic expression of PCSK9. We found that the combination of simvastatin and BRE caused the synergic induction of LDLR expression and LDL-C uptake, whereas simvastatin alone increased the expression of PCSK9 in the HepG2 cells.
Materials and Methods
Luciferase reporter constructs and plasmids
Human PCSK9 promoter fragments from −1460 to −4 bp were constructed in the pGL3 luciferase vector. A human genomic fragment spanning from −1460 to −4 bp was amplified by polymerase chain reaction (PCR) using the following oligonucleotides: forward 5-GGTACCTTCTTGCTCCAGGGGAG and reverse 5-CTCGAGGGCCAGGGGAGAGGTTGCTGTCCTG; the fragment was then cloned into the TopoTA vector (Invitrogen, USA). Next, the fragment was transferred to the PGL3 basic vector between the KpnI and XhoI sites, and the construction of the amplified DNA sequences was verified by DNA sequencing. The expression vectors HNF4α and sterol-response element binding proteins (SREBP)-1c were constructed as described previously. 21
Preparation of BRE
Dried black raspberry was purchased from Naemome DAH Co. (Ulsan, Korea). To prepare BRE, dried black raspberry (300 g) was extracted with 3 L of water at 90–100°C for 3 h. The crude extract solution was filtered, evaporated, and lyophilized in a freeze-dryer. BRE was stored at the Korea Institute of Oriental Medicine (KIOM, Daejeon, Korea) and dissolved in distilled Dulbecco's phosphate-buffered saline (DPBS) before its application in cells.
Cell culture
HepG2 cells were obtained from the American Type Culture Collection (Manassas, VA). The cells were maintained in a 1:1 mixture of Dulbecco's Modified Eagle's Medium and F-12 (DMEM/F12, 50:50; Life Technologies, USA) supplemented with 100 U/mL penicillin G/streptomycin sulfate (Invitrogen) and 10% heat-inactivated fetal bovine serum (Life Technologies).
Transient transfection and luciferase reporter assay
For the luciferase reporter assay, HepG2 cells were plated in 24-well plates 24 h before transfection; then, the cells were transfected with the luciferase reporter using the TranslT-LT1 transfection reagent (Mirus Bio, USA) according to the manufacturer's instructions. For each transfection reaction, the Renilla luciferase control driven by the SV40 early enhancer/promoter was mixed with the firefly luciferase reporter construct at 1:30 molar ratio. Cell lysates were assayed for firefly and Renilla luciferase activity using the Dual-Glo® Luciferase Assay System (Promega, Madison, USA) and a TriStar LB941 Luminometer (Berthold Technologies, Germany) according to the manufacturer's instructions. The firefly luciferase activity of individual transfections was normalized to the Renilla luciferase activity.
RNA isolation and quantitative real-time PCR
Total RNA was isolated using the RNeasy Mini Kit (Qiagen, Germany) according to the manufacturer's instructions. To detect relative mRNA expression, reverse transcription and quantitative real-time PCR (Q-PCR) were performed with TaqMan probes (Life Technologies) as described previously. 21 Relative amounts of cDNA were calculated by using the relative quantification (ΔΔCt) method. Experiments were repeated at least thrice with different cell preparations. The relative abundance of each transcript was normalized to that of ubiquitin C.
Western blotting analysis
The total protein was extracted from cells and lysed. A total of 30 μg of each protein sample was separated on a sodium dodecyl sulfate–polyacrylamide gel and transferred to a polyvinylidene difluoride membrane. The membranes were blocked for 1 h at room temperature with a 5% skim milk solution. Then, the membranes were incubated overnight at 4°C with antibodies specific to PCSK9, LDLR, or β-actin. The blots were washed thrice for 10 min each time with Tris-buffered saline with Tween-20 (TBST). The membranes were then incubated for 2 h with anti-rabbit secondary antibody. Proteins were detected using enhanced chemiluminescence solution, and the bands were visualized using a chemiluminescence imaging system (ImageQuant LAS 4000 mini; GE Healthcare, United Kingdom).
Enzyme-linked immunosorbent assay analysis
The Human PCSK9 SimpleStep ELISA kit was purchased from Abcam (United Kingdom). HepG2 cells were seeded in a 12-well plate at 1 × 105 cells/well and grown for 24 h; then, the cells were washed twice with DPBS and starved in DMEM/F12 supplemented with 0.5% FBS for 24 h. Next, the cells were washed twice with DPBS and treated with a statin with or without BRE (10, 50 and 100 μg/mL) in DMEM/F12 supplemented with 0.5% FBS for 24 h; finally, the media were collected. PCSK9 levels in the culture media were measured using the human PCSK9 ELISA kit according to the manufacturer's instructions.
LDLR immunofluorescence staining
To determine the protein expression levels of LDLR, HepG2 cells were seeded in glass base dishes (ThermoFisher Scientific, USA) at 1 × 104 cells/dish and grown for 24 h. The cells were washed twice with DPBS and starved in DMEM/F12 supplemented with 0.5% FBS for 24 h; then, the cells were washed twice with DPBS and treated with a statin with or without BRE (100 μg/mL) in DMEM/F12 supplemented with 0.5% FBS for 24 h. The cells were maintained at room temperature until the end of the protocol. The cells were then fixed with 1% paraformaldehyde in PBS for 15 min; next, the cells were permeabilized with 0.1% Triton X-100/PBS containing 2% BSA and blocked with 0.5% normal serum to reduce the nonspecific adherence of antibodies. The cells were incubated with primary anti-LDLR antibody at a dilution of 1:200 for 1 h at 37°C in a humidified chamber. After incubation with the primary antibody, the cells were rinsed and incubated with anti-rabbit IgG-FITC (Santa Cruz Biotechnology, USA). The cell nuclei were stained with Hoechst stain (blue) and then washed with PBS and treated with mounting solution (Wako, Kumamoto, Japan). Finally, confocal images were captured with an Olympus FV10i-W confocal laser scanning microscope.
LDL uptake assay
The LDLR activity of the HepG2 cells was determined using an LDL uptake assay kit (Biovision, USA) according to the manufacturer's instructions. Briefly, HepG2 cells grown on 24-well plates were starved in 0.5% serum-containing media for 12 h. The media were removed, and the cells were washed thrice with an assay buffer. Diluted fluorescently labeled LDL was added with or without BRE to each well. To determine the specificity of LDL uptake, the signal was competed out by adding unlabeled LDL to the fluorescently labeled LDL in one of the wells. The cells were incubated for 24 h, and the media were then removed. The cells were carefully washed four times with the assay buffer, and 100 μL of assay buffer was added to each well. The fluorescence intensity of each well was measured with a fluorescence spectrophotometer (Molecular Devices SpectraMax M2; excitation: 540 nm; emission: 575 nm).
HMG-CoA reductase activity assay
The effect of BRE on HMG-CoA reductase activity was determined using the HMG-CoA Reductase Activity/Inhibitor Screening Kit (Biovision) according to the manufacturer's instructions. Briefly, HMG-CoA reductase and BRE were added to a 96-well plate. For the positive control, 5 μL of HMG-CoA reductase was added to one of the wells. For the inhibitor control, 5 μL of HMG-CoA reductase and 2 μL of an inhibitor (atorvastatin, 100 μM) were added to one of the wells. For the reagent background control, 10 μL of HMG-CoA reductase assay buffer was added to one of the wells. The reaction was initiated with the HMG-CoA, NADPH, and HMG-CoA reductase assay buffer. Immediately after the reaction, the absorbance was measured kinetically at OD 340 nm in a microplate reader.
Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) assays were performed using a ChIP assay kit (Upstate Cell Signaling Solutions, USA) with slight modifications to the manufacturer's instructions. HepG2 cells were crosslinked in 1% formaldehyde and then sonicated. A cell lysate solution (5%) in ChIP dilution buffer was set aside for use as the “Input.” Anti-hepatocyte nuclear factor 1α (HNF1α) (Santa Cruz Biotechnology) was added to precipitate DNA-protein complexes, and nonimmune mouse IgG (Santa Cruz Biotechnology) was used as a control. A 199-bp DNA fragment (−490 to −292) containing the HNF1α binding site of the human PCSK9 promoter was amplified by PCR for 30 cycles and then separated on a 1.5% agarose gel. The PCR primers used for amplification were as follows: forward primer, 5′-TCCAGCCCAGTTAGGATTTG-3′; reverse primer, 5′-CGGAAACCTTCTAGGGTGTG-3′. As a negative control, PCR amplification of the human GAPDH promoter was performed using forward (5′-ATGGTTGCCACTGGGGATCT-3′) and reverse (5′-TGCCAAAGCCTAGGGGAAGA-3′) primers.
Statistical analysis
All experimental data are shown as the mean value ± SEM. Multiple groups were tested by one-way analysis of variance, and Dunnett's multiple comparison test was used to determine which groups were significantly different from the control group. A P value < .05 was considered to be significant. *P < .05; **P < .01
Results
BRE suppresses human PCSK9 promoter activity in HepG2 cells
We first validated the sensitivity and specificity of the reporter in HepG2 cells. Human PCSK9 promoter-Luc (hPCSK9-Luc) and an internal control pRL-TK-Luc were cotransfected. Transfected cells were starved for 24 h in media containing 0.5% FBS and were treated with vehicle (0.1% DMSO), berberine (BBR), simvastatin, or a water extract of Coptis chinensis (CCE); then, the luciferase activity was determined after 24 h. Since BBR was a known inhibitor of PCSK9 expression and CCE contained BBR, they were used as experimental controls. BBR and CCE dramatically inhibited the hPCSK9 promoter activity (Fig. 1A). To validate these reporter systems, we also cotransfected hPCSK9-Luc with an expression vector coding for HNF4α and a mature transcriptionally active SREBP-1c. As a result, SREBP-1c, but not HNF4α, increased the hPCSK9-Luc activity by fivefold (Fig. 1B).
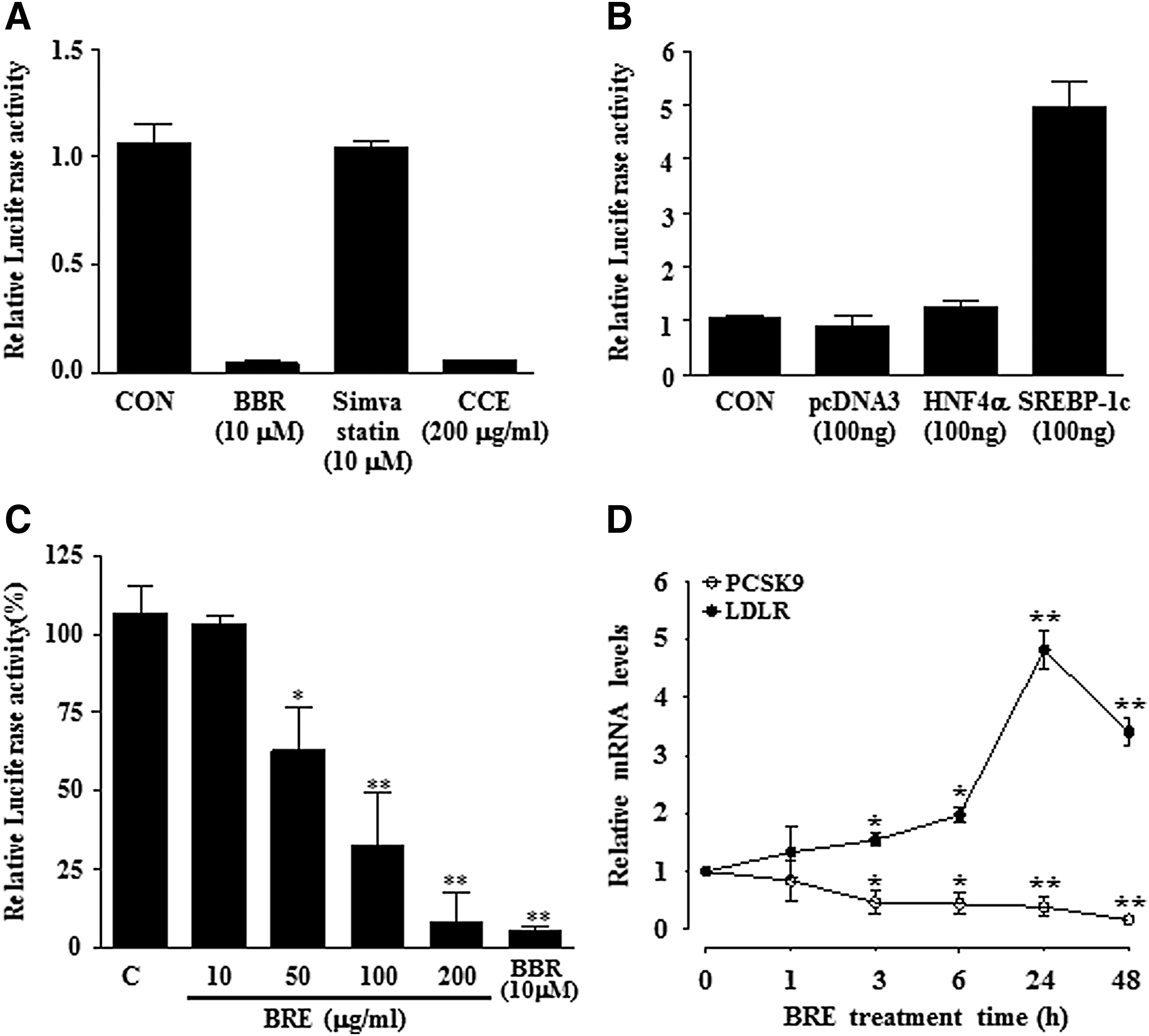
Luciferase reporter assay screening of a herbal extract library for inhibitors of human PCSK9 promoter activity.
Because natural extracts have been long recognized as a potential source of therapeutic drugs, we used a cell-based reporter gene assay to screen natural product extracts for those that inhibited the human PCSK9 promoter activity in HepG2 cells. To identify these extracts, we plated HepG2 cells in 24-well plates, transfected them with the human PCSK9 promoter reporter, and then treated them with 240 natural extracts at a final concentration of 100 μg/mL. After a 24-h incubation, the luciferase activity was measured and normalized. We focused on herbal extracts that inhibited the PCSK9 promoter activity to below 50% compared to that of the vehicle controls. The initial hits were retested in a dose-dependent manner. From this first screen, we identified a herbal extract, BRE that significantly inhibited human PCSK9 promoter reporter luciferase activity by more than 50% (Fig. 1C).
Moreover, HepG2 cells incubated with BRE extract showed reciprocal changes in the expression of two genes directly involved in LDL uptake; LDLR expression levels were upregulated, and PCSK9 expression levels were downregulated (Fig. 1D). This experiment revealed that BRE highly induced and suppressed LDLR and PCSK9 gene expression levels, respectively, at 24 h after BRE treatment and at the six subsequent time points. Therefore, we used a similar dose range in this study to determine the reciprocal changes in the expression patterns of PCSK9 and LDLR at 24 h.
BRE attenuates simvastatin-induced expression of PCSK9, but increases the expression of LDLR and LDL uptake in HepG2 cells
To further confirm whether BRE represses human PCSK9 gene expression with or without the addition of a statin, we examined PCSK9 mRNA and protein levels in HepG2 cells by Q-PCR and immunoblotting. HepG2 cells were treated with BRE (10, 50, 100, and 200 μg/mL) and simvastatin for 24 h, and PCSK9 mRNA and protein levels were decreased in a dose-dependent manner (Fig. 2A, C).
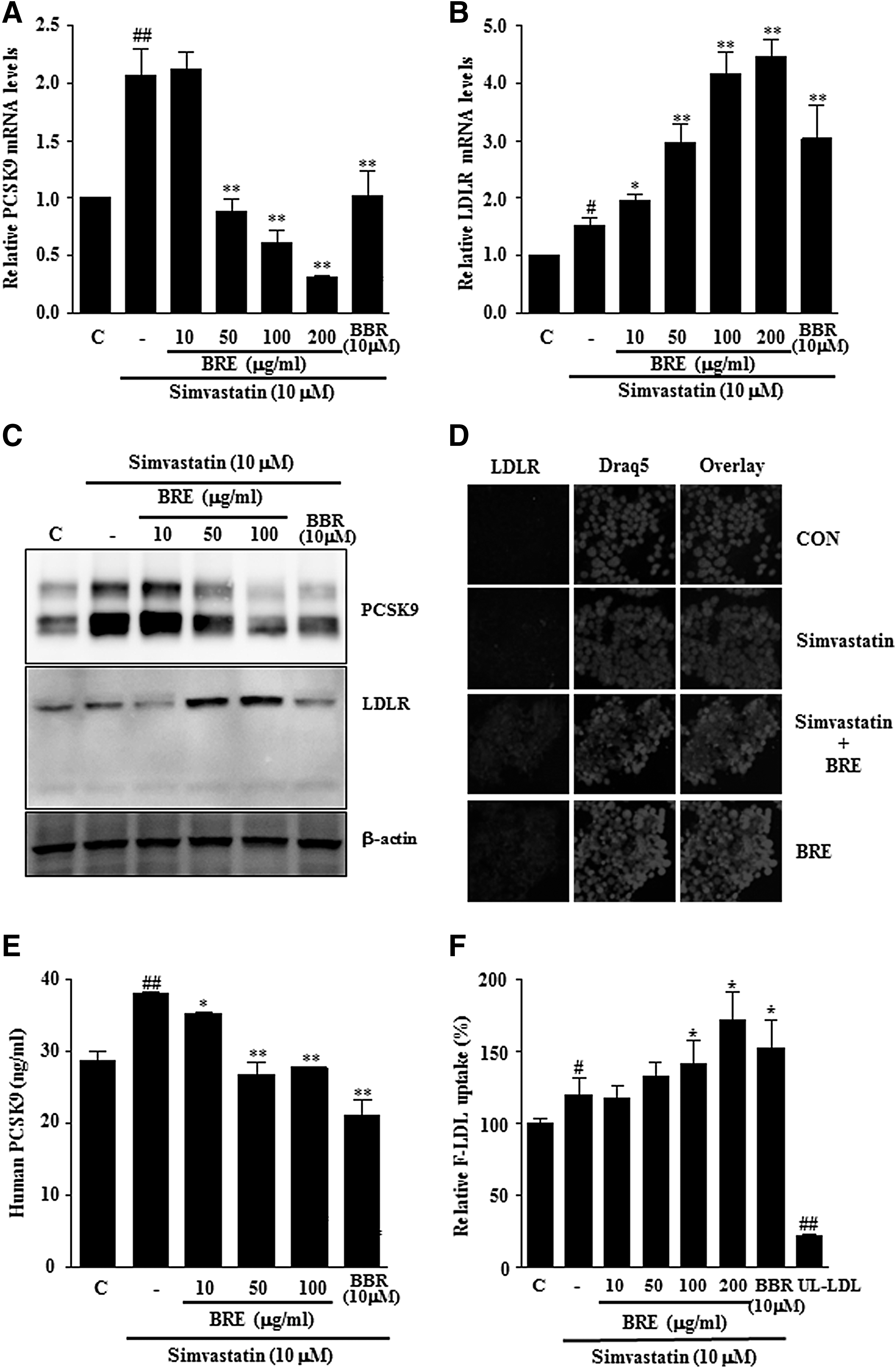
The effect of BRE on PCSK9 and LDLR expression. HepG2 cells were treated with vehicle (C, PBS), BRE (10–200 μg/mL), and BBR (10 μM) with or without simvastatin for 24 h. mRNA levels of PCSK9
We also examined the effects of BRE on LDLR gene expression. BRE and simvastatin treatment synergistically increased LDLR mRNA and protein expression levels in HepG2 cells (Fig. 2B–D). We examined whether decreased PCSK9 expression caused by BRE was associated with decreased PCSK9 secretion. Simvastatin alone significantly increased the secretion of PCSK9 into the culture media, but cotreatment with BRE and simvastatin inhibited the secretion of PCSK9 (Fig. 2E).
Since treating HepG2 cells with BRE increased the expression levels of LDLR and decreased the expression levels of PCSK9, we evaluated the effect of BRE on cellular LDL uptake. We quantified the uptake of fluorescently labeled LDL by HepG2 cells treated with various concentrations of BRE with or without simvastatin for 24 h. Treatment with BRE resulted in a significant increase in the rate of labeled LDL uptake by ∼50% compared to that of cells treated with simvastatin alone, suggesting that the increased expression of LDLR induced by BRE treatment led to functionally enhanced LDL-C uptake by hepatocytes (Fig. 2F).
BRE modulates HMG-CoA reductase gene expression and activity
We next examined whether BRE enhanced the inhibitory effect of simvastatin on HMG-CoA reductase. As shown in Figure 3A, BRE inhibited the HMG-CoA reductase activity in a concentration-dependent manner. Moreover, HMG-CoA reductase mRNA was significantly increased by simvastatin treatment, but cotreatment with simvastatin and BRE suppressed this increase (Fig. 3B).
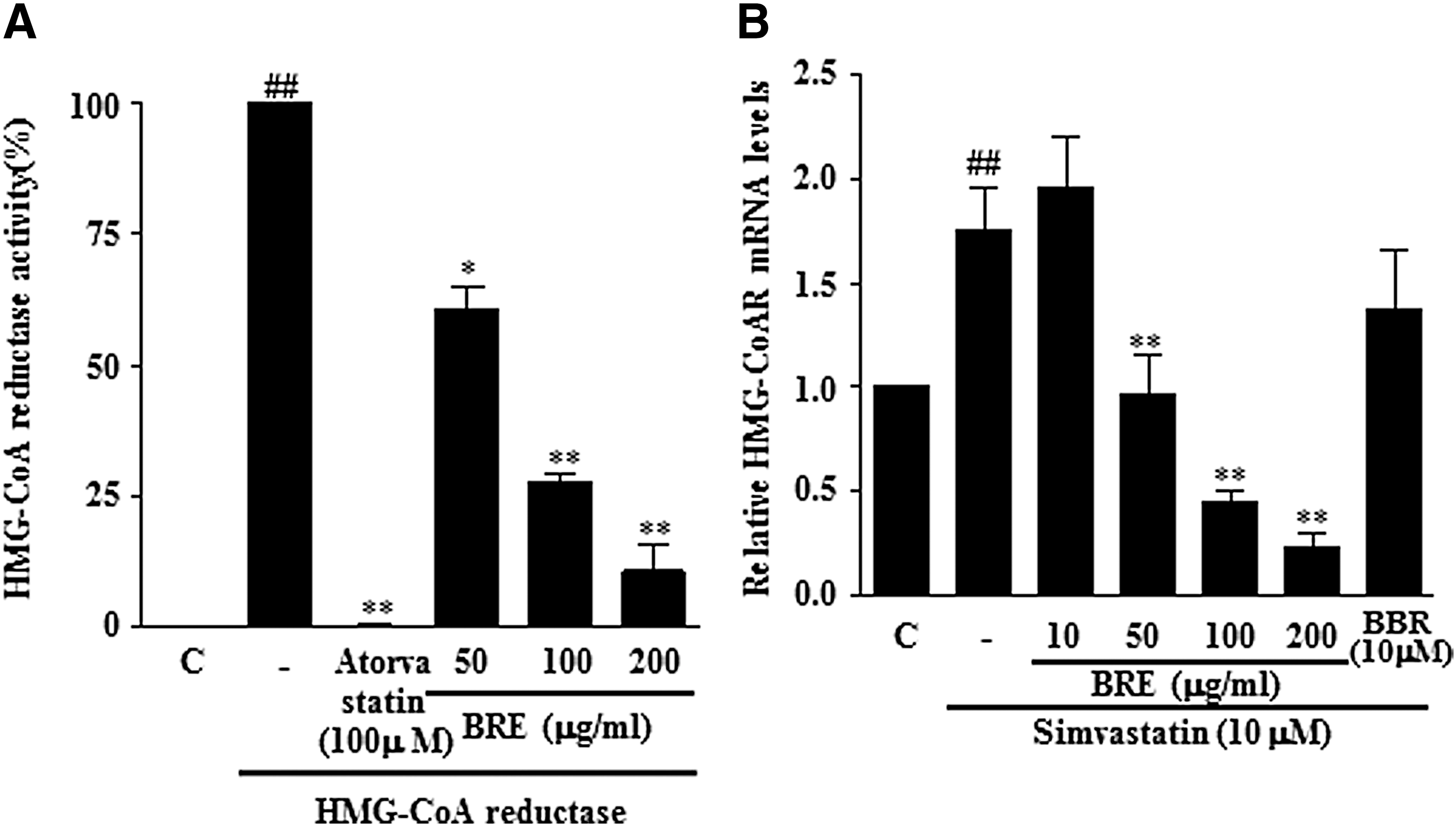
Inhibitory effect of BRE on HMGCR activity and mRNA expression induced by simvastatin.
BRE reduced HNF1α expression and binding activity toward the human PCSK9 promoter in cultured hepatic cells
As HNF1α is critically involved in PCSK9 transcription, we evaluated the effect of BRE on HNF1α mRNA levels in HepG2 cells. Notably, the Q-PCR and Western blot results revealed that the expression of HNF1α was significantly decreased by BRE treatment (Fig. 4A, B). Since previous studies have reported that HNF1α plays a critical role in PCSK9 gene transcription and our Q-PCR and Western blot results revealed that BRE treatment decreased HNF1α expression, we speculated that the BRE-mediated decrease in PCSK9 expression involves HNF1α.
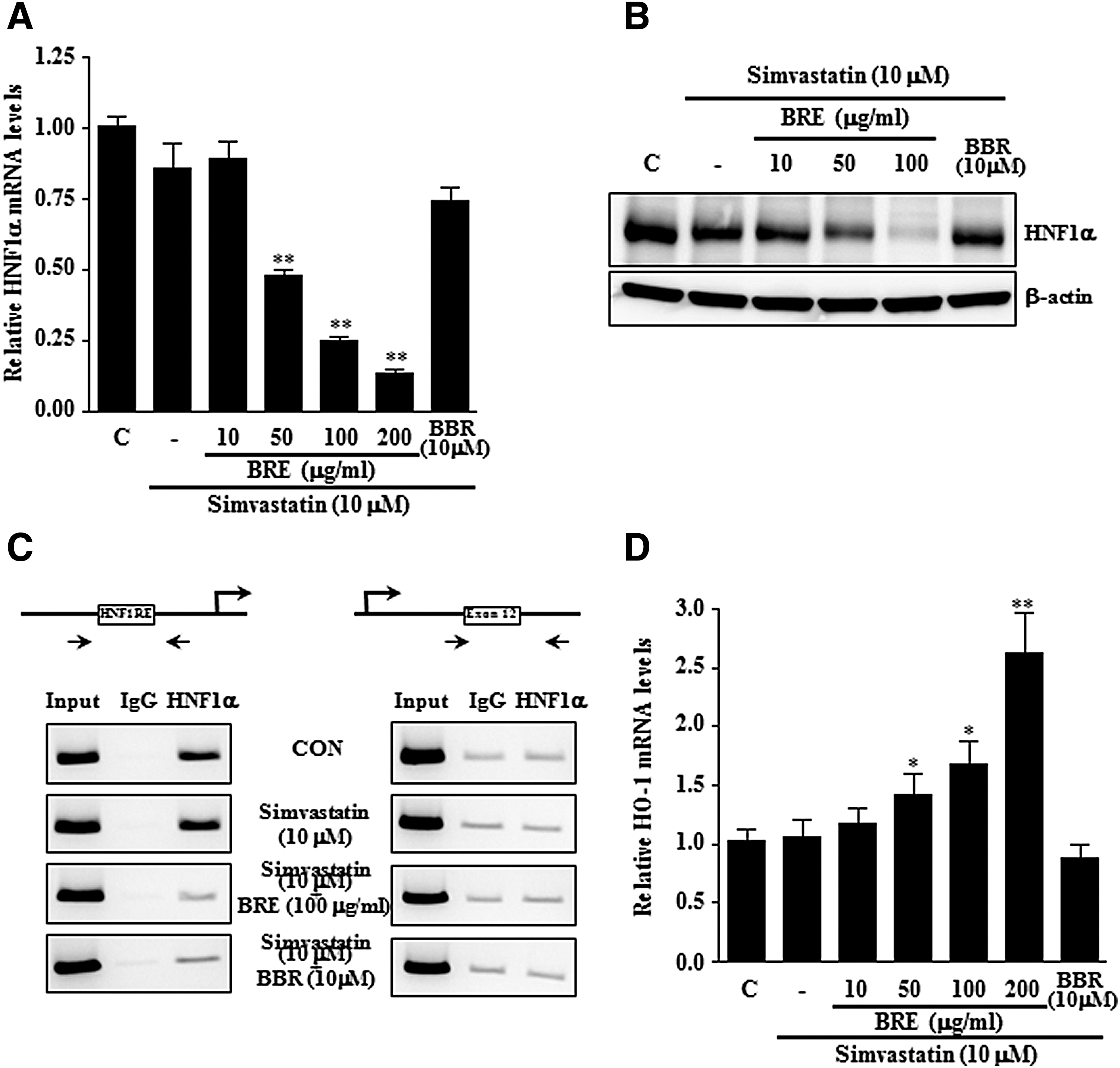
BRE reduced HNF1α expression in HepG2 cells. HepG2 cells were treated with vehicle (C, PBS), BRE (10–200 μg/mL), and BBR (10 μM) with or without simvastatin for 24 h. mRNA and protein levels of HNF1α were measured by Q-PCR
To further validate our finding, we used ChIP assays to determine the transcription factor binding activity toward the promoter (Fig. 4C). PCR amplification of a region containing the HNF1α binding region of the hPCSK9 promoter indicated that the accumulation of HNF1α at the promoter site was decreased by BRE and BBR treatment; however, no significant recruitment of HNF1α was observed at the exon 12 region, which served as a negative control. In addition, no PCR amplification was observed when immunoprecipitation assays were carried out using mouse IgG. These findings showed that BRE suppresses HNF1α expression, thus reducing its transcriptional activity, and this BRE effect effectively suppressed simvastatin-induced PCSK9 expression.
Several studies have indicated that the therapeutic benefits of statins involve not only their cholesterol synthesis inhibition effects but also their cholesterol-independent cytoprotective pleiotropic effects, including heme oxygenase-1 (HO-1) induction. Since HO-1 is an important mediator of the cholesterol-independent cytoprotective actions of statins, we also examined whether BRE and simvastatin can induce the expression of HO-1 in HepG2 cells. As shown in Figure 4D, a significant increase in HO-1 mRNA was observed after BRE and simvastatin treatment. However, BBR and simvastatin treatment showed little synergistic induction effects on HO-1 mRNA expression.
Discussion
Hypercholesterolemia is a major risk factor for atherosclerosis and CVD. Lowering plasma cholesterol levels is expected to be beneficial in treating hypercholesterolemia-related diseases or disorders, including metabolic syndrome. Statin therapy is recommended for most patients because statins are effective cholesterol-lowering drugs that have been directly associated with a reduced risk of CVDs. Although statins increase LDLR expression by inhibiting cholesterol synthesis, which leads to the clearance of cholesterol from the blood, they have also been reported to induce the expression of PCSK9, which causes the degradation of the LDLR in hepatocytes and limits the effects of statins. Therefore, a combination of lipid- lowering medicines is currently used, particularly in patients whose cholesterol levels are not decreased when using only statins. Previous studies have implied that lipid-lowering small molecules have effects on PCSK9, but the impacts of combinations of lipid-lowering drugs on PCSK9 have not yet been extensively evaluated. 14,22
In this study, the antihypercholesterolemia activity of BRE and the potential molecular mechanisms implicated in its antihypercholesterolemia activity were investigated in the human liver cell line HepG2. We screened various compounds and identified BRE as a suppressor of the upregulation of PCSK9 expression caused by statins in hepatocytes. Moreover, we also demonstrated that the combination of BRE and a statin significantly increased LDLR expression and LDL uptake in hepatocytes. These results suggest that BRE has potential as a cholesterol-lowering drug for treating patients who do not achieve their therapeutic goals with a statin alone or patients with only mild hypercholesterolemia.
Furthermore, BRE has been commonly used in oriental medicine because of its ability to improve blood circulation. Clinical studies have found that dietary black raspberry supplementation significantly decreased LDL-C levels in patients with metabolic syndrome. 18 Therefore, BRE may contribute to improving LDL-C clearance from the blood by increasing LDLR gene expression due to the reduced HMG-CoA reductase and protecting LDLR from PCSK9-mediated degradation. To test these hypotheses and verify our results in vivo in humans, the amount of circulating PCSK9 and LDL in patients would need to be evaluated before and after treatment with BRE with or without a statin. Nevertheless, our data suggest that BRE might be an interesting candidate for treating hypercholesterolemic patients in whom statin therapy alone does not achieve their therapeutic goals; furthermore, BRE alone exerts potent hypolipidemic activity, suggesting that it could be a potential adjuvant preventative treatment for patients with CVD.
Several studies have shown that various proteins regulate PCSK9 gene expression, including SREBP-1/2, HNF1α, farnesoid X receptor (FXR), and peroxisome proliferator-activated receptor γ (PPARγ). 1,23 –25 Coincubation with fenofibric acid and pravastatin also significantly impaired the upregulation of PCSK9 protein expression. This effect was further confirmed at the transcriptional level by showing that fenofibrate alone and fenofibrate with pravastatin suppressed wild-type PCSK9 promoter activity. 24 We found that BRE caused no significant change in SREBP-2 levels in HepG2 cells (data not shown), but strongly suppressed HNF1α expression and transcriptional activity, such as DNA binding. These results are similar to a recent report that a BBR-mediated reduction in HNF1α protein by the ubiquitin proteasome system negatively regulates PCSK9 gene expression in HepG2 cells. 26
HO-1 is an inducible isoform of HO that plays important antioxidant and anti-inflammatory roles and contributes to protection from vascular inflammation in atherosclerosis. 27,28 The induction of HO-1 after BRE and simvastatin treatment implies the vasculoprotective effects of BRE. Induced HO-1 expression possibly inhibits lipid peroxidation and may result in the attenuation of the development of atherosclerotic lesions. The active components of BRE are purported to be flavones, including ellagic acid, gallic acid, caffeic acid, and tiliroside. Dietary BRE consumption may cause the accumulation of active ingredients within the circulatory system, as well as in hepatocytes, thus affecting hepatic PCSK9 and LDLR expression. We also used HPLC to standardize the extracts and set the index components that characterize the manufacturing process, and confirm the content of these single substances (data not shown). Furthermore, we are in the process of separating and identifying the core compounds responsible for the observed effects from the extract. These compounds may provide useful tools for investigating the functions of PCSK9 during statin treatment.
In conclusion, we showed that BRE, a herbal medicine, may be a potential alternative intervention that can be used in combination with other drugs for lower circulating lipids in patients with hypercholesterolemia. The sustained effects of BRE included the suppression of simvastatin-induced PCSK9 expression and the improvement of LDL-C uptake by hepatocytes by the induction of LDLR expression. These results suggest that the suppression of PCSK9 expression by BRE may potentiate the hypolipidemic effects of statins. Although this study has demonstrated the effects of BRE at the cellular level, further in vivo studies may provide additional understanding of the pharmacological function of BRE and support the potential usefulness of BRE as an effective novel antihypercholesterolemia drug.
Footnotes
Acknowledgments
This research was supported by the Development of Herbal Medicine for Aging-Related NAFLD (K15302) funded by KM Convergence Research Division of Korea Institute of Oriental Medicine (KIOM) and Technical Commercialization of Anti-Dyslipidemic Material (K15700) funded by the Mibyeong Research Center of KIOM. The author thanks Dr. Ohn-Soon Kim for technical assistance with HMG-CoA reductase activity analysis.
Author Disclosure Statement
No competing financial interests exist.