
Abstract
Echinacea purpurea has been widely used for the prevention and treatment of upper respiratory tract infections and the common cold. The restraint stress has been reported to suppress a broad spectrum of immune functions. The aim of this study was to investigate the protective effects of the pressed juice of E. purpurea (L.) Moench (EFLA®894; Echinacea) against restraint stress-induced immunosuppression in BALB/c mice. Echinacea significantly normalized the restraint stress-induced reduction in splenocyte proliferation and splenic natural killer (NK) cell activity (P < .05). Echinacea treatment significantly increased the percentages of CD4+ and CD8+ T lymphocytes in the blood (P < .05). In addition, Echinacea restored serum cytokine levels, including interleukin-6 (IL-6), interleukin-10 (IL-10), and interleukin-17 (IL-17), as well as the mRNA expressions of these cytokines in spleen (P < .05). Our findings suggest that Echinacea might have beneficial effects on restraint stress-induced immunosuppression by increasing splenocyte proliferation and NK cell activity, while modulating T lymphocyte subsets and cytokine levels in the blood.
Introduction
T
The negative effects of restraint stress on immune function are well documented. 3,5,6 Restraint affects both innate and adaptive immunity, as demonstrated by the decreased lymphocyte proliferation, cytotoxic activity of natural killer (NK) and T cells, and antibody production in mice. The suppression of circulating CD4+ and CD8+ T lymphocyte subsets in mice under restraint stress has been reported. Cytokines, such as interleukin-6 (IL-6), interleukin-10 (IL-10), and interleukin-17 (IL-17), which are related to autoimmune disorders and inflammation, are increased in restraint-stressed mice. 7,8 In addition, restraint stress leads to decreased expression of cytokines, such as IL-2 and IFN-γ, which are associated with T helper 1 (Th1) cells. 9
There have been efforts to regulate immune homeostasis with medicinal herbs. 10 Among these, the genus Echinacea (Asteraceae family), in particular, is widely used in European countries and the United States due to its proven immuno-enhancing and anti-inflammatory effects. 11 Especially, Echinacea purpurea has been cherished for the prevention and treatment of upper respiratory tract infections and common cold. 12 –14 The alkylamides and polysaccharides such as β-1,2-D-fructofuranosides have been suggested to be bioactive compounds of Echinacea. 15
Immunomodulation by Echinacea seems to occur by stimulation of NK cell activity in human peripheral blood mononuclear cells. 16 Also, Echinacea effectively induced cytokine production, in particular the early immune response mediators such as IL-l, TNF-α, and IL-10. 17 The same preparation improved immunological function in humans, especially T cell-related immunity. 18 However, the modulatory effects of Echinacea on restraint stress-induced immunosuppression in mice have not yet been tested. Therefore, we aimed to investigate the effects of Echinacea on immune function in restrained mice, with an emphasis on splenocyte proliferation and NK cell activity in the spleen and on blood T lymphocyte subsets and cytokine production.
Materials and Methods
Preparation of Echinacea
The Echinacea sample (EFLA®894) was provided by Frutarom (Wädenswil, Switzerland). Cold-pressed juice of fresh aerial parts of E. purpurea (L.) Moench harvested at flowering time was vacuum-belt dried. The sample to extract ratio was 40–50:1 (by weight), related to the fresh collected plant. The contents of fructofuranosides and alkylamides in Echinacea preparations are shown in supplementary data.
Animal experiments
Male BALB/c mice (6-week-old, 18–20 g, n = 70) were purchased from Doo Yeol Biotech (Seoul, Korea) and individually housed in a plastic cage maintained at a controlled temperature (22°C ± 2°C), humidity (55% ± 5%), and lighting (12-h light–12-h dark cycle). Normal chow (2018S Teklad rodent diet; Envigo, Indianapolis, IN, USA) and water were available ad libitum throughout the study. Experiments were performed in accordance with the Institutional Animal Care and Use Committee of Ewha Womans University (IACUC No. 15-101). After 1 week of acclimation, the mice were randomly divided into five groups: normal group, without any restraint or orally administered supplement; restraint-induced immunosuppression (IS) group, with restraint and the 0.9% saline vehicle treatment; and IS + Echinacea (at doses of 10, 30, and 100 mg/kg of body weight, respectively) groups. The treatments were orally administered once daily for 2 weeks. The mice were restrained individually in a well-ventilated horizontal 50 mL disposable polypropylene conical tube to induce immunosuppressive conditions. To test the effects of Echinacea on spleen cell proliferation and NK cell activity, mice (n = 6 per group) were restrained for 18 h, 3 days before sacrifice. At the end of the experiment, all mice were anesthetized using a combination of Zoletil (Virbac Laboratories, Carros, France) and Rompun (Bayer Korea, Seoul, Korea). Spleens were harvested and single splenocytes were isolated.
Another experiment was designed to test the effects of Echinacea on blood T lymphocytes, serum cytokine levels, and cytokine mRNA expression in spleen. Mice (n = 8 per group) were restrained for 18 h, 1 day before sacrifice. Supplementation and anesthesia were given as described in the above experiment. Blood was collected by cardiac puncture and either centrifuged (2000 g, 16°C, 20 min) to separate serum or immediately mixed with 0.1 M EDTA for flow cytometric analysis. Spleens were frozen immediately in liquid nitrogen and stored at −70°C for analysis.
Spleen cell proliferation assay
Spleen tissues were minced by a syringe with 10 mL RPMI 1640 medium containing 10% fetal bovine serum, 10,000 U/mL penicillin, 10,000 μg/mL streptomycin, and 25 mg/mL Fungizone®. The homogenates were filtered through a 40-mm cell strainer and centrifuged (800 g, 4°C, 5 min). After discarding the supernatant, splenocytes were resuspended in ammonium-chloride-potassium lysing buffer for 10 min to lyse erythrocytes and then washed in RPMI 1640 medium. Splenocytes were resuspended in RPMI 1640 and cultured in preparation for assays.
The proliferation of spleen cells was assessed using a Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan).The spleen cells were seeded at a concentration of 2 × 105 cells/well in 96-well culture plates. The mitogen response assay was performed by co-culture of splenocytes with concanavalin A (ConA; 5 μg/mL) or lipopolysaccharide (LPS; 10 μg/mL) and incubated for 48 h at 37°C. The cultures were treated with 10 μL/well of CCK-8 solution for at least 4 h, and absorbance were measured at 450 nm using a Varioskan plate reader (Thermo Electron, Waltham, MA, USA). A proliferation index was calculated as the fold change relative to the unstimulated IS group.
Splenic NK cell activity
Splenic NK cell activity was assessed using a lactate-dehydrogenase (LDH) cytotoxicity detection kit (Takara, Tokyo, Japan). YAC-1 cells were obtained from the Korean Cell Line Bank (KCLB, Seoul, Korea). Spleen effector cells (5 × 105 cells) and YAC-1 target cells (5 × 104 cells) were seeded at a 10:1 ratio into 96-well culture plates and incubated for 4 h at 37°C. Cell culture supernatants were centrifuged at 1800 rpm on an Eppendorf Centrifuge 5810 R (Eppendorf AG, Hamburg, Germany) for 10 min, and the LDH activity was quantitated by measuring absorbance at 490 nm. NK cell activity was calculated by the following formula: NK cell activity (%) = experimental release − spontaneous release/maximum release − spontaneous release × 100.
Blood T lymphocyte immunophenotyping
The blood was incubated for 10 min at 4°C with purified CD16/32-specific antibodies to block the Fc (fragment, crystallizable) receptor (BioLegend, San Diego, CA, USA) and then stained with phycoerythrin anti-mouse CD3, fluorescein isothiocyanate anti-mouse CD4, and allophycocyanin anti-mouse CD8a monoclonal antibodies (BioLegend). After incubation, red blood cells were lysed using FACS lysing solution (BD Biosciences, San Jose, CA, USA) for 15 min at room temperature and then washed in FACS buffer (BioLegend). Samples were resuspended in FACS buffer and stored at 4°C in darkness before analysis using a flow cytometer (FACS Calibur; BD Biosciences) and Cell Quest Pro Software (BD Biosciences).
Serum cytokine concentration
The serum osteoprotegerin (OPG) was measured using a Quantikine ELISA Kit (R&D Systems, Minneapolis, MN, USA). Serum levels of IL-6, IL-10, and IL-17 were simultaneously analyzed using Luminexx MAP technology and kits (MCYTOMAG-70K-15; Millipore, Billerica, MA, USA). Median fluorescence intensity values were analyzed in 3.1 xPONET software using a five-parameter logistic method to calculate cytokine concentrations by comparison with standard curves.
Real-time quantitative reverse transcription–polymerase chain reaction
Total RNA was isolated from spleen using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA). The corresponding cDNA was synthesized using an M-MLV Reverse Transcriptase kit (Bioneer Co., Daejeon, Korea), and quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was performed using AccuPower 2X Greenstar qPCR Master Mix (-ROX Dye) (Bioneer Co.) on a fluorometric thermal cycler (Rotor-Gene 3000; Corbett Research, Sydney, Australia). The sequences of the sense and antisense primers are presented in Table 1. The ΔΔCt method was used for relative quantification. 19 Values were expressed as a fold change over the IS group.
Il-6, interleukin-6; Il-10, interleukin-10; Il-17, interleukin-17.
Statistical analysis
All data are expressed as the mean ± SEM. Statistical analyses were performed using SPSS software version 19 (SPSS, Inc., Chicago, IL, USA). Quantitative data were analyzed using one-way analysis of variance, followed by polynomial contrast and Tukey's post hoc multiple comparison tests. P < .05 was considered statistically significant.
Results
Spleen cell proliferation
Splenocyte proliferation was assayed in the absence or presence of mitogen (ConA or LPS). As shown in Figure 1, splenocyte proliferation was inhibited in the IS group compared with that in the normal group. Splenocyte proliferation index was increased by Echinacea treatment in unstimulated or ConA- or LPS-stimulated splenocytes (linear effect, P < .001, P < .001 and P < .001, respectively). In unstimulated splenocytes, oral administration of Echinacea at 30 and 100 mg/kg enhanced splenocyte proliferation compared to the IS group. Similarly, in the presence of ConA, splenocyte proliferation was significantly inhibited in the IS group compared to the normal group, whereas oral administration of Echinacea at 30 and 100 mg/kg enhanced splenocyte proliferation compared to the IS group. With LPS, the proliferation index of the IS group decreased compared to that of the normal group. Administration of Echinacea at 10, 30, and 100 mg/kg enhanced splenocyte proliferation compared to the IS group.
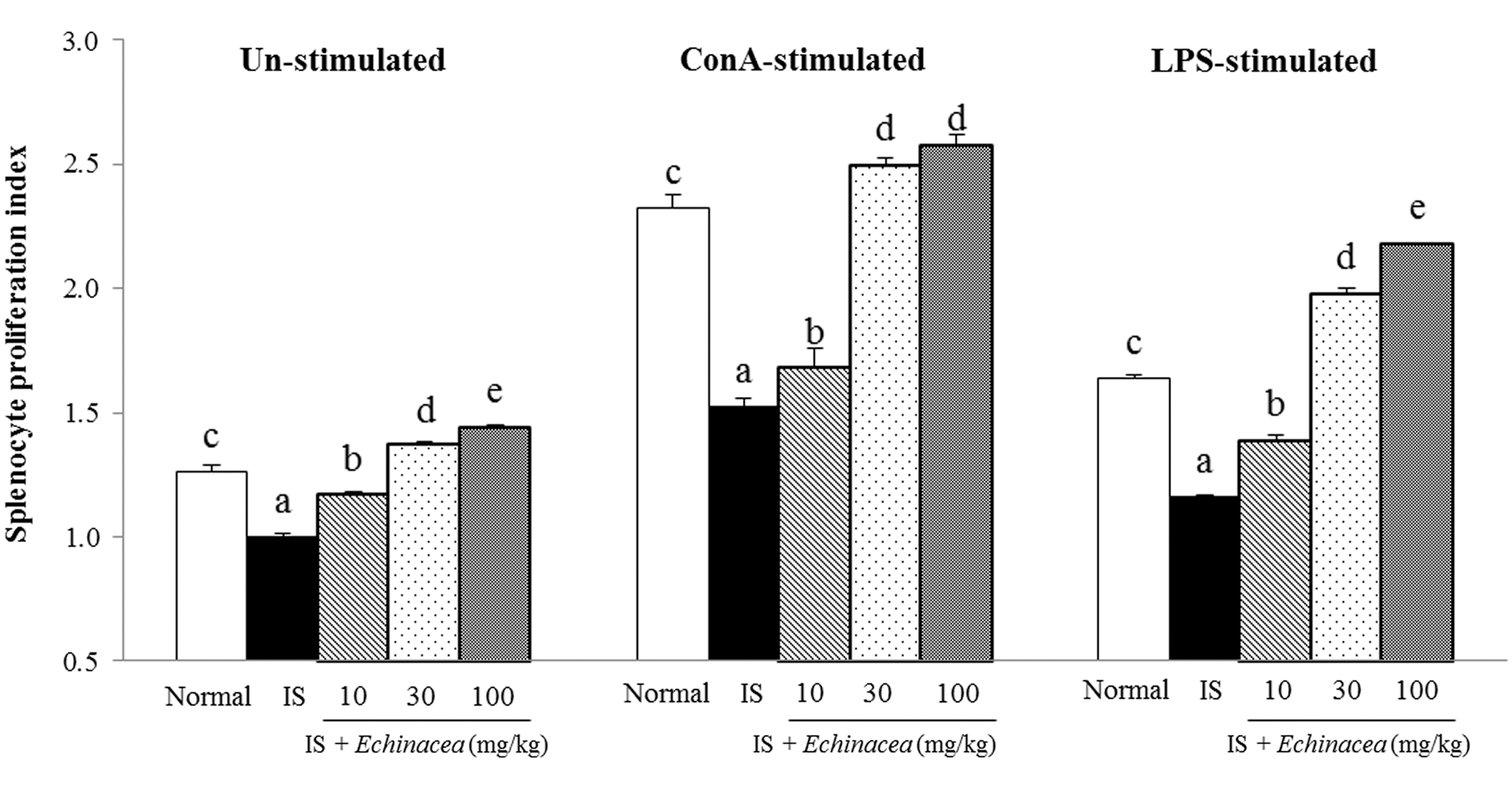
Effects of Echinacea on splenocyte proliferation index in restraint stress-induced immunosuppressed mice. Mice were orally administered with Echinacea EFLA®894 (10, 30, and 100 mg/kg) for 2 weeks. Splenocytes were either unstimulated or stimulated with ConA or LPS for 44 h. The results are presented as proliferation indices. Proliferation index = mean optimal density in test wells/mean optical density in IS control wells. Values are expressed as the mean ± standard error (n = 6). abcdeThe means without a common superscript letter are significantly different at the P < .05 level by Tukey's multiple range test. ConA, concanavalin A; LPS, lipopolysaccharide; IS, restraint-induced immunosuppression group control.
Splenic NK cell activity
NK cell activity in the IS group was significantly decreased by 40% compared to the normal group. However, Echinacea increased NK cell activity (linear effect, P < .01). In specific, mice treated with 30 and 100 mg/kg Echinacea showed an increase in NK cell activity by 17% and 20%, respectively, compared with the IS group (Fig. 2).

Effects of Echinacea on splenic NK cell activity in restraint stress-induced immunosuppressed mice. Mice were orally administered with Echinacea EFLA 894 (10, 30, and 100 mg/kg) for 2 weeks. Effector cells that target YAC-1 cells were mixed as a 10:1 ratio and incubated for 4 h. Values are expressed as the mean ± standard error (n = 6). abcThe means without a common superscript letter are significantly different at the P < .05 level by Tukey's multiple range test. IS, restraint-induced immunosuppression group control; NK, natural killer.
Blood T lymphocyte subsets
CD4+ and CD8+ T cells, which are markers for either helper (Th) or cytotoxic T (Tc) cells, were analyzed using flow cytometry. CD4+ and CD8+ T cells in blood were significantly decreased by 11.8% and 2.9% in the IS group compared to the normal group, respectively (Fig. 3). However, these levels were ameliorated by Echinacea at 30 mg/kg compared to those of the IS group.
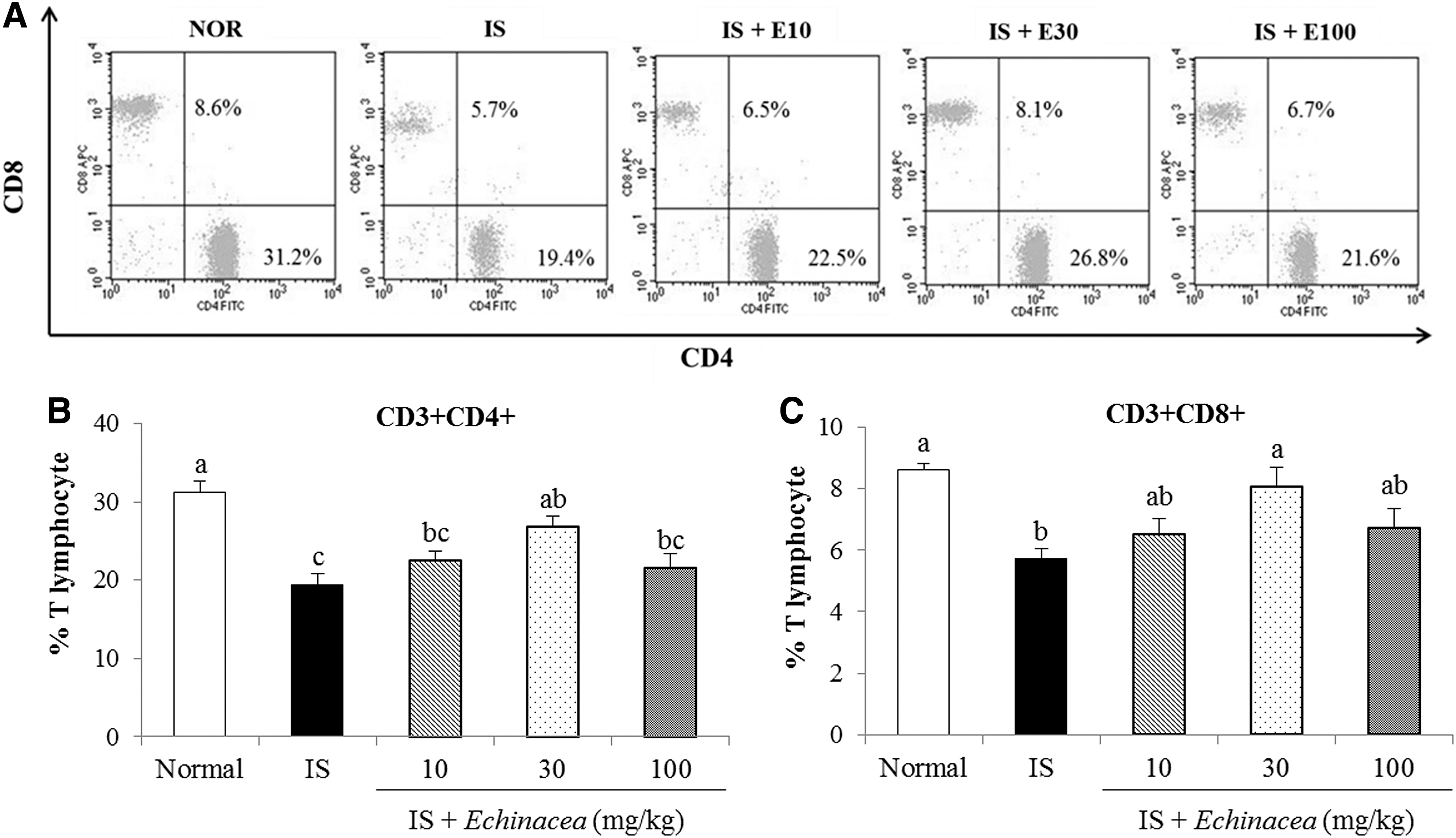
Effects of Echinacea on blood T cells
Serum cytokine production
As shown in Figure 4, the serum OPG level was significantly decreased by 45.5% in the IS group compared with the normal group. The OPG level in mice treated with Echinacea at 30 mg/kg significantly increased by 32.2% compared to the IS group.

Effects of Echinacea on serum levels of OPG in restraint stress-induced immunosuppressed mice. Mice were orally administered with Echinacea EFLA 894 (10, 30, and 100 mg/kg) for 2 weeks. Values are expressed as the mean ± standard error (n = 8). abcThe means without a common superscript letter are significantly different at the P < .05 level by Tukey's multiple range test. IS, restraint-induced immunosuppression group control; OPG, osteoprotegerin.
The serum IL-6, IL-10, and IL-17 levels were significantly higher in the IS group than in the normal group (Fig. 5). However, the IL-6, IL-10, and IL-17 cytokine levels were linearly reduced by Echinacea treatment (linear effect, P < .05, P < .01, and P < .01, respectively). Particularly, treatment with 100 mg/kg Echinacea significantly reduced all these cytokine levels. Also, the Echinacea 10 mg/kg group significantly decreased the levels of IL-6 and IL-10, while treatment with Echinacea 30 mg/kg reduced IL-10 and IL-17 levels compared with the IS group (P < .05).
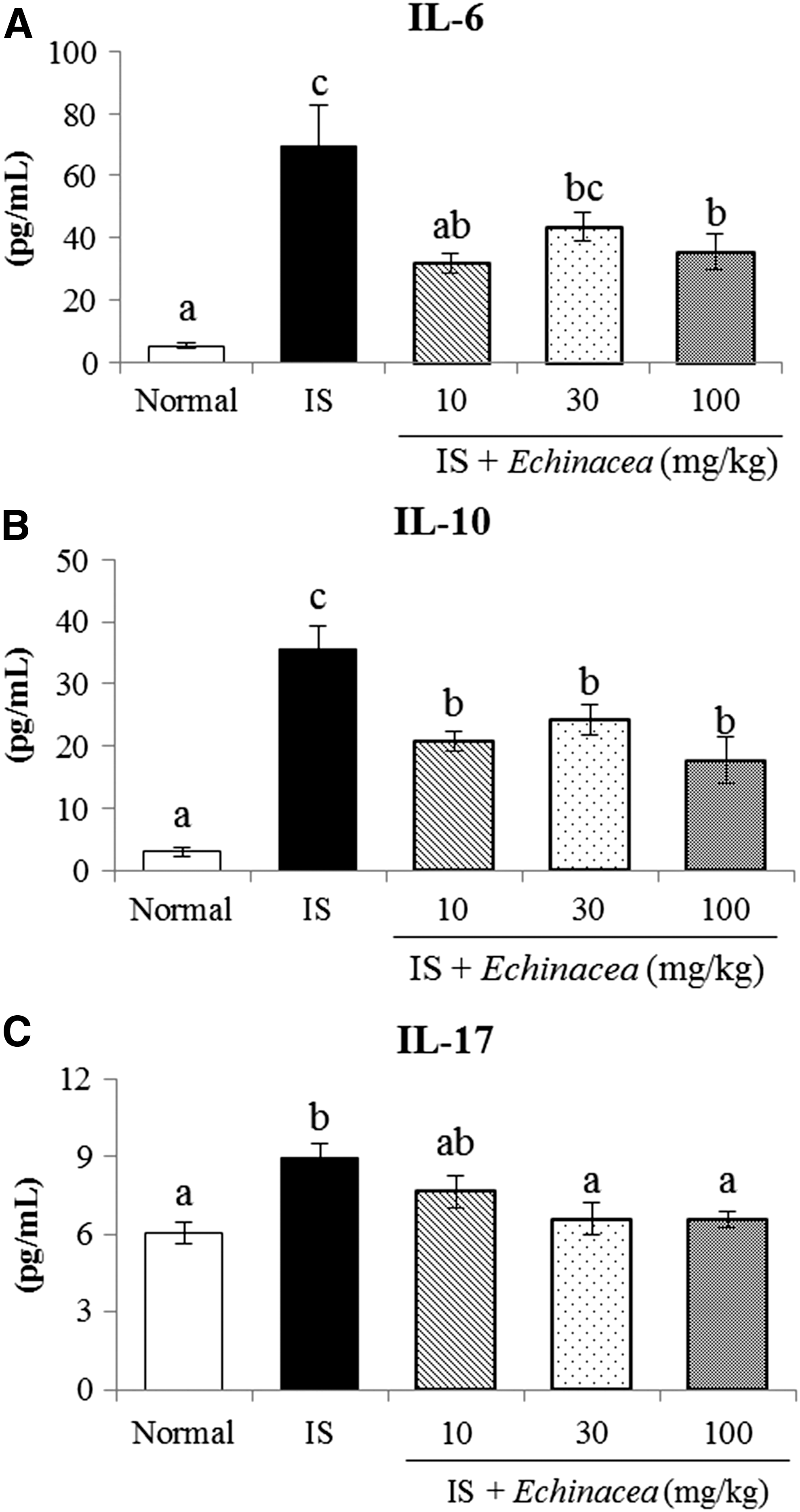
Effects of Echinacea on serum levels of IL-6
Cytokine mRNA expression
We next analyzed whether Echinacea treatment affects the mRNA expression of cytokines in spleen (Fig. 6). The mRNA levels of cytokines such as Il-6, Il-10, and Il-17 mRNA were significantly increased in the IS group compared to the normal group. However, treatment with Echinacea at 10, 30, and 100 mg/kg significantly downregulated mRNA levels of Il-6, Il-10, and Il-17a (linear effect, P < .01, P < .01, and P < .01, respectively).

Effects of Echinacea on spleen mRNA expression of Il-6
Discussion
Restraint-induced stress alters lymphocyte subsets and antibody production and suppresses the cytotoxicity of T and NK cells. 3,5,6 Echinacea improves immune function and reduces inflammation and oxidative stress. 20,21 However, the role of Echinacea in mice challenged with restraint stress remains unexplored. In this study, Echinacea enhanced the immune function by promoting splenocyte proliferation and NK cell activity. In addition, Echinacea modulated blood T cell distribution, serum levels, and mRNA expressions of cytokines such as IL-6, IL-10, and IL-17, which are responsible for inflammation and autoimmune diseases.
The spleen is responsible for differentiation and maturation of lymphocytes and induction of cellular and humoral immunity. 22 Cellular immune responses are mediated mainly by T cells, whereas the humoral immune response involves B cells. 23,24 T and B lymphocytes are known to be stimulated by mitogens and can be used as an immune parameter to investigate proliferative effects because of their high sensitivity. 25 In this study, spleen cell proliferative responses in both the absence and presence of a mitogen were suppressed by restraint stress, relative to those in the normal group. These observations are consistent with results from other studies in which restraint stress suppresses lymphocyte proliferation in the spleen. 2,9 Echinacea enhanced the ConA- and LPS-stimulated splenocyte proliferation in a dose-dependent manner in immunosuppressed mice. In addition, cells grown in the absence of mitogens showed a proliferative response indicating that Echinacea, itself, might act like a mitogen to promote splenocyte proliferation in mice with restraint stress. A previous study reported that orally administered Echinacea ethanol extract for 7 days increases T lymphocyte proliferation in response to ConA stimulation in BALB/c mice immunized with sheep erythrocytes. 21 Taken together, Echinacea might improve cellular and humoral immune responses by promoting T and B cell activations.
NK cells play key roles in innate immunity. 26 They kill pathogen-infected cells and tumor cells through direct cytotoxicity and the secretion of cytokines. 27 Previous studies have shown that the number and cytotoxic function of NK cells decrease in restraint stress-immunosuppressed mice, leading to increased vulnerability to infections or occurrence of tumors. 5,6 In this study, splenic NK cytotoxic activity decreased with restraint stress, relative to that in the normal group, whereas Echinacea restored NK cell activity in restrained mice. A previous study reported a significant increase in NK cell activity in sRBC-immunized mice with oral administration of Echinacea for 7 days. 21 Thus, Echinacea can be assumed to have beneficial effects on innate immunity by enhancing NK cell cytotoxicity in the spleen.
All T lymphocytes have CD3+ surface receptors that can be used to define the total T cell population; CD4+ and CD8+ are T cell surface markers for Th and Tc cells, respectively. 9 In this study, mice with restraint stress showed lower T lymphocyte counts than the normal group. These results are similar to those of previous reports, in which restraint stress markedly reduced the number of CD4+ and CD8+ cells. 9,28 Echinacea increased the T lymphocyte population (CD3+CD4+, and CD3+CD8+ T cells) in restrained mice. Similarly, a previous study showed that E. purpurea dried powder increased the number of CD4+ and CD8+ T cells in mouse peripheral blood. 20
OPG is a soluble protein that inhibits differentiation and activation of osteoclasts by binding to its ligand. 29 More than 50% of bone marrow-derived OPG is produced by mature B cells, and activated T cells promote OPG production by interacting with B cells. 30 Both B cell knockout mice and T cell-deficient nude mice showed decreased OPG secretion. 31 Accordingly, increased OPG levels might contribute to T cell and B cell activation. Our results showed that restraint stress significantly reduced the OPG levels in serum, but that Echinacea restored OPG levels. This result might suggest that T cells and B cells are inactivated in mice with restraint stress-induced immunosuppression and T and B cells are activated by Echinacea.
Altered stress-induced cytokines may be the connection between stress and infections, autoimmune diseases, and cancer. 7 Previous studies have shown that serum levels of IL-6, IL-10, and IL-17 are higher in restraint stress conditions compared to those in non-stress conditions. 7,8 IL-6 is secreted by various cells, such as T cells and macrophages, and is associated with immune disorders, inflammation, and lymphatic tumors. 32 When psychologically stressed, the body secretes stress hormones, such as cortisol, which can release IL-6 into circulation. 33 IL-17 is a proinflammatory cytokine that is secreted primarily by Th17 cells and induces autoimmune disease and tissue inflammation. 34 IL-6 induces differentiation of Th17 cells and production of IL-17 through the JAK2 and STAT3/ROR-γ pathways. 35,36 IL-10 is a suppressor cytokine secreted mostly by Th2 and regulatory T cells (Tregs). IL-10 controls T and B cell activities and has been implicated in the development of autoimmune disorders, allergies, and cancer. 37 Previous studies have shown that restraint stress-induced changes in T lymphocytes are highly associated with change in cytokine production. 9,38 In our study, higher levels of IL-6, IL-17, and IL-10 in serum were observed in restraint-stressed mice, and Echinacea decreased levels of those cytokines. Also, Il-6, Il-10, and Il-17 mRNA expression levels were upregulated in restraint-stressed mice and Echinacea-supplemented mice showed notable reduction of those cytokines. Therefore, Echinacea might alleviate inflammation and autoimmune diseases by restoring cytokine levels, including IL-6, IL-10, and IL-17, and normalizing its mRNA expression in the spleen. The effect on IL-17, in particular, is noteworthy due to the pivotal role of Th17 cells in pathogenesis of autoimmune diseases. 39
Polysaccharides and alkylamides are major bioactive substances of Echinacea, which have been reported to play pharmacological roles, such as immunostimulatory, anti-inflammatory, and antioxidant effects. 15 In this study, Echinacea dried pressed juice showed predominance of β-1,2-D-fructofuranosides and alkylamides. These compounds probably contributed to the beneficial effects of Echinacea in immunosuppressed mice. A previous study demonstrated that Echinacea pressed juice stimulates macrophage activity and cytokine synthesis and fructofuranosides may be responsible for these immune-enhancing effects. 17 Therefore, we suggest that β-1,2-D-fructofuranosides and alkylamides may partially explain the protective effect of Echinacea on immunosuppressed condition in this study. However, as this is an assumption, further investigation is needed to determine whether β-1,2-D-fructofuranosides and alkylamides are actually responsible for the favorable activity on the immune system.
Taken together, our study suggests that the pressed-juice powder of E. purpurea flowers enhances immune function in restraint stress-induced immunosuppressed mice. It may promote splenocyte proliferation, accelerate NK cell activity in the spleen, and regulate T lymphocyte subsets and cytokine levels in blood, leading to its beneficial effects. These findings further contribute and support to the evidence-based use of Echinacea as an immune-promoting herbal medicine.
Footnotes
Acknowledgments
This work was supported by the Frutarom, Switzerland; Novarex, Republic of Korea; and PRogram for Industrial needs - Matched Education (PRIME), Ewha Womans University funded by the Ministry of Education of Korea.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.