
Abstract
Parkinson's disease (PD) is characterized by the progressive degeneration of dopaminergic (DA) neurons in the substantia nigra pars compacta and decreases in striatal dopamine levels. These changes led to several clinical symptoms: rigidity, resting tremor, and bradykinesia. Although the cause of PD remains unclear, it is widely accepted that oxidative stress, neuroinflammation, mitochondrial dysfunction, and insufficient support of neurotrophic factors are involved in the pathophysiology of the disease. However, novel regimens to prevent neurodegeneration and restore the degenerated nigrostriatal DA system are still required. In recent years, there has been a growing interest in naturally occurring phytochemicals, which are believed to reduce the risk of neurodegenerative diseases. This review provides an overview of the scientific literature concerning the preventive and protective roles of flavonoids, one of the largest families of phytochemicals, against PD. In addition to providing antioxidant and anti-inflammatory effects, flavonoids exhibit a neuroprotective effect by activating antiapoptotic pathways that target mitochondrial dysfunction and induce neurotrophic factors. This review suggests that flavonoids may be promising natural products for the prevention of PD and could potentially be utilized as therapeutic compounds against PD, even though there was no report showing that the treatment with flavonoids could restore the aberrant phenotypes of patients with PD.
Introduction
P
Parkinson's Disease
Two key changes characterize PD: the progressive degeneration of dopaminergic (DA) neurons, leading to depletion of striatal dopamine, and the formation of Lewy bodies in the substantia nigra (SN). 6 The main clinical symptoms of PD include resting tremor, rigidity, bradykinesia, gait difficulty, and postural instability. 6 Several nonmotor symptoms, including depression, anxiety, emotional changes, cognitive impairment, sleep difficulty, and olfactory dysfunction, can also occur. 6 Although the precise pathogenic mechanism leading to neurodegeneration in PD is still not fully understood, the pathogenesis is thought to involve oxidative stress, mitochondrial dysfunction, neuroinflammation, and insufficient neurotrophic factor support. 7,8
Oxidative stress occurs as a result of an imbalance between reactive oxygen species (ROS) generation and cellular antioxidant activity. In the PD brain, excess ROS can be generated during dopamine metabolism and exacerbated by low levels of glutathione and high levels of iron and calcium in the SN pars compacta (SNc). 9 Moreover, the brain contains high levels of polyunsaturated fatty acids, which are the major substrates in the process of lipid peroxidation under oxidative stress. 10 DA neurons are particularly prone to oxidative stress because of the presence of ROS-generating enzymes, such as tyrosine hydroxylase (TH) and monoamine oxidase. 11
Mitochondria are involved in various cellular processes, including energy generation, calcium homeostasis, stress response, and cell death pathways. Impaired mitochondrial function can cause excessive formation of ROS and cellular damage, which is linked to neurodegeneration. 11,12 Neurons are highly dependent on mitochondrial ATP production, and hydrogen peroxide (H2O2) and superoxide radicals are normally generated during oxidative phosphorylation as byproducts from the mitochondria. 13 Any environmental or genetic factors leading to the impairment of mitochondrial function can induce dramatic increases in ROS, which then overwhelm the cellular antioxidant defense systems, leading to cell damage. 14
Neuronal loss in PD is also related to neuroinflammation, which is mainly controlled by microglia. Microglia are glial cells that act as the first and main form of active immune defense in the central nervous system. 11 In both idiopathic and genetic PD patients, microglial activation has been observed. 15,16 The activated microglia can produce free radicals, leading to oxidative stress. This stress is aggravated by inflammatory signals generated by the molecules released from damaged DA neurons such as α-synuclein and matrix metalloproteinase-3. These molecules can contribute to the induction of reactive microgliosis. 11 Moreover, activated microglia produce lysosomal cysteine proteases such as cathepsin B, which is involved in microglial production of pro-inflammatory cytokines and induces neuronal death under various pathological conditions. 17
It has been suggested that neurotrophic factors, such as glial cell line-derived neurotrophic factor (GDNF) and brain-derived neurotrophic factor (BDNF), are indispensable for the survival and protection of DA neurons. In the SN of a patient with PD, the distribution of GDNF and BDNF is decreased. 18 Moreover, in animal models of PD, these factors have been shown to produce neuroprotective effects. 19 N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) causes permanent Parkinsonian-like symptoms by destroying DA neurons in the SN of the brain. In MPTP mouse models of PD, a direct injection of GDNF into the SN or striatum led to an increased density of DA fibers and improved the MPTP induced motor impairments. In contrast, conditional GDNF-null adult mice displayed a delayed and progressive loss of DA neurons. 19 In neurotoxic models of PD, treatment with specific neutralizing antibodies for BDNF leads to anatomical, neurochemical, and behavioral deficits. This indicates that decreased expression of BDNF may contribute to DA neuronal degeneration. 19 However, the usage of these factors to treat PD is limited as they do not penetrate the blood–brain barrier (BBB). Therefore, it is difficult to target GDNF and BDNF specifically in the DA neurons without several side effects. 20,21 Taken together, this shows that a strategy for protecting against oxidative stress, mitochondrial dysfunction, and neuroinflammation, as well as the replacement of neurotrophic factors, could protect against DA neurodegeneration in PD.
Flavonoids
Flavonoids are one of the largest families of plant-derived phenolic compounds and are widely found in foods and beverages of plant origin. 22 Chemically, flavonoids are based on a 15-carbon skeleton consisting of 2 benzene rings (A and B rings) connected by a heterocyclic pyrone ring (C ring) (Fig. 1). They are divided into six major subclasses (flavones, flavonols, flavanones, flavanols, isoflavones, and anthocyanins) depending on which carbon of the C ring the B ring is attached to and the degree of oxidation and substitution of the C ring. 22

Most flavonoids are ingested in the form of glycosides, which are bound to one or more sugar molecules. 22 After ingestion of flavonoid-containing foods, the glycosides are hydrolyzed by intestinal hydrolases or gut microflora. 22 The released aglycones are absorbed by the intestinal cells and converted into their respective metabolites in intestinal and hepatic cells. 22 Finally, the metabolites are transported by the blood to various cells and or excreted in the urine. 22 In general, the bioavailability of flavonoids is low due to poor absorption, rapid excretion, and the extensive biotransformation and conjugation that occur during their absorption from the intestine, in the liver and in cells. 22
Antiparkinsonian Effects of Flavonoids
Despite their low bioavailability, extensive studies have reported that flavonoids can provide numerous health benefits, including a reduction in the risk of PD. This is due to their biologic effects, which include antioxidative, anti-inflammatory, antiapoptotic, and lipid-lowering properties. 23
Flavones
Baicalein
In in vitro studies, baicalein, a flavone that occurs in the roots of Scutellaria baicalensis, inhibited the formation of α-synuclein oligomers and consequently prevented their fibrillation. 24,25 In neurodegenerative diseases, including PD, α-synuclein, a presynaptic neuronal protein, is aberrantly folded and forms an abnormal oligomer, protofibril, or amyloid fibril that induces synaptic disturbance and neuronal death. 26 Furthermore, secreted α-synuclein may be transferred to neighboring cells, accelerating the aggregation process, thus contributing to disease propagation. 26 Therefore, α-synuclein–targeting therapy has emerged as a promising approach for the treatment of PD. 27 Jiang et al. 28 also reported that baicalein protected cells against toxicity induced by E46K (a point mutation in α-synuclein, which causes familial Parkinsonism with Lewy body dementia) in cellular models of PD. In a more recent study performed on a PD mouse model generated by the intragastric administration of rotenone, a classical PD causing toxin, baicalein prevented the progression of α-synuclein accumulation and protected DA neurons against α-synuclein oligomer-induced toxicity. The study also found that baicalein inhibited the formation of α-synuclein oligomers. 29 Baicalein has been consistently shown to inhibit elevation in α-synuclein aggregates, as well as autophagy in rats treated with 1-methyl-4-phenylpyridinium (MPP+), a neurotoxin that has been used to induce PD in animal models. 30 Abnormal autophagy is observed in the brains of animal models and patients with PD and is proposed to be responsible for the aggregation of toxic proteins. 31
In addition to its beneficial effects on α-synuclein, numerous studies have reported neuroprotective effects of baicalein through anti-inflammatory, antioxidant, and antiapoptotic actions. In MPTP-treated mice, baicalein prevented abnormal behavior by increasing DA neurons, as well as dopamine and serotonin levels in the striatum. It also inhibited oxidative stress and the astroglial response. 32 Lee et al. 33 reported that baicalein attenuated MPTP-induced astroglial activation in mice by downregulating the activation of nuclear factor-κB (NF-κB), extracellular signal–regulated kinase (ERK), and c-Jun N-terminal kinase (JNK). In PD, the NF-κB pathway is activated in various cell types, including astrocytes, and plays an important role in the activation and regulation of inflammatory mediator production. 34 Stress response kinases, such as ERK and JNK, contribute to astrocyte activation. Activated astrocytes then release pro-inflammatory cytokines, which can exacerbate DA neuron degeneration in PD, leading to the alteration of DA control of striatal glutamatergic transmission. 35,36 Baicalein inhibited the upregulation of pro-inflammatory cytokines in the SN and striatum of a PD mouse model. 37 The neuroprotective efficacy of baicalein has been shown to be related to its ability to attenuate the increased striatal basal glutamatergic strength observed in PD, by inhibiting presynaptic glutamate release and the reassembly of postsynaptic glutamate receptor subunits. 37 Other studies using the parkinsonism–inducing neurotoxin 6-hydroxydopamine (6-OHDA) PD model in vivo and in vitro have demonstrated the protective effects of baicalein on DA dysfunction by its antioxidative, antiapoptotic, and anti-inflammatory actions. 38 –40
Luteolin and apigenin
Luteolin and apigenin are flavones that are similar in structure, with only one-hydroxyl difference. Luteolin exists in many edible plants, including celery, broccoli, parsley, thyme, and olive oil. 41 Apigenin is also present in many fruits, vegetables, and herbs. 42 In an in vitro study, luteolin and apigenin were shown to modulate microglial activation by inhibiting inflammatory mediators, suggesting that they may possess neuroprotective effects. 43 In another in vitro study, luteolin protected DA neurons against inflammation-induced neurotoxicity by inhibiting microglial activation. 44 Luteolin also reduced ROS production and cytotoxicity induced by 6-OHDA in neuronal PC12 cells, by modulating the expression of ROS-dependent stress response genes. 45
Several in vivo studies have supported the potential for these compounds to improve PD symptoms. 46 –49 In MPTP-treated mice, luteolin and apigenin improved locomotor and muscular activity. They also protected DA neurons by reducing oxidative damage, neuroinflammation, and microglial activation and enhancing neurotrophic effects. 46 In a rotenone-induced rat model of PD, apigenin ameliorated behavioral impairments, as well as DA neuronal loss. These effects were associated with the suppression of neuroinflammation and oxidative stress-mediated apoptosis. 49 Apigenin also decreased α-synuclein aggregation and modulated DA neurotransmission, by increasing dopamine biosynthesis and dopamine D2 receptor expression. 49
Nobiletin and tangeretin
Nobiletin, a methoxylated flavone found in citrus fruits, can inhibit microglial activation and activate intracellular prosurvival pathways, which are involved in neuronal survival and protection. 50,51 In MPTP-treated mice, nobiletin (25 and 50 mg/kg) rescued motor and cognitive dysfunction by enhancing dopamine release in the striatum and hippocampal CA1. However, it did not ameliorate DA neuron loss in the midbrain. 52 Similarly, a daily intraperitoneal injection of nobiletin (20 mg/kg) did not protect DA neurons against MPP+-induced neurotoxicity. 53 However, at 10 mg/kg, nobiletin protected DA neurons and attenuated interleukin-1β production by inhibiting microglial activation and preserving GDNF expression in the SN. This suggests that a higher dosage of nobiletin can provide protective effects against DA neuron degeneration. 53 Another methoxylated flavone, tangeretin, was reported to cross the BBB and maintain striatonigral integrity and functionality in an in vivo 6-OHDA model of PD. 54
Flavonols
Quercetin, isoquercitrin, rutin, and troxerutin
Quercetin is a flavonol widely distributed in fruits, vegetables, and grains. It is the aglycone form of other flavonoid glycosides, such as rutin and quercitrin. 55 A study assessing the tissue distribution of quercetin has shown that quercetin and its metabolites become widely distributed in rat tissues, including the brain. 56 There is also evidence that quercetin can accumulate in the central nervous system following oral administration. 57 –59 Moreover, quercetin can travel across the BBB. 60
In the 6-OHDA–induced PD rat model, the application of quercetin led to increased striatal dopamine, neuronal survivability, and antioxidant enzyme levels, suggesting that it may convey neuroprotective effects. 61 Quercetin has consistently been shown to protect against oxidative stress and dopamine depletion, improve motor balance and coordination, and maintain the resting membrane potential of neurons in MPTP-treated mice. 62 Moreover, in a rotenone model of PD, quercetin has been shown to attenuate the loss of mitochondrial complex-I activity and striatal dopamine. Quercetin also led to increases in endogenous antioxidant enzyme activities. 63 Recently, Ay et al. 64 demonstrated that quercetin led to upregulated phosphorylation of protein kinase D1 (PKD1), Akt, cAMP response-element binding protein (CREB), and BDNF (a CREB target gene) expression in DA neuronal cells. PKD1 and Akt mediate neuronal survival-promoting activities, and PKD1 directly phosphorylates CREB. 65 –68 Moreover, quercetin improved mitochondrial biogenesis and bioenergetic capacity in DA neurons and reversed nigrostriatal degeneration, striatal dopamine depletion, and behavioral deficits in MitoPark mice. 64
Isoquercitrin and rutin, the glycosides of quercetin, have also been shown to protect against 6-OHDA–induced neurotoxicity in vitro and in vivo. 69 –71 Moreover, neuroprotective effects of troxerutin, a derivative of rutin, were reported in a 6-OHDA lesion rat model of PD. 72 In this model, pretreatment with troxerutin improved apomorphine-induced rotational behavior bias and decreased both the latency to initiate and the total time on the narrow beam task. Central administration of an ERβ antagonist or PI3K inhibitor abrogated these beneficial effects. 72 Moreover, troxerutin prevented the loss of nigral TH-positive neurons and decreased striatal lipid peroxidation, ROS, astrogliosis, and apoptosis. Therefore, by ameliorating apoptosis, astrogliosis, and oxidative stress, troxerutin could be a potent neuroprotective agent against PD. The beneficial effects of troxerutin may be partly reliant on PI3K/ERβ signaling. 72
Kaempferol
Previous studies have reported neuroprotective effects of the flavonol kaempferol against several apoptosis and necrosis-inducing insults, such as the application of oxidized low-density lipoproteins and L-glutamate. 73 –76 In MPTP-treated mice, kaempferol induced antioxidative activity, leading to improved motor function and increased levels of striatal dopamine and its metabolites. 77 Kaempferol protected against acute rotenone-induced toxicity and preserved the striatal glutamatergic response in the rat brain. 78 Its derivatives also led to decreased H2O2-induced ROS levels and cell death and restored TH activity in a protein deglycase DJ-1-dependent manner in vitro. 79
Myricitrin and myricetin
The flavonol myricitrin, derived from the root bark of Myrica cerifera, and its aglycone myricetin have been shown to possess neuroprotective capabilities in vitro and in vivo. 80 –85 In PC12 cells, myricitrin ameliorated 6-OHDA–induced mitochondrial damage and apoptosis by inhibiting mitochondrial oxidation. 80 Myricitrin also attenuated MPP+-induced mitochondrial dysfunction in DA SN4741 cells. 81 Interestingly, its protective efficacy was lost when there was a deficiency of DJ-1. DJ-1 is a familial recessive PD gene, involved in antioxidant function and mitochondrial maintenance. This indicates that myricitrin can alleviate mitochondrial dysfunction through DJ-1 activity. 81 In mice that received an intrastriatal injection of 6-OHDA, myricitrin protected DA neurons against neurotoxicity. The protective effects were associated with the activation of mammalian target of rapamycin complex 1 (mTORC1), preservation of TH activity, and anti-inflammatory action. 82 It is suggested that mTORC1 activation is required to protect DA neurons in animal models of PD, as it leads to the production of neurotrophic factors that are important for preserving nigrostriatal DA projections. 19,86,87 TH, an enzyme involved in dopamine production, is also important for sustaining DA activity in the brain. 88
Akin to myricitrin, myricetin prevented the decrease of TH and DA neuron degeneration induced by 6-OHDA in rats. 83 In vitro studies have also shown its neuroprotective effects. 84,85 In DA-like MES23.5 cells, myricetin attenuated MPP+-induced cell loss and nuclear condensation, by suppressing intracellular ROS production and inhibiting the activation of mitogen-activated protein kinase kinase 4 (MKK4) and JNK. 84 Aberrant activation of the MKK4-JNK pathway is known to cause death of SNc DA neurons. 89 Myricetin also decreased methylmercury-induced mouse brain mitochondrial dysfunction and oxidative stress in vitro. 85
Flavanones
Naringin and naringenin
Naringin is a major flavanone glycoside found in citrus fruits, especially in grapefruits. 90 Numerous studies have suggested that naringin may be a potential therapeutic agent against neurodegenerative diseases. 91 –93 In rotenone-treated human neuroblastoma SH-SY5Y cells, a commonly used cellular PD model, the compound exerted antiapoptotic effects. 91 In rats, naringin protected nigrostriatal DA projections from MPP+-induced neurotoxicity through anti-inflammatory activity and GDNF induction. 92 In another in vivo model of PD, pretreatment with naringin exerted anti-inflammatory effects in the mouse brain and activated mTORC1 in DA neurons. This protected nigrostriatal DA projections from 6-OHDA–induced neurotoxicity. 93 However, posttreatment with naringin did not restore the nigrostriatal DA projections in 6-OHDA–treated mice, despite the treatment being continued for 12 weeks. 93
The aglycone of naringen, naringenin, can readily pass through BBB and has been shown to provide neuroprotection in experimental models of PD. 94 –98 In 6-OHDA treated rats, pretreatment with naringenin protected the rats from loss of TH-positive cells in the SN, as well as the decreased dopamine content in the striata. 94 Lou et al. 95 reported that pretreatment with naringenin protected against 6-OHDA–induced neurotoxicity in vitro and in vivo. In SH-SY5Y cells, naringenin ameliorated 6-OHDA–induced neurotoxicity through upregulation of nuclear factor E2-related factor 2 (Nrf2) and subsequent activation of the antioxidant response element (ARE) pathway. 95 Nrf2 is known to activate the ARE pathway, which includes various cytoprotective genes (including antioxidant and anti-inflammatory genes) and several transcription factors involved in mitochondrial biogenesis. Therefore, the Nrf2/ARE pathway is implicated to play a role in protection against neurodegenerative conditions. 99 An in vivo study using 6-OHDA–lesioned mice also demonstrated that by activating the Nrf2/ARE signaling pathway, naringenin attenuated nigrostriatal DA neurodegeneration and oxidative damage. 95 In mice, the compound has been shown to lead to improved motor coordination function and reduced microglial activation. These effects were related to the inhibition of neuroinflammatory responses by the activation of suppressor of cytokine signaling 3 expression. 96
Hesperidin and hesperetin
Hesperidin is another flavanone glycoside found in citrus fruits and other plants. 100 Along with antioxidant, anti-inflammatory, anticarcinogenic, and lipid-lowering effects, hesperidin and its aglycone hesperetin are reported to have neuroprotective effects. 101 –106 In an in vitro study, hesperetin protected neurons against various types of insults associated with neurodegenerative disorders, although there was minimal or no protective effects against neuronal damage. 101 However, using a rotenone-induced cellular model of PD, Tamilselvam et al. 102 demonstrated that hesperidin exerted a neuroprotective effect by its antioxidative and antiapoptotic properties, as well as by maintenance of mitochondrial function.
The beneficial effects of hesperidin and hesperetin in PD have been observed in in vivo studies. 103 –106 In 6-OHDA–treated aged mice, hesperidin improved behavioral alterations and protected the striatum against oxidative stress. Increased levels of dopamine and its metabolites were also observed. 103 The compound has been shown to ameliorate MPTP-induced motor dysfunction and neurotoxicity through its antioxidant and anti-inflammatory activities. 104 In addition, hesperetin improved rotational behavior in unilateral intrastriatal 6-OHDA–lesioned rats. 105 The beneficial effects of hesperetin against 6-OHDA–induced behavioral abnormalities are supported by a recent study which demonstrated that oral administration of hesperetin protected against unilateral striatal 6-OHDA damage in rats through the attenuation of apoptosis, astrogliosis, and oxidative stress. 106
Flavanols
Epigallocatechin-3-gallate
Epigallocatechin-3-gallate (EGCG) is a major flavanol in tea. It has been shown to display neuroprotective effects in numerous models of neurotoxicity. 107 –119 In cultured rat cerebellar granule neurons, EGCG selectively protected against neuronal apoptosis triggered by mitochondrial oxidative stress. 107 Moreover, by upregulating the PI3K/Akt and glycogen synthase kinase-3 (GSK-3) pathways and downregulating mitochondrial damage, caspase-3, and poly (ADP-ribose) polymerase (PARP), EGCG diminished H2O2 induced oxidative stress and neuronal death in motor neurons. 108 The PI3K/Akt pathway is an important pathway involved in cell survival, by either transcriptional regulation or direct phosphorylation. 120 GSK-3 is a key downstream target of the PI3K/Akt pathway. It promotes mitochondria-mediated apoptosis by downstream signaling leading to mitochondrial damage and the activation of caspase-3 accompanied by PARP cleavage. 121 Levites et al. 109 also demonstrated a neuroprotective action of EGCG against 6-OHDA in human neuroblastoma SH-SY5Y cells, through the stimulation of protein kinase C (PKC) and modulation of cell survival/cell cycle genes. PKC is implicated in regulation of cell survival and programmed cell death. 122 The loss of neuronal PKC activity contributes to brain damage, while activation of PKC in neurons enhances memory, cognition, and learning and can confer antidementia actions. 123,124 In addition, EGCG inhibited α-synuclein aggregation and β-amyloid fibrillogenesis, 110,111 which are linked to the PD pathology. 112 Bieschke et al. 113 demonstrated that EGCG can bind to β-sheet–rich amyloid fibrils and oligomers and directly alter their morphology.
In addition to the neuroprotective effects of EGCG observed in vitro, EGCG protected against striatal dopamine depletion and SN DA neuron loss in an MPTP mouse model of PD. 114 Kim et al. 115 demonstrated that EGCG inhibited MPTP-induced activation of the neurotoxic molecule, inducible nitric oxide synthase, in both the SN and striatum. It also attenuated neuronal death rate in MPTP-treated mice. Pan et al. 116 suggested that the catechol-like structure of catechins might competitively inhibit dopamine uptake by transporters of MPP+ (metabolite product of MPTP) as these also possess a catechol-like structure, thereby protecting DA neurons against the MPP+-induced injury. The combination of EGCG and rasagiline, an anti-Parkinson monoamine oxidase B inhibitor used as monotherapy to treat symptoms in PD, synergistically restored dopamine neurons in MPTP-treated mice. This effect was due to several different mechanisms: activity at the MPTP-injured molecules/targets in the SN; rasagiline mediated induction of BDNF; EGCG induced increases in membrane levels of the PKC alpha-isoform; and a synergistic replenishment of the downstream effector, serine/threonine kinase Akt/protein kinase B. 117 Other studies using a 6-OHDA rat model of PD reported that EGCG can protect DA neurons by preventing 6-OHDA–induced ROS increases. 118,119
Isoflavones
Daidzein and genistein
Daidzein and genistein are two major isoflavones present in soybeans. 125 Akin to other isoflavones, they exert both estrogenic and antiestrogenic effects. Neuroprotective benefits have been reported. 126 –134 In murine microglial cells stimulated with lipopolysaccharide, daidzein exhibited neuroprotective effects by inhibiting microglial activation and the subsequent release of pro-inflammatory factors. 127 Its metabolite equol also inhibited microglial activation and exerted neuroprotective effects in vitro. 128
In rat mesencephalic neuron-glia cultures, genistein has been shown to protect DA neurons against lipopolysaccharide-induced neurotoxicity by inhibiting microglial activation. 129 In a hemi-parkinsonian rat model, genistein improved rotational behavior and protected neurons of the SNc against 6-OHDA toxicity. 130 Similarly, genistein protected DA neurons and preserved motor dysfunction caused by 6-OHDA in ovariectomized rats. This suggests that it may be useful for the prevention of PD in postmenopausal women. 131 Moreover, genistein improved spatial learning and memory in 6-OHDA–treated ovariectomized rats. 132 The neuroprotective effects of genistein on DA neurons were also observed in MPTP-treated mice. Genistein was associated with a reduction in the MPTP-induced cell survival factor, Bcl-2, in the mouse midbrain. 133 Like genistein, estrogen protected against MPTP-induced neuronal damage by enhancing Bcl-2 expression. This suggests that the mechanism underlying the neuroprotective action of genistein may be attributed to its estrogenic characteristics. 133 Arbabi et al. 134 also suggested that genistein substituted for estrogen in ovariectomized rats and prevented 6-OHDA–induced neuronal loss.
Anthocyanins
In SH-SY5Y cells, an anthocyanin-enriched extract protected the cells from 6-OHDA–induced neurotoxicity by antioxidative and antiapoptotic mechanisms. 135 Anthocyanin-induced neuroprotection against MPP+ and 6-OHDA was observed in primary DA neurons isolated from embryonic rats. 135 The compound also prevented MPTP-induced DA neuronal damage and PD-like symptoms such as bradykinesia by its antiapoptotic effects. 135 Interestingly, higher intake of anthocyanins and anthocyanin-rich dietary sources is related to a lower risk of PD in humans. 136,137
Pelargonidin, one of anthocyanidins found in red geraniums, protected against neurotoxicity induced by 5-S-cysteinyl-dopamine. 138 Pelargonidin also ameliorated motor deficits and nigral DA cell death in the 6-OHDA rat model of hemi-parkinsonism by partially inhibiting oxidative stress. 139
Limited Effects of Naringin for Neurorestoration Against Pd
Although the fundamental causes of PD remain unclear and no therapeutic agents restore the loss of DA neurons in PD, one treatment area that has gained significant momentum is the reconstruction of the nigrostriatal DA projections, which could be induced by the sustained activation of the mTORC1 signaling pathway, from damaged DA neurons in patients with PD. 86,87 However, a long-standing belief in neuroscience has been that the adult mammalian central nervous system is incapable of producing an axonal regenerative response, unlike the peripheral nervous system in which axons are able to regrow and restore function. 86 Recently we reported that mTORC1 activation in DA neurons by a specific gene delivery could reconstruct the nigrostriatal DA projection in a mouse model of PD, even though the gene delivery was performed after disruption of the DA system in the animal brain in vivo. 86,87 These results suggest that induction of mTORC1 activation may be necessary for axonal regeneration in the lesioned adult brain in vivo, and it may be useful as a therapeutic strategy to treat patients with neurodegenerative diseases such as PD.
In the previous study, we showed that naringin treatment induced an increase in mTORC1 activity in DA neurons in the SN in vivo, which consequently resulted in protection of the nigrostriatal DA projections in the animal models of PD 92,93 and that its activation could be preserved in the SN lesioned by a neurotoxin used in a mouse model of PD. 93 Although there was no report of natural/chemical compounds, which are capable of inducing axonal regeneration in vivo, these results raise the possibility that treatment with naringin may have the potential to regenerate new axons from lesioned DA neurons, similar to the previous results obtained from a specific gene transduction in DA neurons in vivo. 86,87 Unfortunately, however, our observations showed that posttreatment with naringin induced no significant changes in DA phenotypes in both the SN and striatum compared with 6-OHDA alone. In addition, no changes were found in the number of phospho-TH-positive neurons or the levels of phospho-TH, which were the indicators of functional activity of DA neurons in the SN, despite administration of naringin for 12 weeks. These results suggest that treatment with naringin alone may not be enough to restore the lesioned DA system, even though its administration can be used to protect the nigrostriatal DA projections involved in PD. Therefore, to elucidate the potential use of naringin or many various flavonoids in the treatment of patients with PD, further study is needed to determine the posttreatment effects of those, such as changes in the phenotypes of axonal regeneration and dopamine metabolic activity after damage to the DA system of adult brains.
Conclusion
Substantial evidence supports the proposal that naturally occurring flavonoids have the potential to be an effective alternative approach for the prevention and treatment of PD. Although precise mechanisms are still unclear, flavonoids regulate a number of important physiological responses, which may contribute to neuroprotective effects in PD. These responses led to decreased DA neuronal loss and dopamine depletion, reduction of neuroinflammation, improved antioxidant status and mitochondrial dysfunction, activated antiapoptotic pathways, induction of neurotrophic factors, and the inhibition of α-synuclein aggregation (Table 1 and Fig. 2). However, further long-term dietary intervention studies are required to fully evaluate the effectiveness of flavonoids as agents for the management of PD.
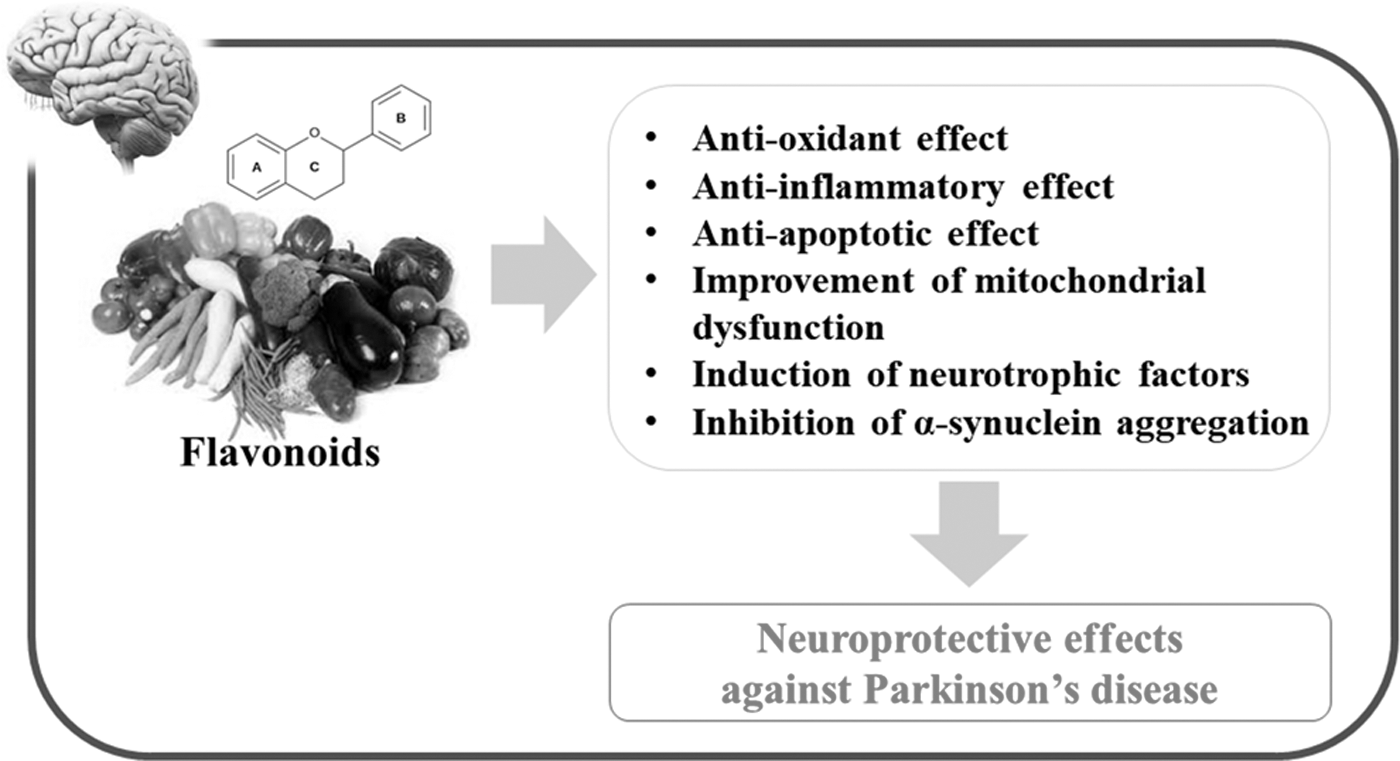
Schematic representation of the beneficial effects of flavonoids against PD. Many kinds of flavonoids can induce antioxidant, anti-inflammatory, and antiapoptotic effects, resulting in neuroprotection in the SN of adult brain. In addition to these beneficial effects, some flavonoids can induce the improvement of mitochondrial dysfunction, induction of neurotrophic factors, and inhibition of α-synuclein aggregation, which may intensify the neuroprotective effects. PD, Parkinson's disease; SN, substantia nigra.
DA, dopaminergic; EGCG, epigallocatechin-3-gallate.
Footnotes
Acknowledgments
This work was supported by grants from the Korea Healthcare Technology R&D Project, Ministry of Health & Welfare (HI15C1928), and the National Research Foundation of Korea (NRF-2014R1A1A4A01007858, NRF-2016R1D1A3B03931424, and NRF-2017R1A2B4002675).
Author Disclosure Statement
No competing financial interests exist.