
Abstract
Epimedium brevicornu Maxim has been used as a traditional herbal drug in China. In this study, the anti-inflammatory effects of E. brevicornu Maxim ethanol extract (EBME) were investigated in RAW264.7 macrophages and mice challenged with lipopolysaccharide (LPS). Results showed that EBME attenuated inflammation by decreasing the production of several proinflammatory mediators, such as nitric oxide (NO), prostaglandin (PG) E2, inducible nitric oxide synthase, and cyclooxygenase-2, in LPS-stimulated RAW264.7 macrophages. EBME increased the expression of heme oxygenase-1 (HO-1) and promoted the nuclear translocation of nuclear factor erythroid 2-related factor 2. The inhibitory effects of EBME on LPS-stimulated NO and PGE2 expression were partially reversed by HO-1 inhibitor. EBME also elicited an anti-inflammatory effect by inhibiting the production of tumor necrosis factor-α, interleukin (IL)-1β, and IL-6 in LPS-induced peritonitis. Therefore, EBME exhibited anti-inflammatory effects in vitro and in vivo.
Introduction
I
Herba Epimedii, the dried aerial part of Epimedium brevicornu, 4 is a frequently used herb in Chinese formulations to treat various diseases, such as osteoporosis and sexual dysfunction. 5 –7 In addition, it has been used as an ingredient of healthy alcoholic and tea beverages in China for its beneficial effects on the cardiovascular, reproductive, immune, skeletal, and nervous systems. 8 This herb also exhibits several biological activities, including inducing estrogenic 9 activity and peripheral nerve regeneration. 10 However, the anti-inflammatory effects of Herba Epimedii have not yet been fully described. To further elucidate the biological activities of Epimedium, we investigated the anti-inflammatory effects of the ethanol extract of E. brevicornu Maxim, a major Herba Epimedii species. 7
Heme oxygenase (HO)-1 is a rate-limiting enzyme that catalyzes the degradation of heme to free iron, biliverdin, and carbon monoxide. 11 HO-1 and its enzymatic byproducts act as important cytoprotective factors in counteracting the detrimental effect of inflammation by attenuating the expression of proinflammatory mediators, such as nitric oxide (NO) and prostaglandin (PG) E2. 12,13 Nuclear factor erythroid 2-related factor 2 (Nrf2) plays an important role in modulating the expression of HO-1. Under normal conditions, Nrf2 binds to Kelch-like ECH-associated protein 1 (Keap1) in the cytoplasm, 14 and the transcription repressor Bach1 suppresses HO-1 gene transcription by binding to an antioxidant response element (ARE) in the HO-1 promoter. 15 Under stressful conditions, Nrf2 becomes released from Keap1 and translocates to the nucleus. 14 Bach1 then dissociates from the HO-1 promoter, and Nrf2 activates HO-1 gene transcription by binding to ARE. 15 An increasing number of therapeutic agents, such as diallyl disulfide, 16 benzaldehyde, 17 and isovitexin, 18 induce HO-1 expression and exert anti-inflammatory effects through the Nrf2/HO-1 pathway.
So far, the anti-inflammatory effect and mechanisms related to inflammation of Epimedium ethanol extract is unclear. In this study, the potential anti-inflammatory activities of E. brevicornu Maxim ethanol extract (EBME) were evaluated in lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages. To investigate the possible anti-inflammatory mechanism of EBME, we examined whether EBME could activate the Nrf2/HO-1 signaling pathway and analyzed the anti-inflammatory activity of orally administered EBME in vivo.
Materials and Methods
Chemicals and reagents
The following chemicals and reagents were used: Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), and other tissue culture reagents (Invitrogen, Carlsbad, CA, USA); the Enzyme-Linked Immunosorbent Assay (ELISA) Kits for PGE2, tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 (R&D Systems, Inc., Minneapolis, MN, USA); primary antibodies, including HO-1, cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS), and Nrf2, and responsible secondary antibodies for Western blot analysis (Santa Cruz Biotechnology, Santa Cruz, CA, USA); and tin protoporphyrin IX (SnPP), which is an inhibitor of HO activity (Porphyrin Products, Logan, UT, USA). All other chemicals were purchased from Sigma Chemical Co. (St. Louis, MO, USA), unless indicated otherwise.
Experimental animals
Male Kunming mice (8–9 weeks old) were purchased from Qingdao Institute for Food and Drug Control (approval number: SYXK (Lu) 2017 0002). All animals were maintained in a room at 22°C ± 2°C under a 12-h light/12-h dark cycle, with free access to food and water. All animal experiments were performed according to the rules and regulations of the Animal Ethics Committee at Qingdao University of Science and Technology (approval number: ACQUST-2017-023).
Sample preparation
E. brevicornu M. samples were obtained from the botanical garden of Wonkwang University, Iksan, Korea, in August 2012. Dried E. brevicornu M. leaves were finely powdered and used for extraction. In brief, 50 g of the material was extracted twice with ethanol solvent (70%) by refluxing for 2 h in each round. After the sample was cooled, the extract was filtered, dried by evaporation in vacuo, and stored in desiccators before use. The sample solution was prepared for cell experiments by dissolving in dimethyl sulfoxide (DMSO).
High-performance liquid chromatography
Chromatography was conducted through gradient elution on an Agilent 1220 Infinity LC System (Agilent Technologies, USA). Chromatographic separations were performed using a Hypersil BDS C18 column (4.6 × 250 mm, 5 μm; Thermo, USA) at 40°C with a flow rate of 1 mL/min. The mobile phase consisted of water (A) and acetonitrile (B). The gradient elution program was set as follows: 24% (B) for 0–16 min, 24–26% (B) for 16–26 min, 26–30% (B) for 26–35 min, and 30% (B) for 35–50 min. Chromatograms were acquired with a UV detector at 270 nm.
Cell culture
The RAW264.7 macrophage cell line was obtained from American Type Culture Collection (Rockville, MD, USA). The cells were cultured at 37°C in a humidified atmosphere containing 5% CO2 in DMEM supplemented with 10% heat-inactivated FBS, penicillin G (100 units/mL), and streptomycin (100 mg/mL).
Cell viability assay
The cytotoxicity of EBME was evaluated with a 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyl tetrazolium bromide (MTT) assay. RAW264.7 macrophages were seeded into 96-well plates (100 μL, 1 × 105 cells/mL in each well), treated with various EBME concentrations (12.5, 25, 50, 100, 200 μg/mL), and incubated for 24 h. Afterward, 2.5 mg/mL MTT was added to each well, and the cells were further incubated for 4 h. The supernatant was removed, and DMSO (150 μL) was added to dissolve formazan crystals formed in the viable cells. The absorbance of each well was determined at 490 nm by microplate reader.
Determination of NO and PGE2
RAW264.7 macrophages were cultured in 24-well plates (5 × 105 cells/mL), pretreated with EBME for 12 h in the presence or absence of SnPP (50 μM) and subsequently stimulated with LPS (1 μg/mL) for 24 h. The NO and PGE2 were measured according to previous protocol. 19 NO was determined by measuring nitrite, which is a stable end product of NO oxidation, 17 using Griess reaction. In brief, 100 μL of supernatant was mixed with the same volume of Griess reagent; absorbance of the mixture at 525 nm was determined with a microplate reader. In addition, 100 μL of supernatant was collected for the determination of PGE2 concentration using the ELISA Kit.
Preparation of cytosolic and nuclear fractions
RAW264.7 macrophages were homogenized in a PER-Mammalian Protein Extraction Buffer (Pierce Biotechnology, Rockford, IL, USA) containing freshly added protease inhibitor cocktail I (EMD Biosciences, San Diego, CA, USA) and 1 mM phenylmethylsulfonyl fluoride (PMSF). The cytosolic fraction of the cells was prepared through centrifugation. The cytoplasmic and nuclear extracts of RAW264.7 macrophages were isolated with NE-PER nuclear and cytoplasmic extraction reagents (Pierce Biotechnology) according to the manufacturer's instructions. Protein concentrations were determined using the Bicinchoninic Acid (BCA) Protein Assay Kit.
Western blot analysis
RAW264.7 macrophages were lysed in Tris-HCl buffer containing protease inhibitor mixture (0.1 mM PMSF, 5 mg/mL aprotinin, 5 mg/mL pepstatin A, and 1 mg/mL chymostatin). Protein extracts (30 μg) were separated through 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to a nitrocellulose membrane. The membrane was blocked with 5% dry nonfat milk in Tris-buffered saline Tween-20 (10 mM Tris-HCl, pH 7.6, 150 mM NaCl, and 0.05% Tween-20) for 1 h and subsequently incubated with anti-iNOS, anti-COX-2, anti-HO-1, anti-Nrf2 primary antibodies, and horseradish peroxidase-conjugated secondary antibody. The bands were observed using an enhanced chemiluminescence substrate (Amersham Bioscience, Buckinghamshire, UK) and quantified with an image analysis program (Image Gauge v3.12 software, Fujifilm, Tokyo, Japan). Protein concentration was determined using the BCA Protein Assay Kit.
LPS-induced peritonitis
Male Kunming mice were divided into six groups (n = 10). In the control and model groups, the mice received 0.9% saline solution through intragastric (i.g.) gavage for 7 days. The EBME groups included low-dose-EBME group (L-EBME), middle-dose-EBME group (M-EBME), high-dose-EBME group (H-EBME), the mice were administered EBME for 7 days at doses of 60, 120, and 240 mg/kg body weight (i.g.), respectively. In the positive control group, mice were intraperitoneally (i.p.) injected with dexamethasone (0.5 mg/kg) for 60 min before LPS (serotype 055:B5) was injected. After the treatment was administered, LPS (1 mg/kg) or sterile saline solution was given intraperitoneally. After 4 h, mice were sacrificed, and their peritoneal cavities were opened and washed with 1 mL of sterile phosphate-buffered saline containing heparin (25 UI/mL). Peritoneal lavage was collected using a syringe for TNF-α, IL-1β, and IL-6 assay by using the ELISA Kit.
Statistical analysis
Data are expressed as mean ± standard deviation (SD) for each experimental group and tested for normality by using SPSS version 17.0 for Windows. One-way ANOVA followed by Student's t-test and Tukey's range test was conducted to determine significant differences between groups. P < .05 was considered statistically significant.
Results
High-performance liquid chromatography analysis of EBME
The results of the high-performance liquid chromatography (HPLC) analysis of EBME are shown in the chromatograms in Figure 1. The peaks of epimedins A, B, and C and icariin, major bioactive constituents and marker compounds in Herba Epimedii, 20 in the extract (Fig. 1A), were identified according to the retention times of their reference samples (Fig. 1C–F) in HPLC.
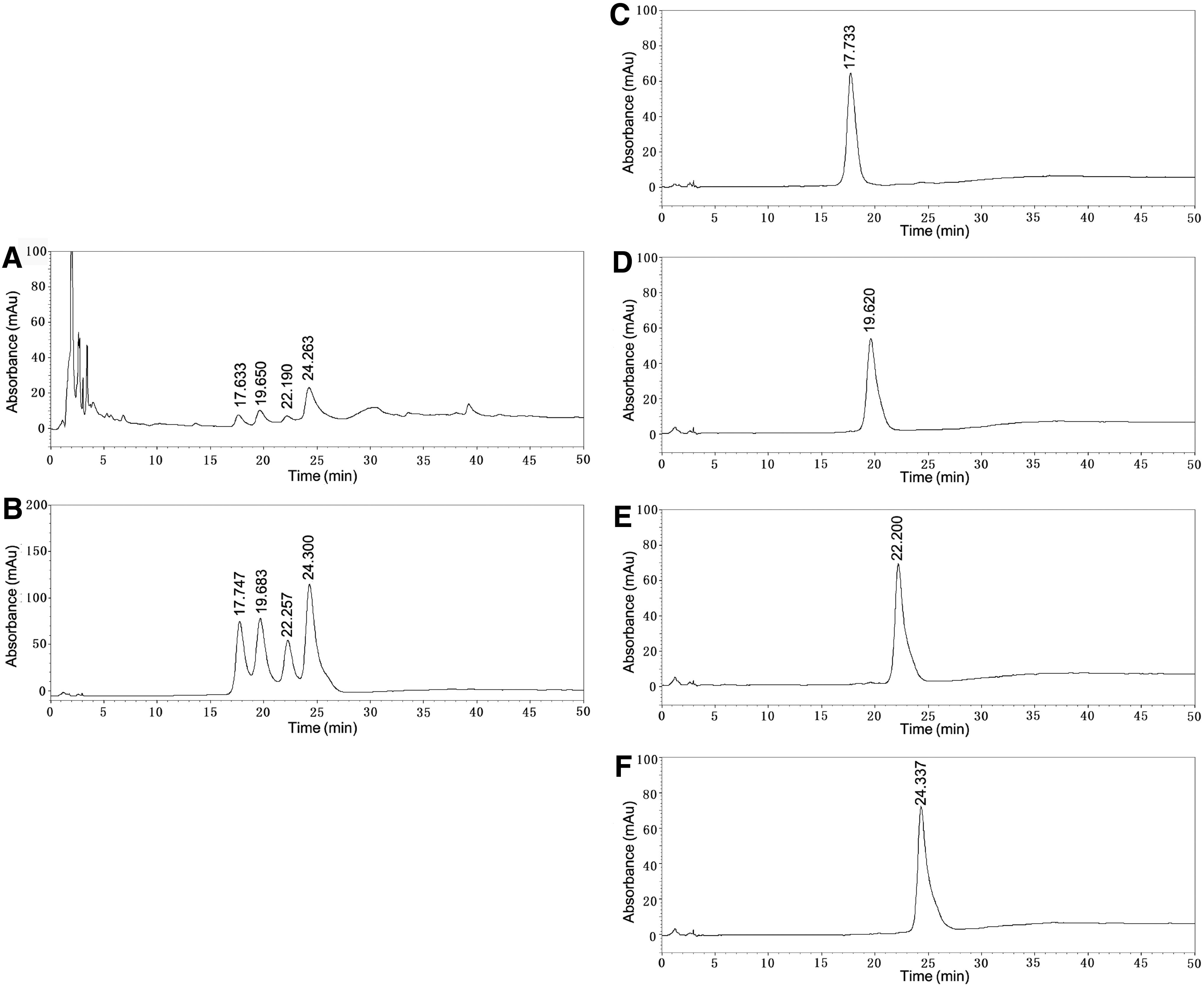
HPLC chromatogram of EBME. HPLC chromatogram of the EBME extract
Effects of EBME on the viability of RAW264.7 macrophages
The cytotoxicity of EBME on RAW264.7 macrophages was determined through the MTT assay. In Figure 2, EBME (up to 200 μg/mL) did not significantly change cell viability. Thus, the concentration range of EBME was maintained between 12.5 and 100 μg/mL for subsequent experiments.
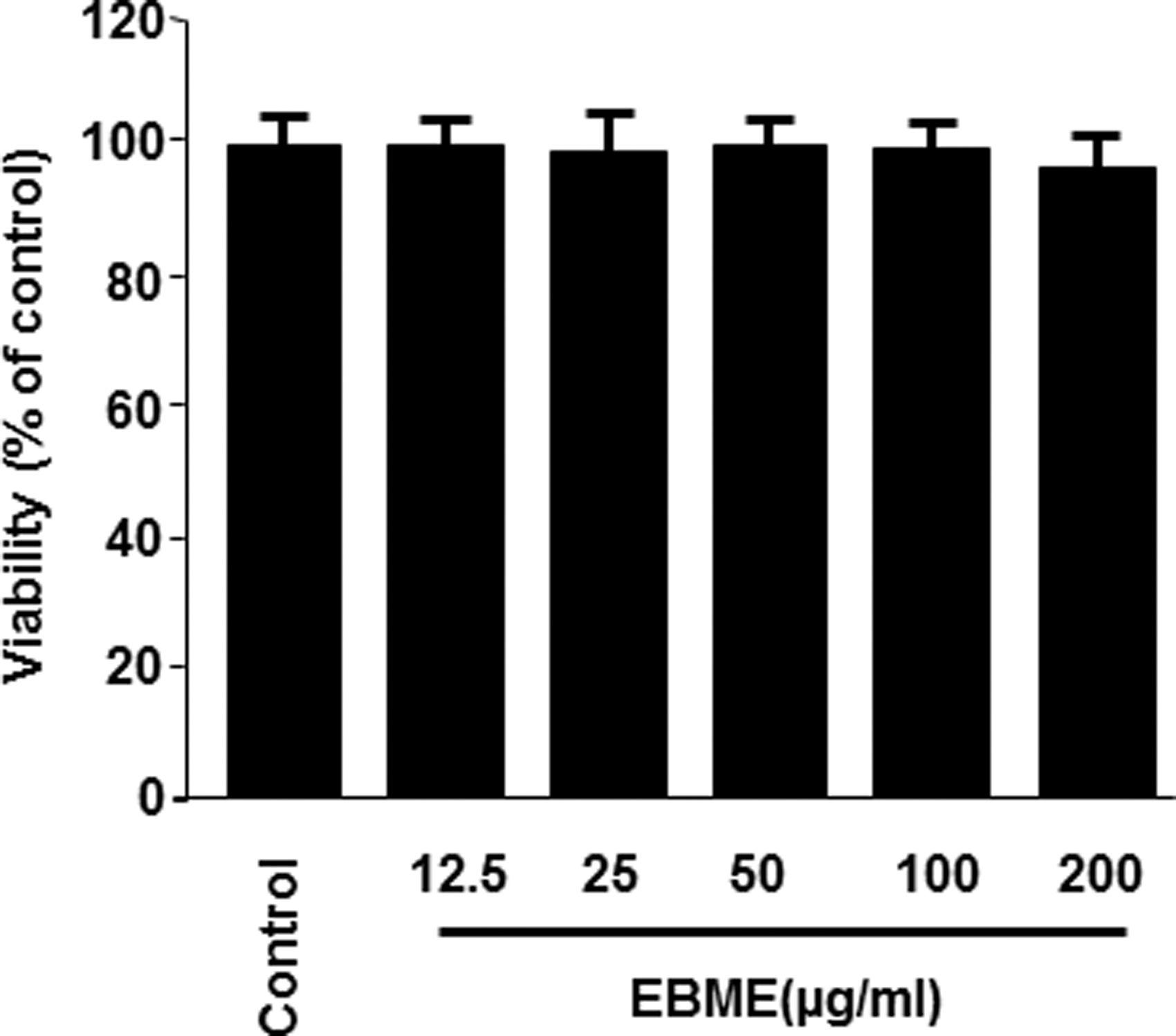
Effect of EBME on the viability of RAW264.7 macrophages. Cells were incubated for 24 h with the indicated concentrations of EBME, and the untreated cells were used as a control group. Cell viability was determined using MTT. The viability of control group was regarded as 100%. Data are represented as the mean values of three experiments ± SD. MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; SD, standard deviation.
Effects of EBME on the expression of proinflammatory mediators in LPS-stimulated RAW264.7 macrophages
To evaluate the anti-inflammatory effects of EBME in LPS-stimulated RAW264.7 macrophages, we investigated the inhibitory effects of EBME on proinflammatory mediators. In Figure 3A and B, EBME dose dependently suppressed LPS-stimulated iNOS and COX-2 protein expression. In Figure 3C and D, EBME dose dependently decreased iNOS-derived NO and COX-2-derived PGE2 production. In particular, EBME at concentrations of 50 and 100 μg/mL, significantly decreased the expression of iNOS, COX-2, NO, and PGE2 compared with LPS treatment only (P < .05). These results suggested that EBME suppressed LPS-induced proinflammatory mediators, including iNOS, COX-2, NO, and PGE2.

Effects of EBME on the expression of proinflammatory mediators in LPS-stimulated RAW264.7 macrophages. RAW264.7 macrophages were pretreated for 12 h with the indicated concentrations of EBME and were subsequently stimulated for 24 h with LPS (1 μg/mL). The expressions for iNOS
Effects of EBME on HO-1 protein expression and Nrf2 nuclear translocation in RAW264.7 macrophages
To clarify the mechanisms underlying the inhibition of proinflammatory mediators by EBME, we investigated the effects of EBME on HO-1 protein expression and Nrf2 nuclear translocation. In Figure 4A, the effects of EBME on HO-1 protein expression were similar to those of cobalt protoporphyrin, a HO-1 inducer, and EBME dose dependently induced the expression of HO-1. In Figure 4B, the HO-1 induction of EBME became evident after 3 h and reached the maximum after 18 h (P < .05). Considering the important role of Nrf2 in HO-1 expression, we further determined whether EBME treatment induces the nuclear translocation of Nrf2. Figure 4C and D shows that EBME dose dependently reduced the cytoplasmic Nrf2 levels and increased the nuclear Nrf2 levels in RAW264.7 macrophages. These results indicate that EBME could induce HO-1 protein expression and Nrf2 nuclear translocation.
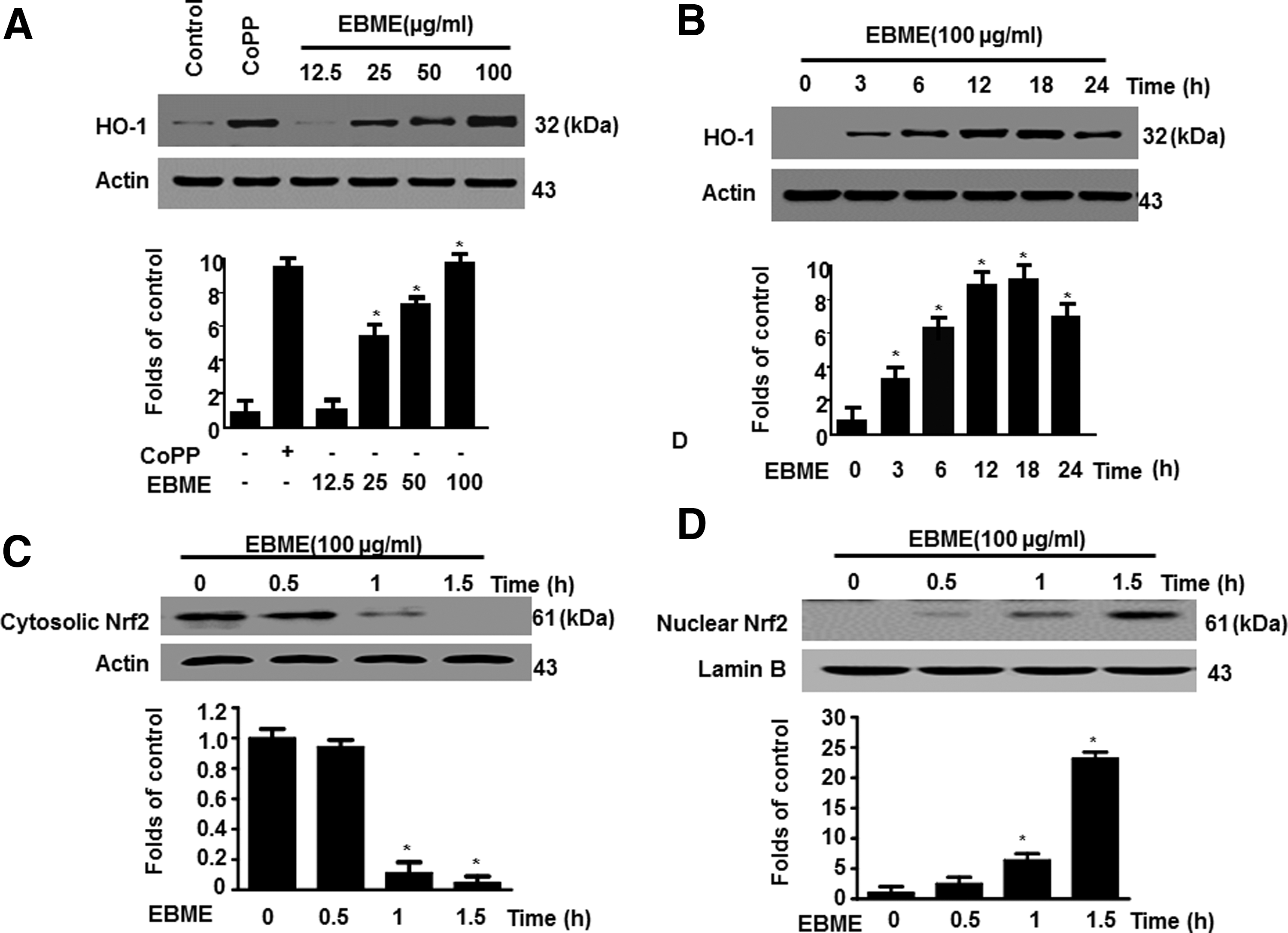
Effects of EBME on the HO-1 protein expression and Nrf2 nuclear translocation in RAW264.7 macrophages. RAW264.7 macrophages were incubated with indicated concentrations of EBME for 12 h
Effects of HO-1 expression on the inhibition of nitrite and PGE2 by EBME in LPS-stimulated RAW264.7 macrophages
To determine whether the anti-inflammatory activities of EBME are associated with the increased HO-1 expression, we investigated the inhibitory effects of EBME on NO and PGE2 expression in the presence or absence of SnPP. In Figure 5, the inhibitory effects of EBME on LPS-induced NO (Fig. 5A) and PGE2 (Fig. 5B) production were partially reversed by SnPP. These results suggested that EBME inhibited the expression of proinflammatory mediators by inducing HO-1 expression.
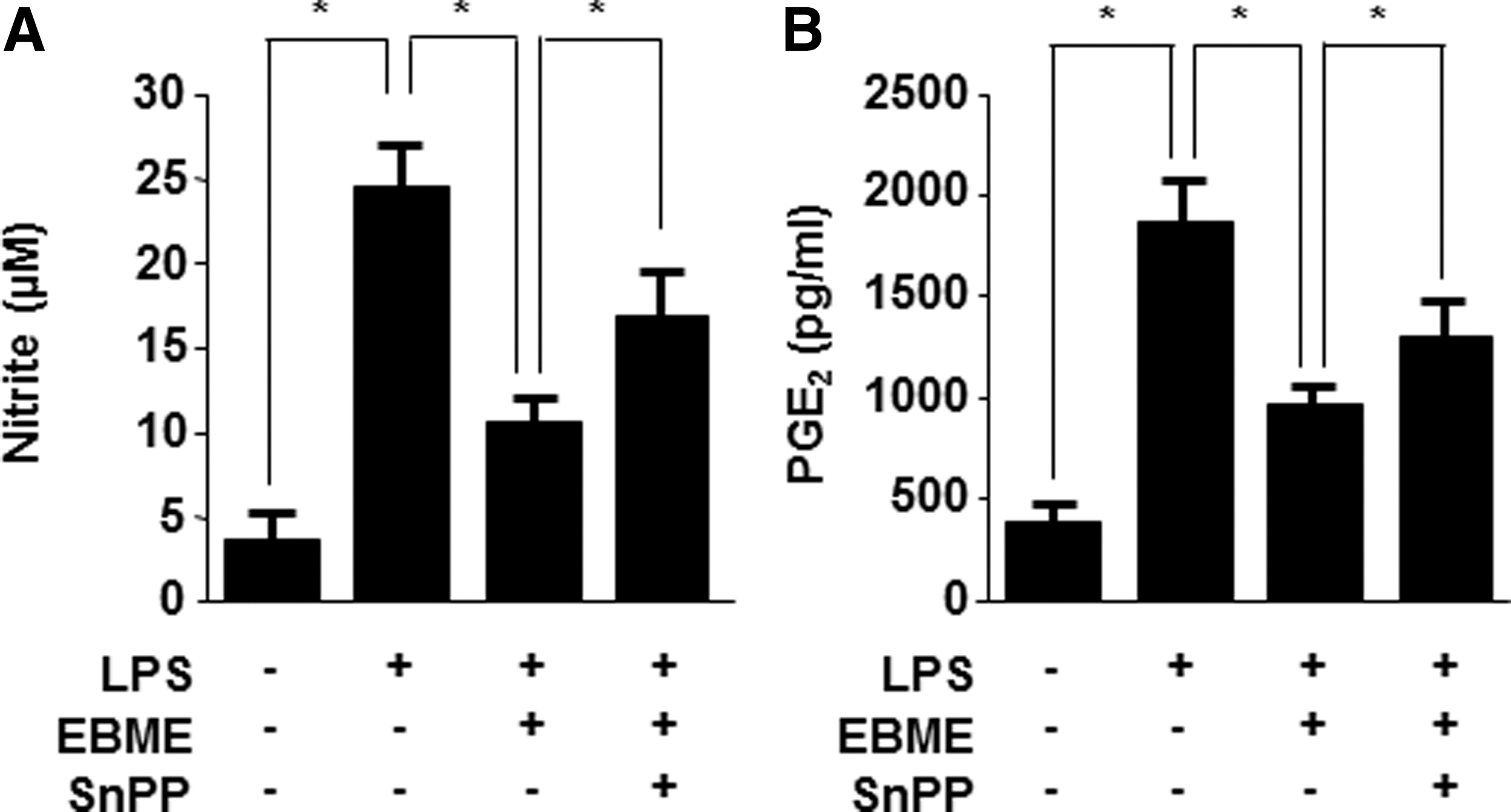
Effects of HO-1 expression on the inhibition of nitrite and PGE2 by EBME in LPS-stimulated RAW264.7 macrophages. RAW264.7 macrophages were pretreated with EBME for 12 h in the presence or absence of SnPP (50 μM) and were subsequently stimulated with LPS (1 μg/mL) for 24 h. The concentrations of nitrite
Effects of EBME on the levels of inflammatory cytokine in LPS-induced peritonitis
To verify the anti-inflammatory effect of EBME in vivo, we investigated the inhibitory effects of EBME on the levels of TNF-α, IL-1β, and IL-6 in LPS-induced peritonitis. In Table 1, the levels of TNF-α, IL-1β, and IL-6 in the peritoneal lavage were significantly increased in the LPS-injected mice compared with that of the control group (P < .05). However, EBME treatment dose dependently reduced the levels of TNF-α, IL-1β, and IL-6. All of the EBME doses could significantly inhibit TNF-α (P < .05), and M-EBME and H-EBME evidently decreased IL-1β and IL-6 (P < .05). In addition, the effects of decreased TNF-α, IL-1β, and IL-6 with H-EBME were similar to those observed in the positive group (dexamethasone) and did not show any significant differences. These results suggested that EBME could be used effectively to treat LPS-induced peritonitis.
Values in the same column with different superscript letters indicate significant difference between groups (P < .05) by the Tukey's test. Control, control group; Dex, dexamethasone-treatment group; EBME, E. brevicornu Maxim ethanol extract; H-EBME, high-dose-EBME group; IL, interleukin; L-EBME, low-dose-EBME group; LPS, lipopolysaccharide; LPS, LPS-induced model group; M-EBME, middle-dose-EBME group; TNF-α, tumor necrosis factor-α.
Discussion
Many natural herbal medicines have been demonstrated to be safe and effective, some of their extracts show potential for anti-inflammatory applications. 21 –24 As a traditional herbal drug, E. brevicornu M. has several biological activities. In this study, the anti-inflammatory effects and mechanisms of EBME were evaluated using in vivo and in vitro inflammation models.
LPS-induced macrophages produce various inflammatory mediators, 17 such as NO and PGE2. 25,26 NO, which plays an important role in nonspecific immune responses to infection, host defense, and cytotoxicity, 27 is also implicated as a proinflammatory agent because of its high expression levels during human inflammatory reactions. 28 PGE2, a principal mediator of inflammation, 29 can trigger inflammatory responses, such as increasing pain responses and promoting pyrogenic effects. 30 iNOS and COX-2, activated by LPS and inflammatory cytokines, are key enzymes that regulate NO and PGE2 production, respectively. 28,31 Inhibiting the production of proinflammatory mediators and suppressing the mechanisms responsible for the activation of inflammatory responses are regarded as new strategies to control inflammation. 32 –34 Thus, compounds that reduce NO, PGE2, iNOS, and COX-2 production are desirable as anti-inflammatory agents. In this study, EBME suppressed the expression of NO, PGE2, iNOS, and COX-2 in LPS-stimulated RAW264.7 macrophages (Fig. 3), suggesting that EBME could act as an anti-inflammatory agent. In addition, the inhibitory effects of EBME against inflammatory mediators, including NO and PGE2, might be attributed to the inhibited expression of iNOS and COX-2.
HO-1 is an important cytoprotective factor that counteracts the detrimental effects of inflammation. 12,35 –37 Stress-dependent HO-1 induction is primarily regulated by Nrf2. 15 Nrf2-mediated HO-1 expression is a possible therapeutic target for the treatment of inflammatory disorders. In this study, EBME induced HO-1 protein expression (Fig. 4A, B) and Nrf2 nuclear translocation (Fig. 4C, D). These results indicated that EBME might exert anti-inflammatory effects by inducing HO-1 expression through Nrf2 nuclear translocation.
The inhibitory effects of EBME on LPS-stimulated NO and PGE2 production were partially reversed by SnPP (Fig. 5). This result further confirmed that HO-1 expression was involved in the anti-inflammatory activities of EBME. Our results provided evidence that EBME exerted anti-inflammatory activity through the Nrf2/HO-1 pathway. Conversely, SnPP did not completely reduce the LPS-induced expression of NO and PGE2 (Fig. 5), indicating that other signaling pathways were involved in the anti-inflammatory activities of EBME.
In a previous study, it was generally demonstrated that the aqueous extract of Epimedium Herb exerted anti-inflammatory effects in xylene-induced mouse ear edema. 4 In this study, we further investigated the anti-inflammatory effects of Epimedium in LPS-induced peritonitis. Peritonitis, characterized by an increase in the concentrations of inflammatory cytokines, including TNF-α, IL-1β, and IL-6, recruited to the peritoneal cavity, is widely used to evaluate the anti-inflammatory activity of drug candidates. 38 TNF-α, a prototype of proinflammatory cytokines, not only induces its own production, but also stimulates the secretion of other inflammatory cytokines. 39 It is a central regulator of inflammation. 40 IL-1β also exerts a strong proinflammatory activity and promotes the recruitment of inflammatory cells to the site of inflammation by inducing the expression of adhesion molecules and the release of chemokines. 39 IL-6 participates in the development of inflammation to cancer and acts as an endogenous mediator in LPS-induced peritonitis. 38 In the present study, pretreatment with EBME robustly attenuated the production of TNF-α and reduced the production of IL-1β and IL-6 in LPS-induced peritonitis. This result indicated that the oral administration of EBME could prevent acute inflammation by downregulating the production of inflammatory mediators.
In summary, EBME inhibits the LPS-stimulated release of inflammatory mediators, including NO and PGE2, in RAW264.7 macrophages by suppressing iNOS and COX-2 expression. The anti-inflammatory effects of EBME are partly mediated by the induced HO-1 expression through Nrf2 signaling. The anti-inflammatory effects of EBME in vivo were confirmed in LPS-induced peritonitis. Thus, EBME may be beneficial for treating inflammation.
Our study is limited by the exclusion of compounds responsible for the anti-inflammatory activities of EBME. The major compounds that induce this effect should be elucidated in further research.
Footnotes
Acknowledgments
This study was supported by the Outstanding Young Scientists Foundation Grant of Shandong Province (No. BS2014YY010), Key Laboratory of Natural Resources of Changbai Mountain & Functional Molecules (Yanbian University), Ministry of Education, China (No. CSSHZ-2014-03), and National Natural Science Foundation of China (No. 81360686 and 31500288).
Author Disclosure Statement
No competing financial interests exist.