
Abstract
Anthriscus sylvestris (L.) Hoffm. is a common perennial herb that is widely distributed in Europe, Korea, and New Zealand. The root of A. sylvestris has been used in Korean traditional medicine as an antitussive and cough remedy. However, the physiologically active function of A. sylvestris leaves is not yet known. In this study, we evaluated the anti-inflammatory effects, as well as the underlying molecular mechanisms of an aqueous extract of A. sylvestris leaves (AE-ASL) in vitro and in vivo. Our results indicated that pretreatment with AE-ASL significantly inhibited the lipopolysaccharide (LPS)-induced secretion of nitric oxide (NO) and prostaglandin E2 in RAW264.7 cells, without showing cytotoxicity. In addition, the LPS-induced mRNA and protein expression of inducible NO synthase, cyclooxygenase-2, and inflammatory mediators such as tumor necrosis factor alpha interleukin (IL)-1β, and IL-6 was attenuated by pretreatment with AE-ASL in a dose-dependent manner. Therefore, we investigated the activation of nuclear factor (NF)-κB, a transcription factor regulating the expression of inflammation-related genes. AE-ASL inhibited the nuclear translocation of the NF-κB p65 subunit by suppressing the phosphorylation and degradation of the inhibitor of NF-κB (IκBα). Further, AE-ASL inhibited the LPS-induced phosphorylation of mitogen-activated protein kinases (MAPKs) in RAW264.7 cells. Orally administered AE-ASL (50, 100, and 200 mg/kg of body weight [BW]) suppressed the development of carrageenan-induced rat paw edema by 15%, 31%, and 40%, respectively, after 4 h. Altogether, our results suggest that AE-ASL possesses anti-inflammatory activity, based on the suppression of NF-κB and MAPK pathways in vitro and inhibition of the carrageenan-induced paw edema in vivo.
Introduction
I
Medicinal plants produce bioactive components, including various acidic and phenolic compounds, with more than 25% of medicines approved in the past 30 years having been plant derived. 12 Anthriscus sylvestris (L.) Hoffm. belongs to the Apiaceae (syn. Umbelliferae) family and grows in hedges, road verges, and neglected pastures in Europe, Asia, and New Zealand. 12,13 In Asia, including China, dried roots of this plant have traditionally been used as an antipyretic, cough, and diuretic remedy. 14,15 Aerial parts have been used to treat headaches in Ireland and Tunisia and rheumatism and other inflammatory ailments in India. 16 –18 Some studies have reported that ethanol and methanol extracts of dried aerial parts of A. sylvestris showed antioxidant activity, and the main active constituents were quercetin, apigenin, and chlorogenic acid. 19,20 However, the physiologically active fraction of aqueous extract of A. sylvestris leaves (AE-ASL) is as yet unknown.
Therefore, the purpose of this study was to evaluate the anti-inflammatory effects and the underlying mechanisms of AE-ASL in vitro and in vivo.
Materials and Methods
Preparation of AE-ASL
Dried A. sylvestris leaves (1.44 kg), purchased form Heuksando, Korea, were ground and extracted with 27 volumes of water (v/w) at 90°C for 3 h, followed by filtration by utilizing a 55-μm bag filter. The extract was concentrated in a rotary vacuum evaporator (Eyela, Tokyo, Japan) and lyophilized. The dried extract (440 g) was dissolved in distilled water (100 mg/mL), and the solution was filtered through a 0.45-μm filter. The extract was stored at −20°C until use. The yield of the aqueous extract was about 30.6% (w/w).
Reagents
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), lipopolysaccharide (LPS), sulfanilamide, N-(1-naphthyl)ethylenediamine dihydrochloride, phosphoric acid, and carrageenan were purchased from Sigma-Aldrich (St. Louis, MO, USA). A prostaglandin E2 (PGE2) ELISA kit was purchased from R&D Systems (Minneapolis, MN, USA). Primary and secondary antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA), except anti-inducible NO synthase (iNOS) (Abcam, Cambridge, MA, USA) and anti-β-actin (AB Frontier, Seoul, Korea). Dulbecco's modified Eagle's medium (DMEM) and a penicillin/streptomycin solution were purchased from WelGene (Daegu, Korea). Fetal bovine serum (FBS) was purchased from Corning (Corning, NY, USA). Methanol and acetonitrile and water of HPLC grade were obtained from Fisher Scientific (Fair Lawn, NJ, USA). All other common chemicals and solvents were of analytical reagent grade or HPLC grade.
HPLC analysis
The sample was analyzed by using a Shimadzu HPLC system equipped with a photo diode array (Shimadzu, Kyoto, Japan). The sample solutions (1 mg/mL) were separated by using a 5C18 MS ll column (4.6 mm i.d. × 250 mm; Cosmosil, Japan) at 40°C. The mobile phase was a mixture of two liquids distributed by the 0.1% trifluoroacetic acid/DW (TFA/DW) (A) and 0.1% trifluoroacetic acid/acetonitrile (TFA/MeCN) (B) at a flow rate of 1.0 mL/min. The gradient elution program was used as follows: 90% A (0.5 min), 15% A (30 min), 5% A (40 min), and 90% A (40.1 min). The injection volume was 10 μL, and detection was at a wavelength of 320 nm. The mass measurement of compounds (1, 2, and 3) was analyzed by using a Nexera X2 HPLC system coupled to LCMS-IT-TOF (Shimadzu) and included a CBM-20A system controller, an LC-30AD pump, an SIL-20AC autosampler, a CTO-30A column oven, and an SPD-M30A diode array detector with a Capcell Core C18 Column (Shiseido, Kyoto, Japan). The gradient program was used with the mobile phase, combining 0.1% TFA/DW (A) and 0.1% TFA/MeCN (B) as follows: 90% A (0.5 min), 15% A (20 min), 5% A (25 min), and 90% A (25.1 min). The parameters were as follows: flow rate, 0.2 mL/min; injection volume, 5 μL; wavelength, at 320 nm; voltage, 4.50 kV as ESI positive mode; curved desolvation line temperature, 200°C; block heater temperature, 200°C; nebulizer gas flow, 1.5 L/min; and ion accumulation time, 10 msec. All data were recorded and analyzed by Shimadzu software; LCMS solution ver. 3.60, Formula Predictor Ver. 1.2, and Accurate Mass Calculator (Shimadzu).
Cell culture and viability assay
Murine macrophage RAW264.7 cells were obtained from the Korea Research Institute of Bioscience and Biotechnology (Daejeon, Korea). Cells were cultured in DMEM containing 10% FBS and 1% penicillin/streptomycin in a humidified incubator with 5% CO2 at 37°C. For the cell viability assay, 1 × 106 cells/mL were incubated for 12 h and then treated with AE-ASL (0, 10, 25, 50, 100, and 200 μg/mL) for 24 h. Cell viability was determined by the MTT assay.
Measurement of NO and PGE2
RAW264.7 cells (1 × 106 cells/mL) were pretreated with AE-ASL (10, 25, 50, and 100 μg/mL) for 1 h and then cultured with LPS (0.2 μg/mL) for 24 h. Nitrite accumulation in the culture medium was measured by using the Griess reagent. Briefly, 100 μL of each supernatant, mixed with an equal volume of the Griess reagent [1% (w/v) sulfanilamide and 0.1% (w/v) naphthylethylenediamine in 5% (v/v) phosphoric acid], was incubated in a dark room for 5 min, and absorbance was measured at 540 nm in a microplate reader (Epoch; BioTek Instruments, Winooski, VT, USA). Nitrite was quantified based on a sodium nitrite standard curve. PGE2 concentrations in the culture medium were quantified by using a Parameter™ PGE2 assay kit (R&D Systems) according to the manufacturer's instructions.
Isolation of total RNA and reverse transcription–polymerase chain reaction
Total RNA was isolated from RAW264.7 cells by using Trizol reagent (Invitrogen, Carlsbad, CA, USA). RNA (1 μg) was reverse transcribed by using MMLV reverse transcriptase (Enzynomics, Daejeon, Korea) and then amplified by polymerase chain reaction (PCR) by using specific primers shown in Table 1. The glyceraldehyde 3-phosphate dehydrogenase (Gapdh) gene was used as an internal control for normalization.
COX, cyclooxygenase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IL, interleukin; iNOS, inducible nitric oxide synthase; TNFα, tumor necrosis factor alpha.
Preparation of whole-cell, nuclear, and cytosolic extracts
RAW264.7 cells were pretreated with AE-ASL for 1 h and stimulated with LPS (0.2 μg/mL) for 24 h. To obtain whole-cell lysates, cell pellets were lysed with a PRO-PREP protein extraction solution (iNtRON Biotechnology, Korea) for 30 min on ice. After centrifugation at 15,000 g for 15 min at 4°C, the whole-cell lysates were transferred to new tubes. Cytoplasmic and nuclear extracts were prepared by using NE-PER™ nuclear and cytoplasmic extraction reagents (Thermo Scientific, IL, USA) according to the manufacturer's instructions.
Western blot analysis
Proteins (20 μg) were separated by 10% or 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred onto a polyvinylidene difluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA). The membranes were blocked with 5% bovine serum albumin in Tris-buffered saline containing 0.1% Tween 20 (TBST) for 1 h and then incubated with specific primary antibodies at 4°C overnight. After washing three times with TBST, membranes were incubated for 1 h with a horseradish peroxidase-conjugated secondary antibody at room temperature. Proteins were detected by using an enhanced chemiluminescence kit (Millipore, Bedford, MA, USA) and visualized by using a MicroChemi 4.2 imager (DNR Bioimaging Systems, Jerusalem, Israel).
Animals
Male Sprague–Dawley rats, weighing 300 g, were purchased from Koatech (Gyeonggi, Korea). The animals were housed in a temperature (21°C ± 1°C)- and humidity (55% ± 5%)-controlled room with a 12-h light/dark cycle and were allowed free access to commercial pellets and water. All animal handling procedures were in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. 21 The experimental protocol was approved by the Chosun University Institutional Animal Care and Use Committee (CIACUC2017-A0011).
Carrageenan-induced paw edema in rats
The rats were randomly divided into six groups (n = 4 each): group 1 (normal, 0.9% normal saline), group 2 (paw edema, 0.9% normal saline), groups 3–5 (paw edema, 50, 100, and 200 mg/kg AE-ASL, respectively), and group 6 (paw edema, 10 mg/kg diclofenac sodium). The edema was induced by a subcutaneous injection of 0.1 mL of a 1% (w/v) carrageenan type IV suspension in 0.9% normal saline into the left hind paw 1 h after oral administration of AE-ASL. The paw thickness was measured in the middle of the paw by using a digital caliper before (time 0) and at different time points (up to 24 h) after carrageenan injection, as previously described. 22,23 The anti-inflammatory activity was calculated as previously described. 22
Statistical analysis
All data were derived from at least three independent experiments (except the in vivo study). The results are expressed as the means ± standard deviation (SD). A one-way analysis of variance, followed by Dunnett's test, was used for multiple comparisons by using GraphPad Prism (GraphPad Software, CA, USA). Statistical significance was set at P < .05.
Results
Effects of AE-ASL on viability of RAW264.7 cells
The MTT assay was used to evaluate the cytotoxicity of AE-ASL in RAW264.7 cells. As shown in Figure 1, AE-ASL did not exhibit cytotoxic effects up to 100 μg/mL. However, at a higher concentration (200 μg/mL), AE-ASL decreased the cell viability by ∼30%. Therefore, all further experiments were performed at concentrations of up to 100 μg/mL to avoid cytotoxicity.
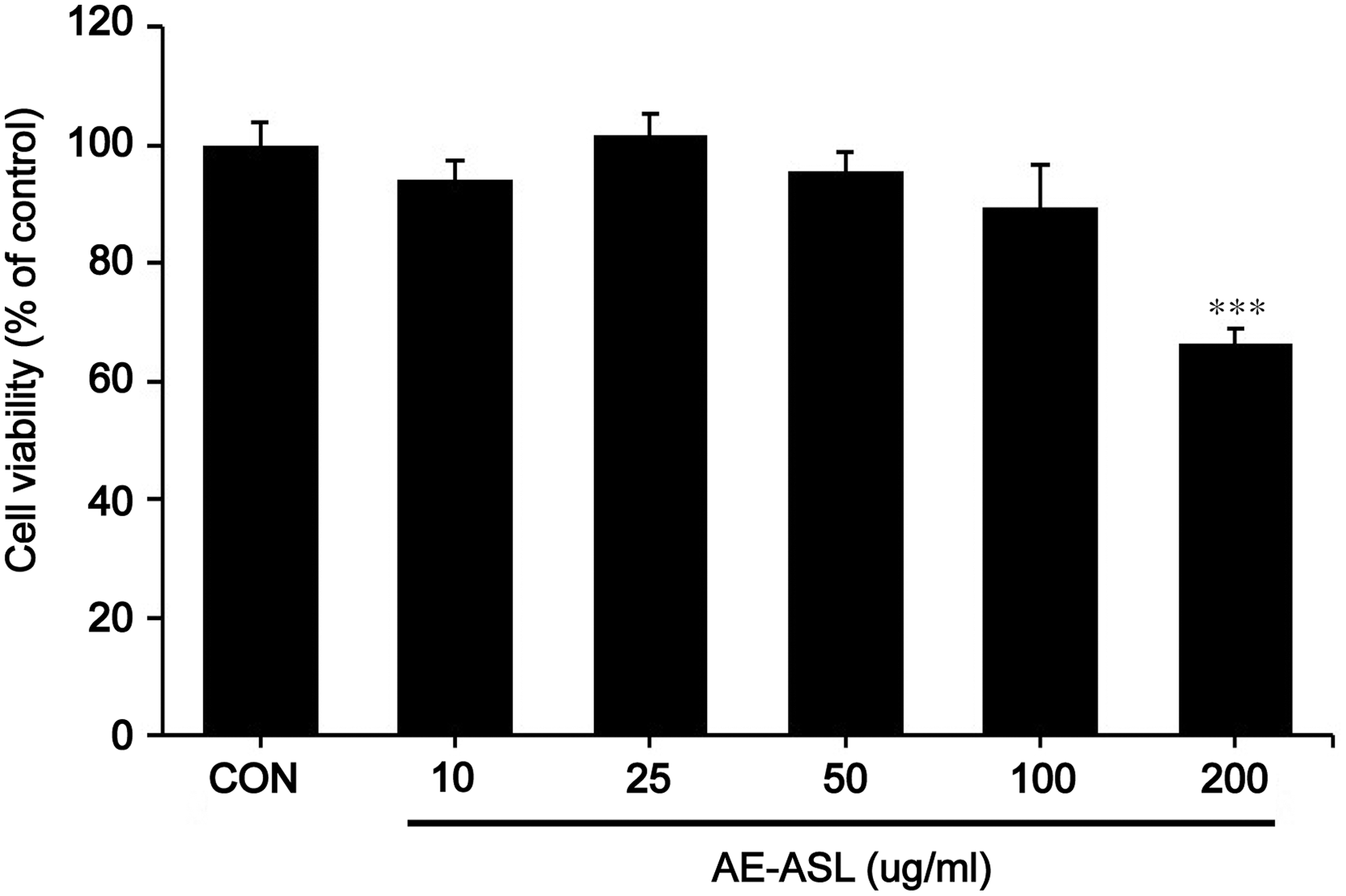
Effects of AE-ASL on cell viability of RAW264.7 cells. Cells were treated with various concentrations of AE-ASL (10, 25, 50, 100, and 200 μg/mL) for 24 h. Cell viability was determined by using an MTT assay, and the results were expressed as the percentage of control. Data are the means ± SD of three independent experiments. ***P < .001 compared with the control group. AE-ASL, aqueous extract of Anthriscus sylvestris leaves; MTT, 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; SD, standard deviation.
Inhibitory effects of AE-ASL on LPS-induced NO and PGE2 production and expression of iNOS and COX-2 in RAW264.7 cells
LPS alone significantly increased the NO and PGE2 production, 18-fold (18.14 ± 0.52 μM) and 2-fold (1700.42 ± 9.39 pg/mL), compared with that in the control (0.599 ± 1.01 μM and 808.48 ± 90.72 pg/mL, respectively). However, AE-ASL inhibited the LPS-induced NO production in a dose-dependent manner, by 30% (14.48 ± 0.52 μM), 60% (6.15 ± 1.0 μM), and 80% (3.24 ± 0.52 μM) at 25, 50, and 100 μg/mL, respectively (Fig. 2A). Moreover, LPS-induced PGE2 production was reduced after pretreatment with 25, 50, and 100 μg/mL AE-ASL by ∼10%, 15%, and 35%, respectively, compared with that observed with LPS alone (Fig. 2E). To elucidate the inhibitory effects of AE-ASL on the LPS-induced NO and PGE2 production, we evaluated the mRNA and protein expression levels of iNOS and COX-2 by using reverse transcription–PCR (RT-PCR) and western blotting. Pretreatment with AE-ASL for 1 h inhibited the mRNA and protein expression of LPS-induced iNOS and COX-2 in a dose-dependent manner (Fig. 2B–D, F–H). Taken together, these data suggest that the AE-ASL-mediated inhibition of NO and PGE2 production was associated with the regulation of iNOS and COX-2 expression in the LPS-stimulated RAW264.7 cells.
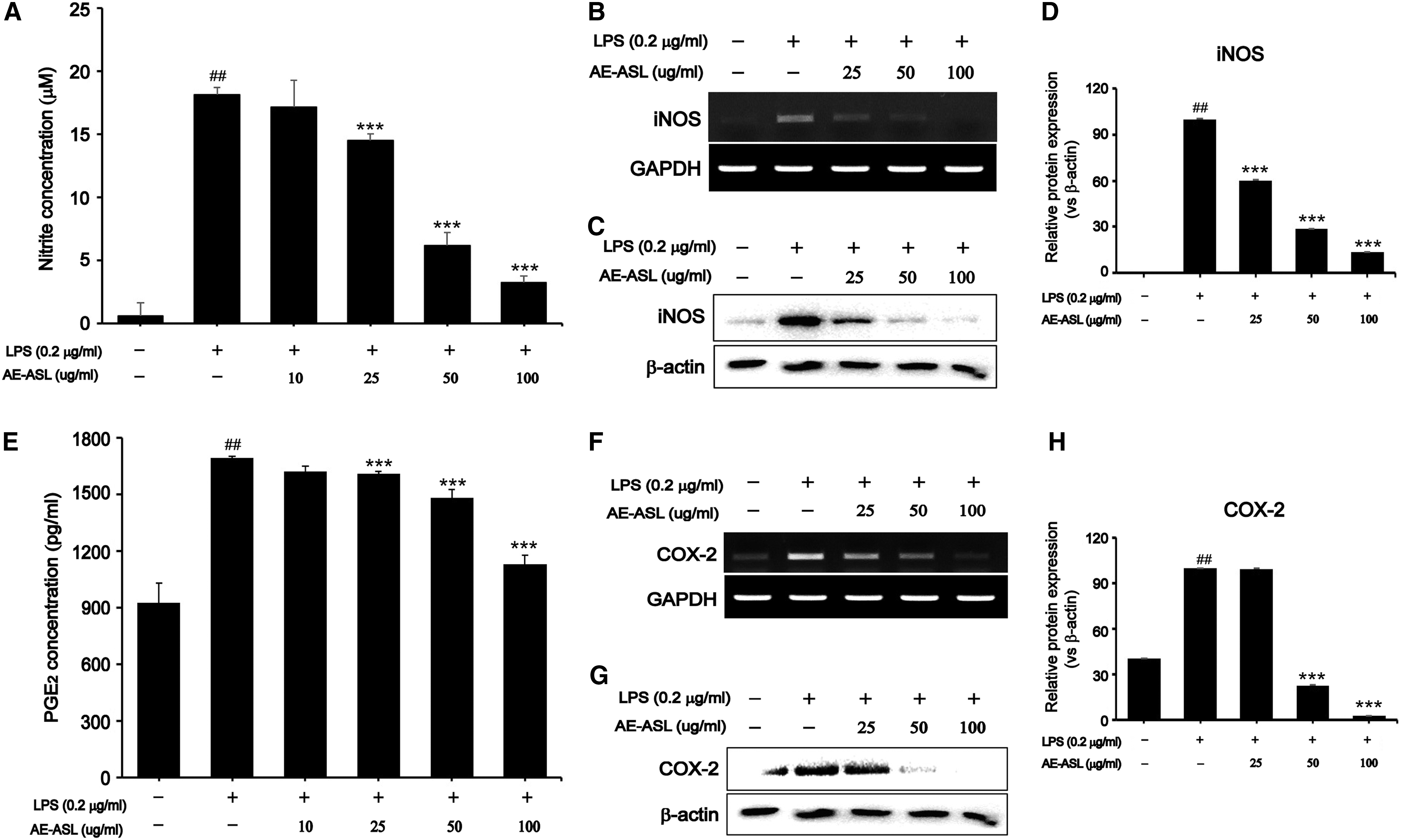
Inhibitory effects of AE-ASL on LPS-induced proinflammatory mediators in RAW264.7 cells. Cells were pretreated with AE-ASL (10, 25, 50, and 100 μg/mL) for 1 h, followed by LPS (0.2 μg/mL) stimulation for 24 h.
Inhibitory effects of AE-ASL on LPS-induced proinflammatory cytokines in RAW264.7 cells
Since AE-ASL inhibited NO and PGE2 production, we examined its effects on the expression of proinflammatory cytokines such as tumor necrosis factor alpha (TNFα), interleukin (IL)-1β, and IL-6, which play important roles in immune responses. As shown in Figure 3, LPS stimulation led to significant increases in the mRNA and protein levels of TNFα, IL-1β, and IL-6 compared with those in the control. However, pretreatment with AE-ASL at 100 μg/mL inhibited the LPS-induced mRNA expression of TNFα, IL-1β, and IL-6 by 76%, 73%, and 59%, respectively (Fig. 3A–D). The protein expression levels were also inhibited on exposure to 25–100 μg/mL AE-ASL (Fig. 3E–H). These results suggest that AE-ASL significantly suppressed the LPS-induced TNFα, IL-1β, and IL-6 expression, which supports the hypothesis that AE-ASL has anti-inflammatory activity.
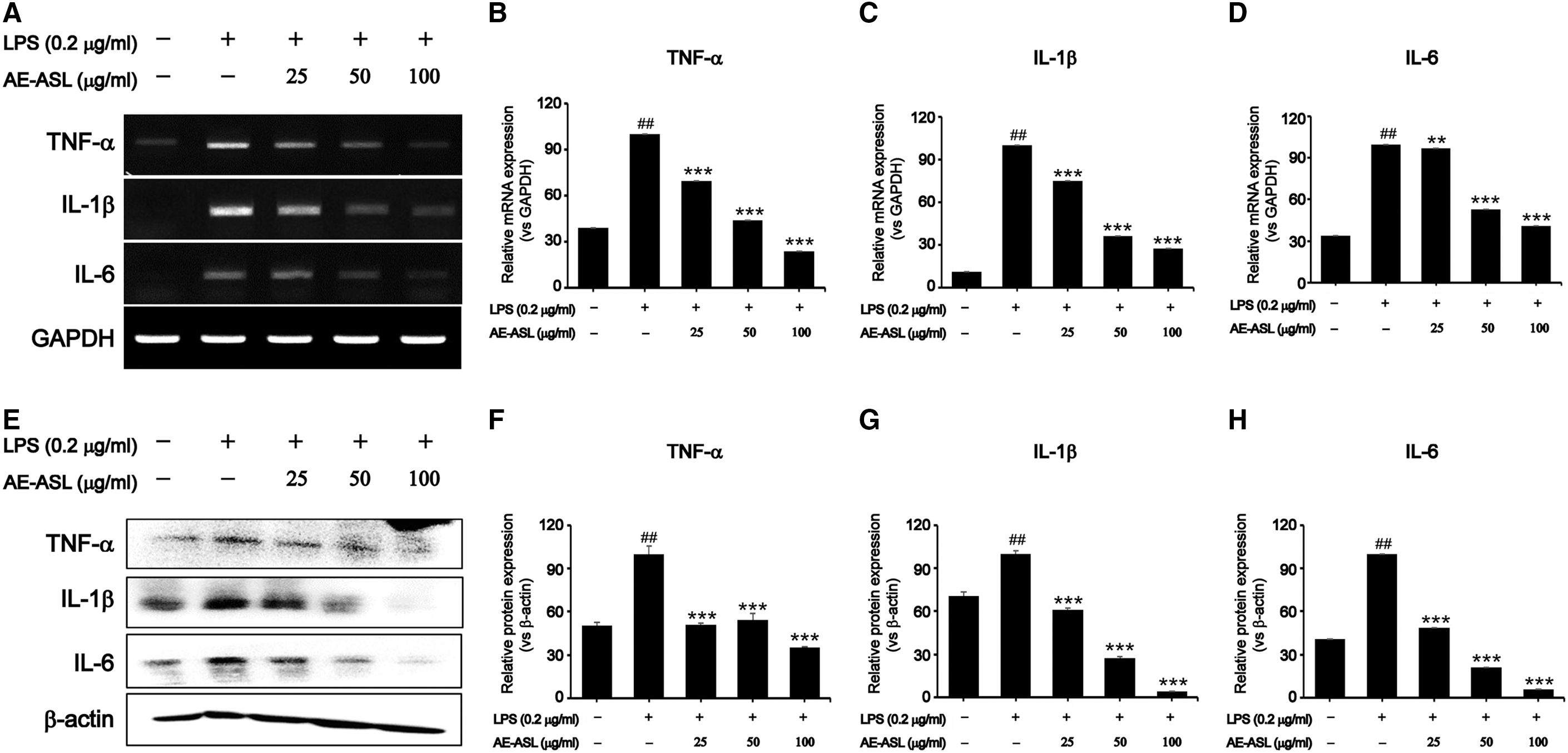
Inhibitory effects of AE-ASL on LPS-induced proinflammatory cytokines in RAW264.7 cells. Cells were pretreated with AE-ASL (25, 50, and 100 μg/mL) for 1 h, followed by LPS (0.2 μg/mL) stimulation for 24 h.
Suppression of NF kappa-B signaling pathway by AE-ASL in LPS-stimulated RAW264.7 cells
LPS stimulation significantly increased the phosphorylation and degradation of IκBα, the inhibitor of NF-κB. However, AE-ASL pretreatment markedly reversed the phosphorylation and degradation of IκBα in a dose-dependent manner (Fig. 4A, B). Consistent with IκBα degradation, the translocation of NF-κB p65 from the cytosol to the nucleus markedly increased on LPS stimulation (Fig. 4C–E). However, the LPS-induced nuclear translocation of NF-κB p65 was significantly inhibited by AE-ASL pretreatment (Fig. 4C–E). These results suggest that AE-ASL effectively suppresses the NF-κB signaling pathway via inhibition of IκBα degradation, preventing the nuclear translocation of the p65 NF-κB subunit in LPS-stimulated macrophages (Fig. 4).
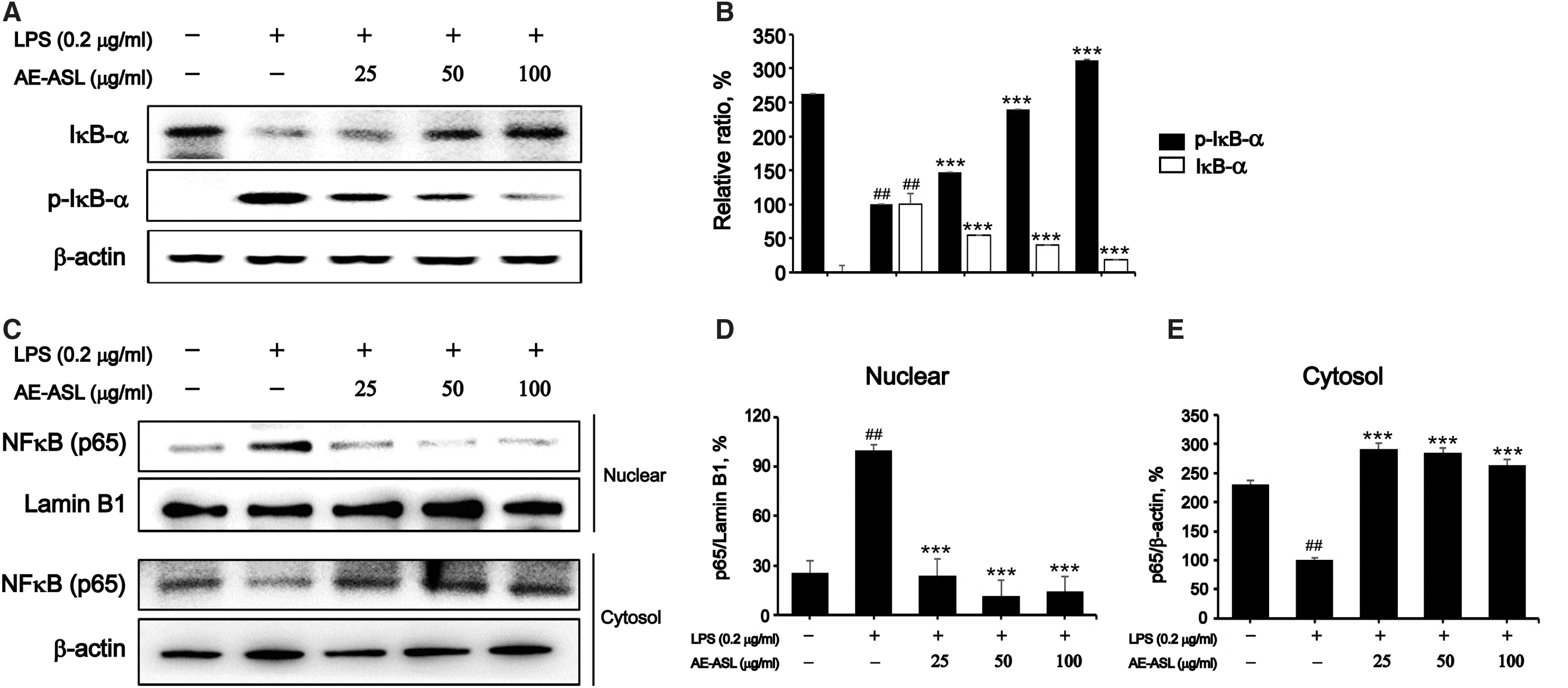
Effects of AE-ASL on LPS-induced phosphorylation of IκBα and NF-κB p65 nuclear translocation in RAW264.7 cells. Cells were pretreated with AE-ASL (25, 50, and 100 μg/mL) for 1 h, followed by LPS (0.2 μg/mL) stimulation for 1 h.
Suppression of MAPK signaling pathway by AE-ASL in LPS-stimulated RAW264.7 cells
Cells were pretreated with 0–100 μg/mL AE-ASL for 1 h and subsequently stimulated with LPS (0.2 μg/mL) for 1 h. Although phosphorylation of extracellular signal-regulated kinases (ERK1/2), c-Jun N-terminal kinase (JNK), and p38 MAPKs significantly increased after exposure to LPS, pretreatment with AE-ASL at 100 μg/mL effectively decreased their phosphorylation, by 44%, 55%, and 26%, respectively (Fig. 5). Meanwhile, the total expression levels of ERK1/2, JNK, and p38 were not significantly different among these groups. These results suggest that AE-ASL exerted its anti-inflammatory effects by inhibiting the activation of MAPKs during the LPS-induced inflammatory response.
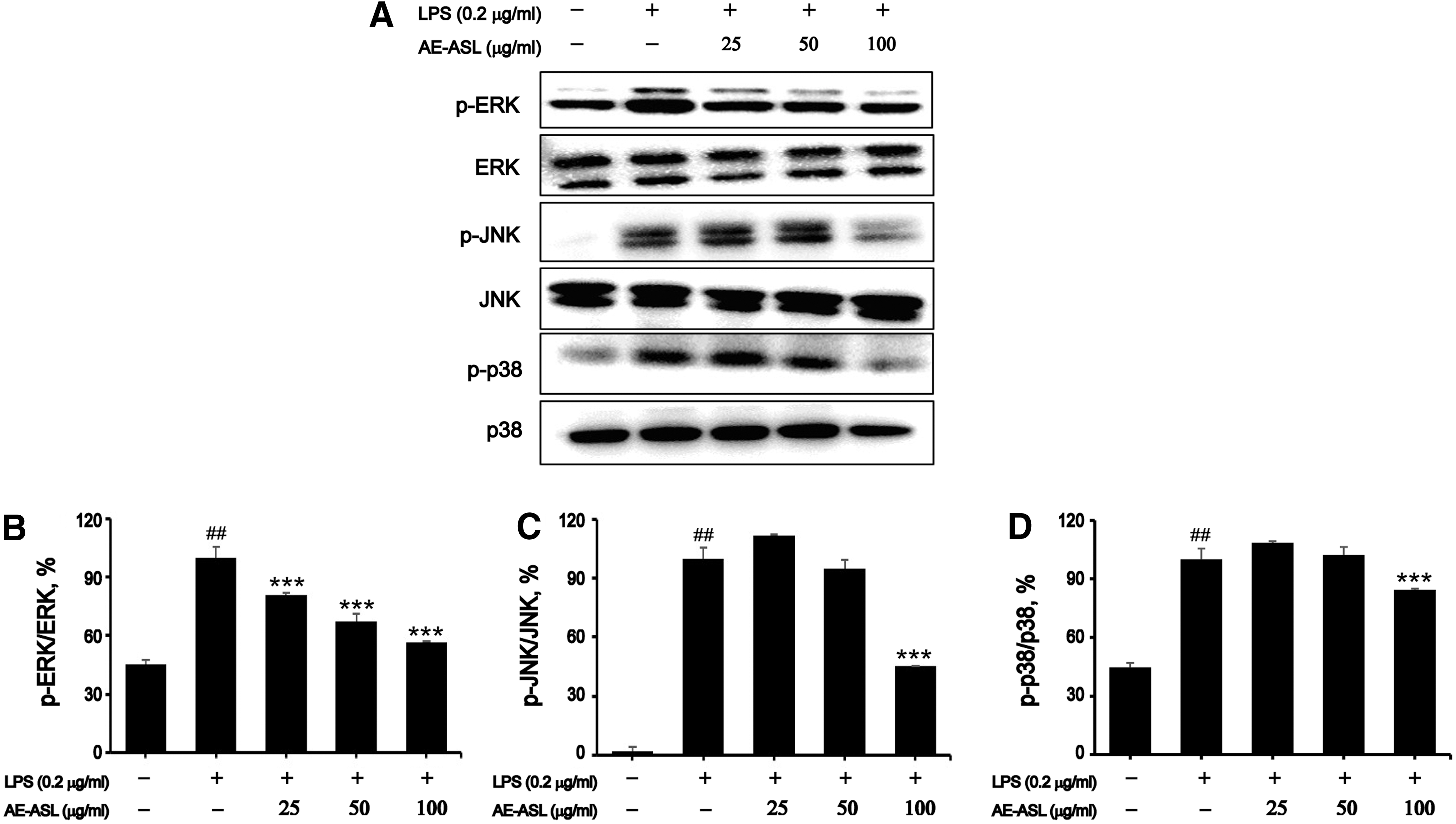
Effects of AE-ASL on LPS-induced phosphorylation of ERK, JNK, and p38 in RAW264.7 cells. Cells were pretreated with AE-ASL (25, 50, and 100 μg/mL) for 1 h, followed by LPS (0.2 μg/mL) stimulation for 1 h.
Effects of AE-ASL on carrageenan-induced rat paw edema
As shown in Figure 6B, AE-ASL (50, 100, or 200 mg/kg) and diclofenac sodium (positive control, 10 mg/kg), orally pre-administered 1 h before carrageenan injection, gradually increased the inhibition rate of the carrageenan-induced paw edema over a period of 24 h, compared with that in the untreated control group. At the high dose of AE-ASL (200 mg/kg), the inhibition of edema thickness was 27% and 40% at 2 h and 4 h after carrageenan injection, respectively; whereas diclofenac sodium showed 14% and 47% inhibition at 2 and 4 h, respectively. Therefore, the anti-inflammatory effect of 200 mg/kg AE-ASL was comparable to that of diclofenac sodium (10 mg/kg). In addition, the middle dose (100 mg/kg AE-ASL) increased the inhibition of the edema thickness from 1% to 31% between 2 and 4 h after carrageenan injection (Fig. 6B). The morphological characteristics are shown in Figure 6A.
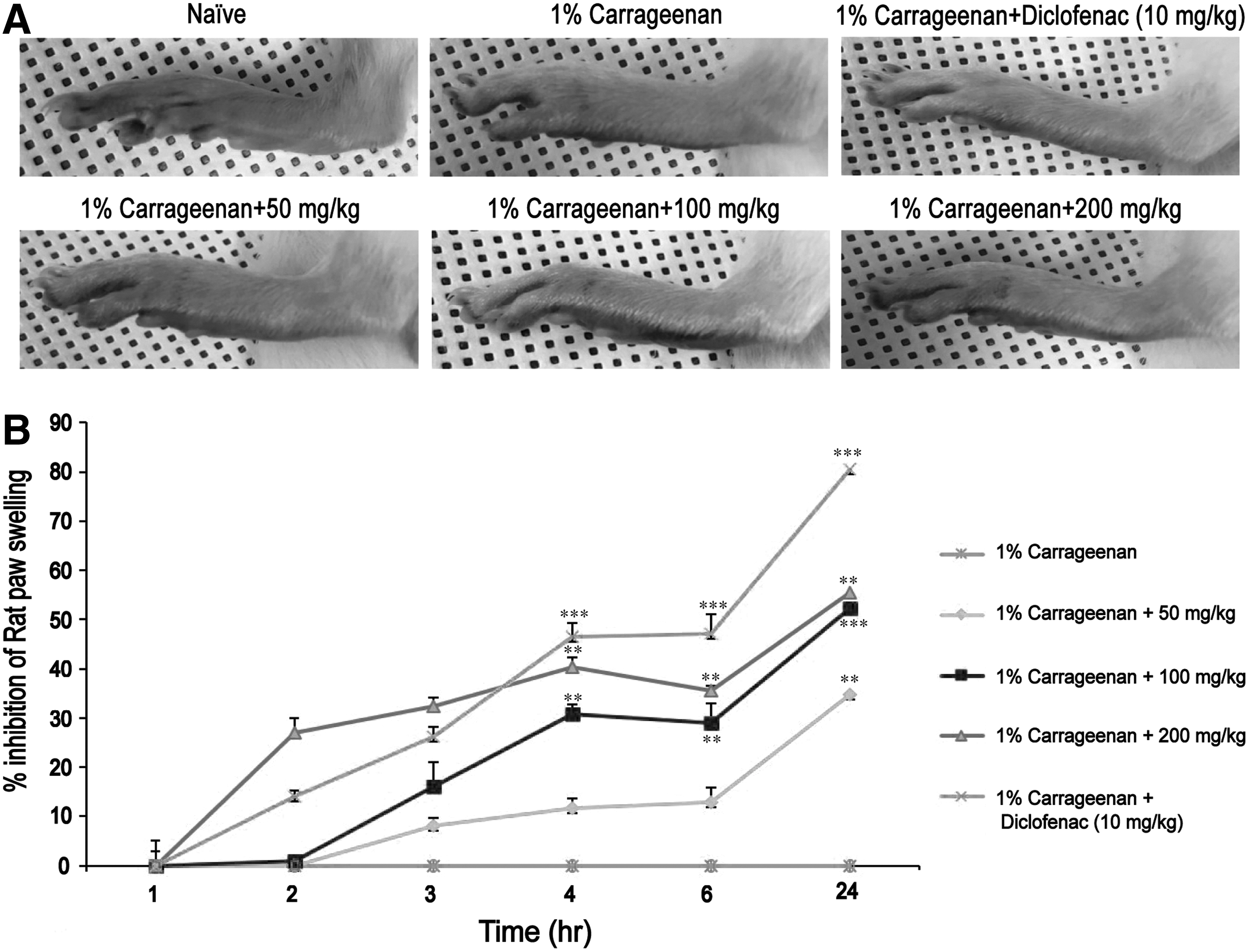
Effects of AE-ASL on carrageenan-induced left hind paw edema. Paw edema was induced by a subcutaneous injection of a 1% carrageenan solution (100 μL/animal) into the left hind paw 1 h after oral administration of AE-ASL (50, 100, and 200 mg/kg) or diclofenac sodium (10 mg/kg).
Identification of AE-ASL
The major components of the extract were separated and analyzed by using preparative HPLC, and three components with a retention time of 8.47, 11.46, and 12.60 min were detected at 320 nm wavelength (Fig. 7A). The mass spectrometric analysis of these fraction was performed by HPLC-ESI-TOF/MS under the same conditions; as a result, each peak with a retention time of 4.75, 7.20, and 8.01 min was detected at 320 nm wavelength (data not shown). As shown Figure 7B, compound 1 is predicted to be a chlorogenic acid having a molecular weight (M.W.) of 354.0951 and a formula of C16H18O9, compound 2 having an M.W. of 48.1006 and a formula of C21H20O11 is shown as a cymaroside, and compound 3 having an M.W. of 534.1010 and a formula of C24H22O14 is expected to be luteolin-7-O-malonylgluces.
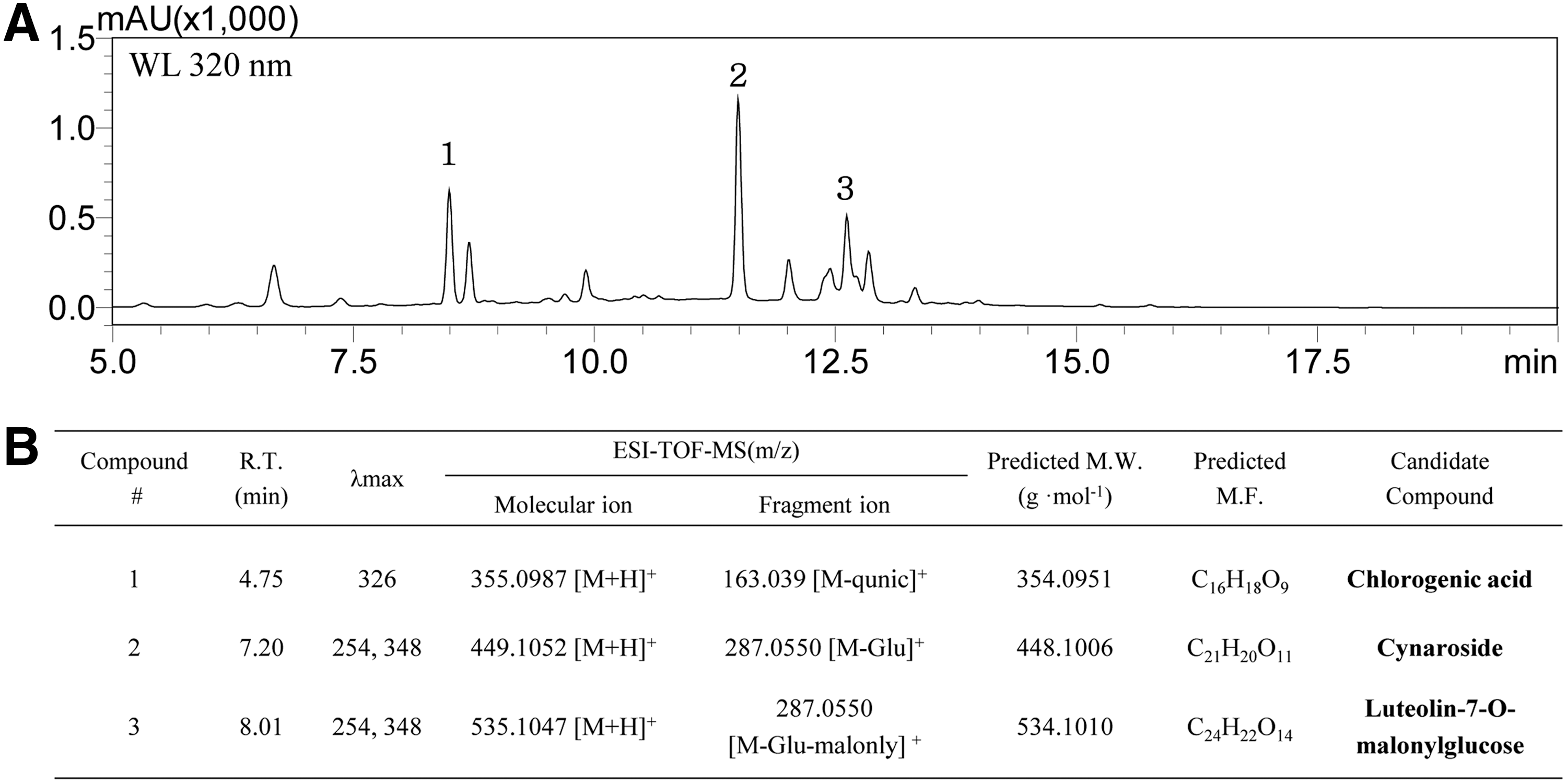
Identification of AE-ASL.
Discussion
Because drugs, including anti-inflammatory drugs, cause serious side effects, interest has increased in traditional herbal medicines having bioactive compounds. A. sylvestris has long been used as a medicinal plant in Korean traditional medicine, and it has been reported that aerial parts of this plant show antioxidant effects, whereas dried roots show antitumor activity by inducing the cell cycle arrest and apoptosis in vitro and in vivo. 14,19,20,24 However, physiologically active functions of AE-ASL are not yet known. Thus, we investigated the anti-inflammatory effects of AE-ASL in LPS-stimulated RAW264.7 cells and in a carrageenan-induced paw edema animal model.
It is well known that excessive production of NO and PGE2 by iNOS and COX-2, respectively, promotes the inflammation, acting synergistically with other inflammatory mediators. 25,26 Therefore, suppression of these inflammatory mediators may be an attractive strategy for the development of anti-inflammatory drugs. Consequently, numerous studies have been conducted to evaluate the inhibitory effects of natural products on the expression of inflammatory mediators. 27 –29 In this study, we found that AE-ASL significantly inhibited the LPS-induced NO and PGE2 production by suppressing iNOS and COX-2 protein and mRNA expression in a dose-dependent manner. We also found that pretreatment with AE-ASL decreased the expression of LPS-induced proinflammatory cytokines, TNFα, IL-1β, and IL-6, in macrophages. These findings agree with the results of other reports, which investigated the anti-inflammatory effects of various plant extracts, 27 –30 and support our hypothesis that AE-ASL has anti-inflammatory activity.
To further clarify the anti-inflammatory mechanism of AE-ASL, we investigated its effects on the activation of the NF-κB and MAPK signaling pathways in LPS-stimulated macrophages. These signaling pathways are well known to regulate the expression of numerous genes associated with the immunity, such as those encoding iNOS, COX-2, TNFα, IL-1β, and IL-6. 31,32 Our results showed that pretreatment with AE-ASL suppressed the translocation of the NF-κB p65 subunit from the cytoplasm to the nucleus via inhibition of the LPS-induced phosphorylation and degradation of IκBα. In addition, pretreatment with AE-ASL inhibited the LPS-induced phosphorylation of ERK, JNK, and p38 MAPKs. We suggest that the NF-κB and MAPK signaling pathways may mediate the AE-ASL inhibition of proinflammatory mediators and cytokines in LPS-stimulated RAW264.7 cells, which agrees with the data from previous studies. 27 –32
Carrageenan is a strong stimulant of proinflammatory mediators, and the inflammatory responses induced by carrageenan occur as a biphasic process. 33 The early phase (2 h after carrageenan injection) is mainly mediated by the release of histamine and serotonin; whereas the late phase (the first 3–4 h) is mediated by the release of bradykinin, TNFα, and leukotrienes and is sustained by PGE2 and COX-2. 33,34 Thus, most conventional anti-inflammatory drugs target the late phase. 35 Our in vivo study showed that orally administered AE-ASL inhibited the carrageenan-induced paw edema in a dose-dependent manner for up to 4 h after the carrageenan injection, and 200 mg/kg AE-ASL was especially effective in the early phase of the carrageenan-induced inflammatory response. These results suggest that the high dose of AE-ASL exhibited anti-inflammatory effects by suppressing the release of the proinflammatory mediators, histamine and serotonin, at the initial stage.
HPLC and HPLC-ESI-TOF/MS analysis identified phenolic compounds (chlorogenic acid, cynaroside, and luteolin-7-o-malonylgluces) as the major bio-active components in the extract. Dall'Acqua et al. reported that chlorogenic acid and cynaroside are the active components with antioxidant activity in the methanol extract of A. sylvestris aerial parts. 20 In addition, chlorogenic acid and cynaroside were reported to exert anti-inflammatory effects in mice with D-gal-induced chronic liver injury and in a carrageenan-induced paw edema rat model. 36,37
This study suggests that AE-ASL has anti-inflammatory activity and may have potential in anti-inflammatory therapy.
Footnotes
Acknowledgments
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Agri-Bio Industry Technology Development Program (or Project), funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (317003-4).
Author Disclosure Statement
No competing financial interests exist.