
Abstract
Tannins are a type of polyphenols found in several fruits such as grapes and berries, and nuts such as aronias and acorns. Both hydrolyzable tannins and condensed tannins are referred to as tannins. Among the hydrolyzable tannins, gallotannin has a strong antioxidative property and is known to protect the skin by inhibiting the precursors of elastolytic enzymes. However, its mechanism of protection against ultraviolet B (UVB) damage in human fibroblasts and keratinocytes has not yet been elucidated. In this study, we investigate the antioxidant and antiaging effect of gallotannin on UVB-irradiated human cells by studying its effect on extracellular signal-regulated kinases/c-Jun N-terminal kinases (EKRs/JNKs) signaling related to cell growth and differentiation/stress apoptosis. The results showed that gallotannin improved collagen synthesis, reduced metalloproteinase-1 (MMP-1) expression in a dose-dependent manner, and downregulated MMP-1 levels through the ERK/JNK signaling pathway in UVB-irradiated human cells. Gallotannin also increased glutathione but did not increase transforming growth factor beta 1, which induces fibrosis. We propose that gallotannin is a novel agent for protection against UVB, and acts as an antiaging agent that can be used in food, pharmaceuticals, and cosmetics.
Introduction
U
Materials and Methods
Cell culture and reagents
Normal human dermal fibroblast (NHDF) cells were provided by MCTT (Seoul, Republic of Korea) and HaCaT cells were purchased from ATCC (Koram Biotech Corp., Seoul, Republic of Korea). Cells were cultured using DMEM with 10% heat-inactivated fetal bovine serum and 100 U/mL to 100 μg/mL of penicillin–streptomycin. The cells were maintained in a humidified incubator with 5% CO2 at 37°C. The doubling times for cells were about 3–4 days. Gallotannins, propyl gallate, and dodecyl gallate were used test samples (5–50 μM) and were purchased from Sigma.
Determination of cell viability
Cell growth experiments were performed using the MTT [3-(4,5-methylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; Sigma] assay as described previously. 34,35 NHDF and HaCaT cells were seeded at a density of 4 × 103 cells/well in 96-well plates, respectively. The cells were grown overnight, and then exposed to different concentrations of samples. After 3 days, MTT was added (10/100 μL media) for 4 h. Formazan products were solubilized with dimethyl sulfoxide, and then shaken for 15 min. Optical density was measured at 570 nm by ELISA plate reader.
MMP-1 expression by ERK signal pathway
HaCaT and NHDF cells were seeded into a six-well plate at a density of 1 × 106 cells/well, respectively, and cultured overnight. After 24 h, gallotannins were used to treat cells for 3 days. The cells were exposed to UVB light at a rate of 110 mJ/cm2 using UVB lamp (280–320 nm, Max 306 nm; Hansung Ultraviolet Co. Ltd., Seongnam, Korea) before 24 h of harvest. ERK phosphorylation inhibitor (PD98059; Sigma, St. Louise, MO, USA) and Jun N-terminal kinases (JNK) phosphorylation inhibitor (SP600125; Sigma) of 20 μM were added before 2 h of harvest, respectively. Harvested cell supernatants were used in experiments. The level of MMP-1 release was assayed by ELISA according to the manufacturer's instructions (Duoset ELISA Development System; R&D System, Minneapolis, MN, USA), and MMP-1 protein expression was detected by Western blot analysis (ChemiDoc XRS System; Bio-Rad, Hercules, CA, USA). The harvested HaCaT cells were washed with phosphate-buffered saline (PBS) which is a buffer solution commonly used in biological research, and then were lysed in ice-cold RIPA buffer containing the protease inhibitors aprotinin (2 μg/mL) and phenylmethane sulfonyl fluoride (100 μg/mL) and 1% phosphatase inhibitor cocktail (Sigma). The cell supernatant was collected and the protein concentration was determined using the Lowry assay. 36 Aliquots of cell lysates were separated by SDS-PAGE (Mini-Protean TGX Gel; Bio-Rad), transferred to polyvinylidene difluoride membranes (Trans-Blot Turbo Transfer Pack; Bio-Rad), and then treated with the appropriate antibodies for Western blotting. These antibodies included rabbit anti-MMP-1 (Abcam, Cambridge, United Kingdom) and goat polyclonal antirabbit IgG (Santa Cruz Biotechnology).
Collagen synthesis
NHDF cells were seeded into a six-well plate at a density of 4 × 105 cells/well and cultured overnight. After 24 h, cells were incubated with test samples and the cells were exposed to UVB light at a rate of 110 mJ/cm2 using a UVB lamp before 24 h of harvest. Harvested cell supernatants were used in experiments. Collagen assay was performed using an indirect ELISA protocol. 37 Cell supernatants were coated onto a 96-well plate (100 μg/well) and incubated at 4°C for 6 days. Each well was then washed three times with 1 × PBST (polysorbate20 of 0.05% in PBS, pH 7.2–7.4) which is a buffer solution commonly used in ELISA study, and then blocked with 1% bovine serum albumin in PBS (pH 7.4) at 37°C for 30 min. Each well was then washed three times with 1 × PBST. Then, 100 μL of primary antibodies solution (monoclonal mouse anticollagen; 1:1000; Sigma) was added and then the plate was incubated at 37°C for 1 h. After washing each well three times with 1 × PBST, 100 μL of secondary antibodies solution (antimouse IgG HRP; 1:1000; Abcam) was added; the plates were incubated at 37°C for 30 min. After washing three times with 1 × PBST, 100 μL of 3,3′,5,5′-tetramethylbenzidine solution was added, and the plates were incubated at 37°C for 30 min. To stop the reaction, 100 μL of 2 M H2SO4 was added, and then absorbance was measured at 450 nm (Spectramax Plus 384; Molecular Devices, Sunnyvale, CA, USA).
TGF-β1 expression
NHDF and HaCaT cells were seeded into a six-well plate at a density of 4 × 105cells/well, respectively, and cultured overnight. Before irradiation, when cells were 80% confluent pretreated with samples. After 24 h, cells were incubated with test samples and the cells were exposed to UVB light at a rate of 110 mJ/cm2 using UVB lamp before 24 h of harvest. Harvested cell supernatants were used in experiments. Level of TGF-β1 was assayed by ELISA according to the manufacturer's instructions (Duoset ELISA Development System).
GSH level
HaCaT cells were seeded at a density of 2 × 107 cells/mL were seeded into a 100mm culture dish at a density of 2 × 107 cells/mL and cultured overnight. Before irradiation, when cells were 80% confluent, they were pretreated with sample. After 24 h, culture media were replaced with PBS of 1.5 mL. The cells were then exposed to UVB light at a rate of 110 mJ/cm2 using a UVB lamp. After 24 h of UVB irradiation, the cells were harvested. Cells were washed with 1 × PBS and then added to 5% 5-sulfosalicylic acid hydrate (Sigma) threefold dilution for cell pellets volume. Cell pellets were frozen and thawed twice using liquid nitrogen, then incubated at 2–8°C for 5 min. The cell supernatants obtained by centrifugation were used to test samples. The GSH level was measured as previously reported. 38 Then 200 μL of PBS, 20 μg/μL of sample, and 4 mg/mL of 5,5′-dithiobis (2-nitrobenzoic acid/methanol solvent; Sigma) were mixed and then incubated at room temperature for 15 min. Absorbance was measured at 412 nm.
Statistical analysis
All data are presented as means and standard deviation from three or more replicates. Statistical comparisons between treatments were performed using the Student's t-test. Differences with P values < .05 were considered statistically significant. The results are averages of three independent experiments, and the data are expressed as mean ± SD. #P < .1, *P < .05, **P < .01, and ***P < .001 indicate statistically significant differences from the vehicle control.
Results
MMP-1 expression through ERK and JNK signaling pathway
Gallotannin significantly inhibited UVB-induced MMP-1 levels in both HaCaT and NHDF cells (HaCaT cells, 52% in 50 μM [P < .01]; NHDF cells, 33% in 50 μM [P < .001], refer to Fig. 1). The MMP-1 gene expression was also significantly inhibited by gallotannin through activation of ERK/JNK pathway in UVB-induced HaCaT cells (Fig. 2).
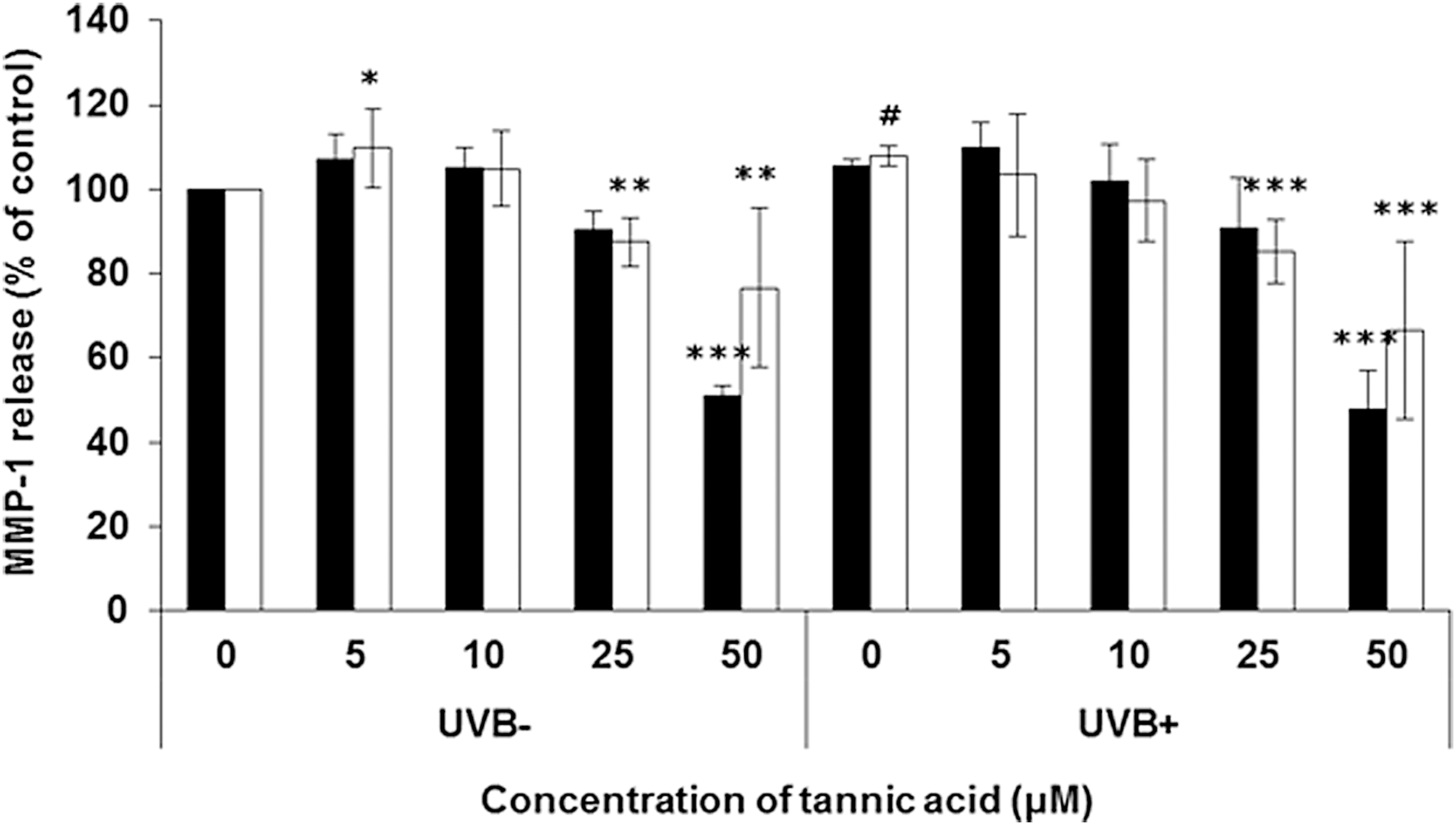
Effect of gallotannin on MMP-1 release in UVB-induced HaCaT (black bar) and NHDF (white bar) cells. Gallotannin of various concentrations was treated for 3 days and the cells were exposed to UVB (110 mJ/cm2) before 24 h of harvest. Assay was evaluated by ELISA protocol. The results are averages of three independent experiments, and the data are expressed as mean ± SD. #P < .1, *P < .05, **P < .01, and ***P < .001 indicate statistically significant differences from the vehicle control. HaCaT, human epidermal keratinocyte; MMP-1, metalloproteinase-1; NHDF, normal human dermal fibroblast; SD, standard deviation; UVB, ultraviolet b.
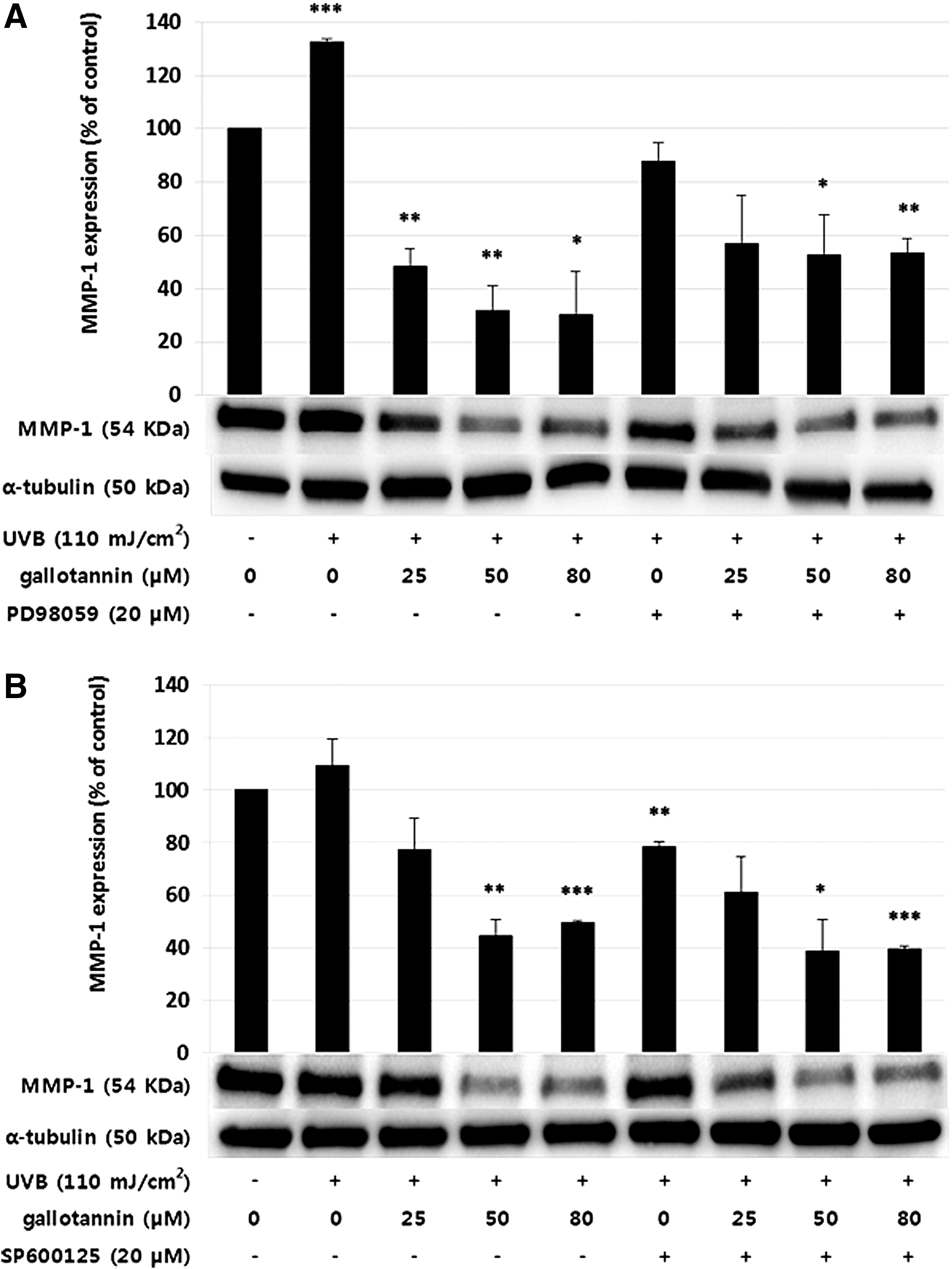
Effect of gallotannin on ERK
ERK phosphorylation is activated by UV exposure
35
and suppressed by the inhibitor, PD98059. Activated ERK was significantly decreased further by treatment with 50 μM of gallotannin. The ERK phosphorylation decreased considerably from the PD98059 treatment for 2 h before harvest and was also significantly decreased by the gallotannin treatment (P < .05). The ERK phosphorylation was decreased by treatment with 50 μM gallotannin but not by 5 μM of the same (Supplementary Table S1 and Supplementary Figs. S1 and S2; Supplementary Data are available online at
Correlation of collagen synthesis and collagenase inhibition with TGF-β1
Gallotannin improved collagen synthesis compared with vehicle control. In UVB-irradiated cells, collagen levels were restored by 5–50 μM of gallotannin. However, there was no effect on the collagen synthesis in the propyl gallate and dodecyl gallate treatment. The two compounds were even decreased in normal conditions and collagen levels did not improve in UVB-irradiated cells (Fig. 3). The hallmarks of fibrosis or antiaging increased, including collagen synthesis and inhibition of MMP-1. 39 –41 We observed that gallotannin induced collagen synthesis and reduced MMP-1 levels. We also tried to estimate inducible fibrosis through TGF-β1 gene expression. If the gallotannin regulates collagen synthesis but does not increase TGF-β1 in UVB-induced human cells, it will be a novel antiaging agent against UVB. Fortunately, the fibrosis inducer, TGF-β1, did not increase (Fig. 4).
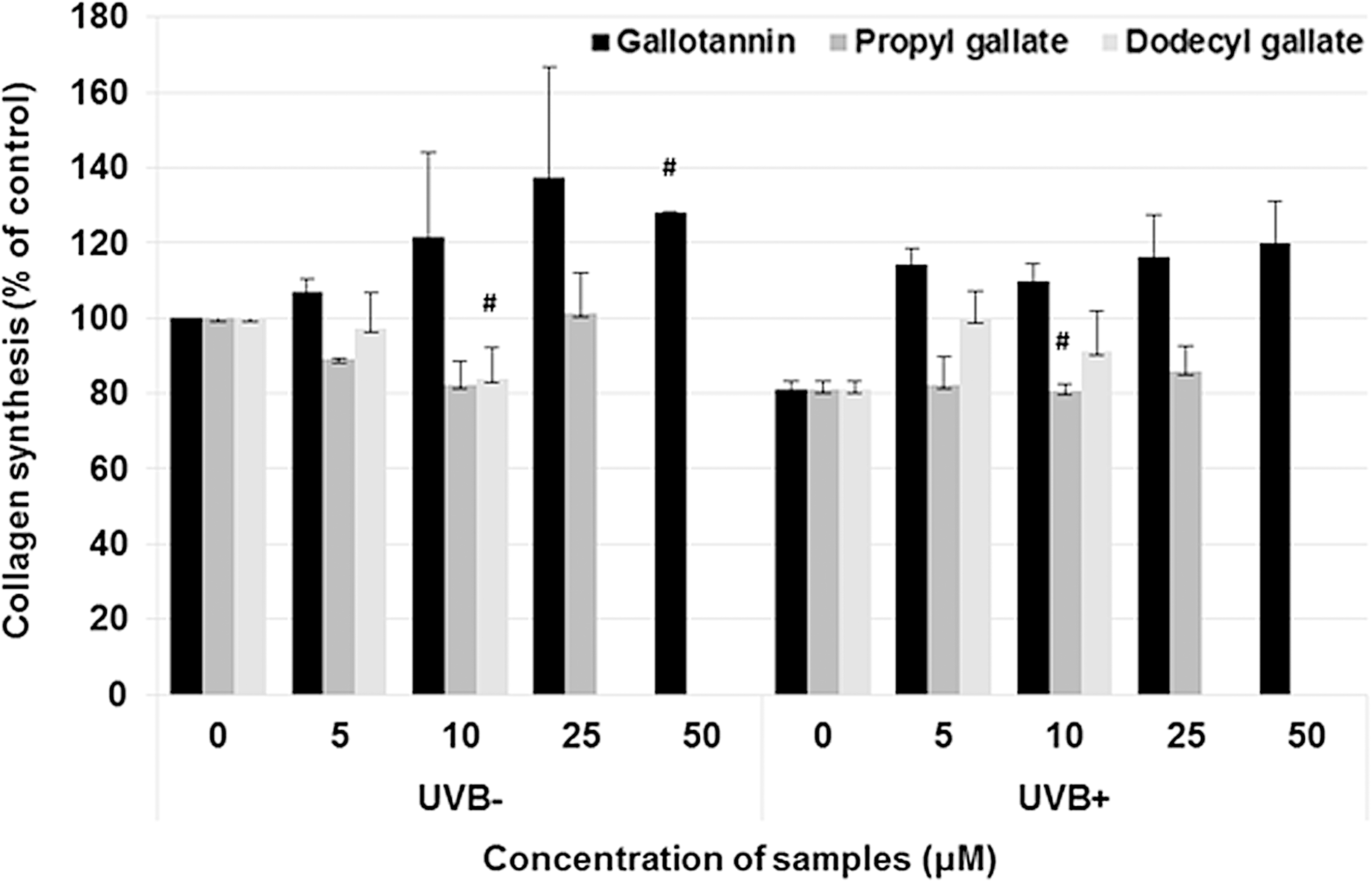
Effect of gallotannin on collagen synthesis in UVB-induced NHDF cells. Gallotannin of various concentrations was used to treat cells for 3 days. The cells were exposed to UVB (110 mJ/cm2) 24 h before harvest. Assay was evaluated by ELISA protocol. The results are averages of three independent experiments, and the data are expressed as mean ± SD. #P < .1 indicates statistically significant differences from the vehicle control. Black bar, gallotannin; gray bar, propyl gallate; and light gray, dodecyl gallate.

Effect of gallotannin on TGF-β1 in UVB-induced HaCaT
GSH level
We used GSH levels as antioxidant index 42,43 and gallotannin treatment resulted in significantly higher GSH levels when compared with that of propyl gallate and dodecyl gallate. Gallotannin increased GSH amount in a dosage-dependent manner. GSH levels were at 144% ± 0.7% in 25 μM gallotannin and 155% ± 9.8% in 50 μM gallotannin (Fig. 5). In contrast, propyl gallate and dodecyl gallate had no significant effect.
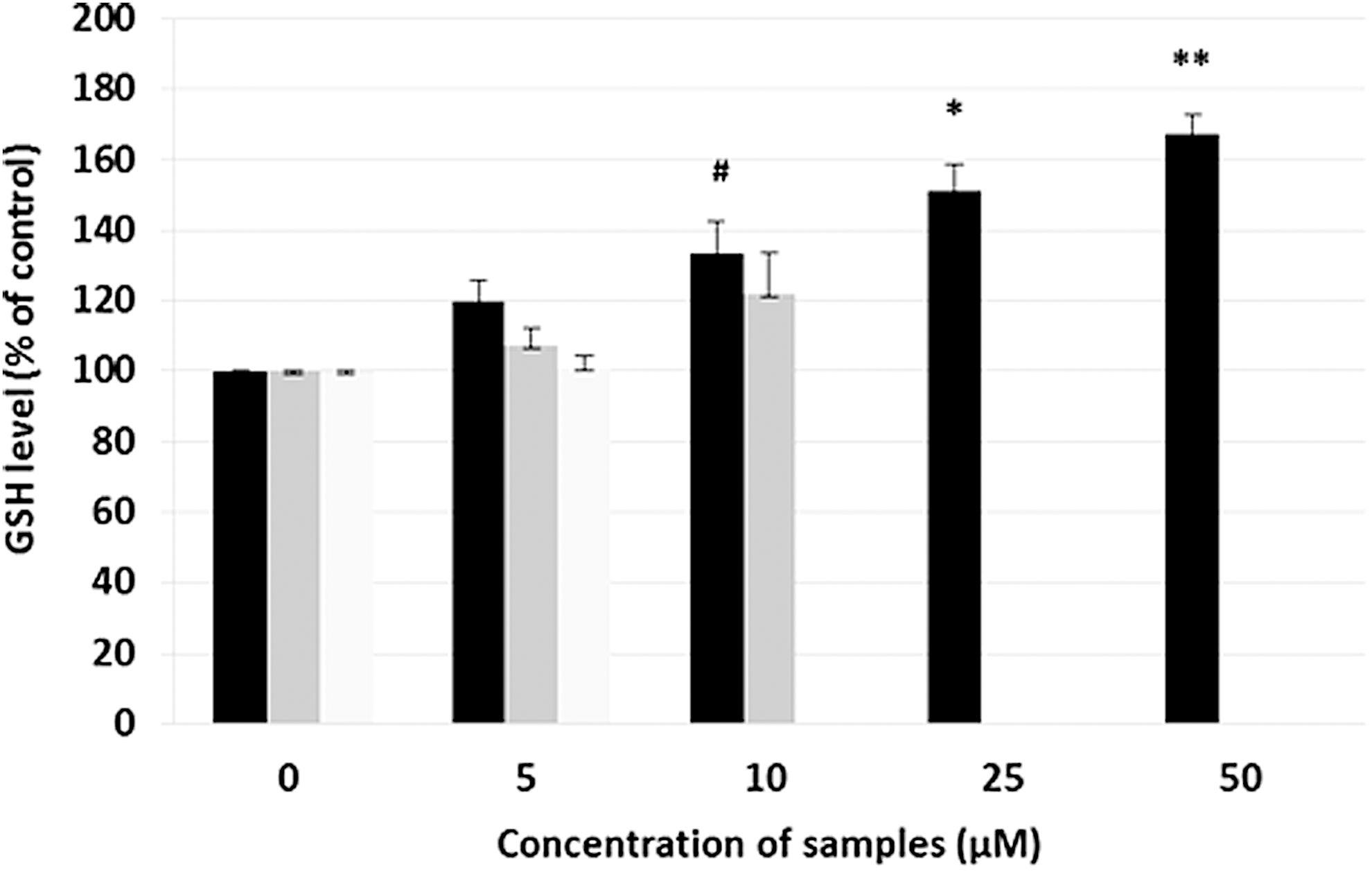
Effect of gallotannin on GSH level in UVB-induced HaCaT cells. Gallotannin of various concentrations was used to treat cells for 48 h and the cells were exposed to UVB (110 mJ/cm2) before 24 h of harvest. The results are averages of three independent experiments, and the data are expressed as mean ± SD. #P < .1, *P < .05, and **P < .01 indicate statistically significant differences from the vehicle control. Black bar, gallotannin; gray bar, propyl gallate; and light gray, dodecyl gallate. Propyl gallate exhibited cell cytotoxicity at >25 μM and dodecyl gallate showed cell cytotoxicity at >10 μM in HaCaT. GSH, glutathione.
Cell viability
Gallotannin resulted in a cell viability of >80% in NHDF and HaCaT cells. Propyl gallate exhibited toxicity at >25 μM in HaCaT cells and dodecyl gallate showed cell cytotoxicity at >10 μM in HaCaT cells and >25 μM in NHDF cells (Fig. 6).

Cell viability of gallotannin, propyl gallate, and dodecyl gallate in HaCaT
Discussion
We observed that UVB irradiation induces MMP-1 protein production in human keratinocytes and fibroblast cells using Western blotting analysis and ELISA. It has been reported that MMP-1 levels increase and that collagen synthesis decreases in photoaged skin. 44 –47 In this study, we examined the effects of gallotannin on UVB-induced cells.
To confirm whether MMP-1 regulates ERK/JNK signal transduction, we treated HaCaT cells with ERK/JNK inhibitors at a concentration of 20 μM for 2 h and analyzed the effect of gallotannin. ERK/JNK are involved related to cell growth and differentiation/cytokines, UV irradiation, heat shock, osmotic stress and apoptosis. 15,48 The observed effect of gallotannin was more pronounced in JNK signaling than in ERK signaling. These results indicate that gallotannins regulate the UVB-induced MMP-1 levels through the MAPK signaling pathway, which is related to aging, 49 melanogenesis, or inflammatory response. 50
The physiological activity of gallotannin on UVB-induced human keratinocytes has not been examined in recent studies. However, we were able to determine that gallotannin regulates MMP-1 through ERK and JNK pathways in the keratinocyte cell line, HaCaT. The gallotannin derivatives, propyl gallate and dodecyl gallate, have been reported to not only block prolyl hydroxylation activity and inhibit oxidative stress but also have anticancer activity. 51 –53 In contrast, it has been reported that propyl gallate, octyl gallate, and dodecyl gallate cause dermatitis. 54 Even though they have antioxidant effects, we could not measure their antioxidant or antiaging activities since they induce cytotoxicity in human cells (Fig. 6). However, gallotannin showed superior cell viability when compared with propyl gallate and dodecyl gallate. In addition, gallotannin displayed a distinct antiaging effect.
Just before UVB irradiation, the culture medium of cells was replaced with PBS to avoid the generation of toxic photoproducts by the medium. The UVB dose was selected based on previous reports 45,46 and the results on cell viability (Supplementary Tables S2–S4). Under these conditions, the cells used in the experiment did not exhibit a loss in cell viability.
TGF-β1 is an important cytokine involved in cell functions such as inhibition of cell growth, cell differentiation, apoptosis, and immune response. TGF-β1 is overexpressed in keratinocytes in several immune-related diseases and occurs in skin disorders by the activation Smad through TGF-β1. 14,16 In TGF-β1-induced hepatic stellate cells, tannins exhibited antiproliferation and anticytotoxic effects (antifibrosis). 55 It has also been reported that tannic acid ameliorates carbon tetrachloride-induced liver fibrosis in vivo and in vitro. 56 As a result, we predicted that tannins may play a role as antiaging agent in human keratinocytes. In this study, we observed that gallotannin regulates ERK signaling and suppresses MMP-1 through ERK signals in human keratinocyte cells. Gallotannin accelerated collagen synthesis but did not regulate TGF-β1. As a result, we could presume that gallotannin did not promote fibrosis. Therefore, gallotannin can serve as a novel substance for antiaging and accelerate wound healing but not fibrosis.
It has been previously reported that liver cirrhosis and liver tumors are induced by prolonged subcutaneous administration of tannic acid solutions. 57,58 However, its antifibrosis activity has also been reported in recent studies. 55,56 Whether tannins induce fibrosis or not, we have clearly confirmed in this study that gallotannin acts as an ERK/JNK inhibitor and regulates signal transduction in the ERK/JNK signaling pathway. Ultimately, gallotannin induced collagen synthesis and inhibited MMP-1. In addition, it did not mediate the expression of TGF-β1, which causes fibrosis. It also has an antioxidant property, thereby elevating GSH levels. These results demonstrate that gallotannin might be a potential novel antiwrinkle agent. Further studies have been scheduled to evaluate the effect of gallotannin on mRNA levels for antiaging regulation and to measure its effect on anti-inflammatory regulation. Although gallotannin is known to be a fibrosis inducer, these results suggest that gallotannin can serve as a potential candidate for the regulation of antiaging or wound healing in industries related to food, drugs, or cosmetics.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.