
Abstract
This study investigated the effects of Portulaca oleracea L. extract on glucose uptake in 3T3-L1 adipocytes. P. oleracea extract (POE) markedly enhanced glucose uptake, which was caused by increased GLUT4 expression at the plasma membrane (PM) in 3T3-L1 adipocytes. This increase in PM-GLUT4 expression was associated with insulin receptor substrate-1 (IRS-1) phosphorylation, phosphatidylinositol 3-kinase (PI3K) activation, and Akt phosphorylation, and finally, enhanced intracellular glucose uptake. POE was not associated with protein kinase C (PKC)λ/ζ phosphorylation in the insulin signaling pathway, but did promote 5′-AMP-activated kinase (AMPK) phosphorylation. Increased glucose uptake through POE was inhibited through treating with the PI3K inhibitor or AMPK inhibitor in 3T3-L1 adipocytes. This result suggested that POE may enhance glucose uptake by stimulating GLUT4 translocation to the PM through activating the PI3K and AMPK pathway in 3T3-L1 adipocytes.
Introduction
D
Portulaca oleracea is an herbaceous annual plant that is distributed extensively in Korea and China. It is known as a nutritious herb and traditionally makes use of it as medicinal food. 3,4 Previous studies found that P. oleracea extract (POE) exerted biological activities, such as antioxidant, anti-inflammatory, antiatherogenic, antiviral, antitumor, antidiabetic, and neuroprotective activities. 5 –10 However, no study has yet investigated whether POE enhances glucose uptake through insulin signaling pathway. Therefore, this research investigated the efficacy of POE on glucose uptake in 3T3-L1 adipocytes.
Materials and Methods
Materials
P. oleracea was purchased from Hongcheon Hyosung Food, Inc. (Hongcheon, Korea). It was carefully washed with pure water and freeze dried (Samwon Freezing Engineering Co., Busan, Korea). They were lyophilized and pulverized to a powder form. The sample was extracted three times with decuple of 80% ethanol for 3 days at room temperature. The POE was concentrated in a rotary vacuum evaporator. After the extract was thoroughly dried to remove the solvent, the dried extracts were stored in a cryogenic freezer (−80°C).
Cell culture and adipocyte differentiation
Mouse 3T3-L1 preadipocyte cells were purchased from Korean Cell Line Bank (KCLB, Seoul, Korea) and were grown in Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine serum (FBS) at 37°C in a 5% CO2 atmosphere. These cells were induced to differentiate into adipocytes in the following manner. Briefly, 2 days after confluence, the medium was changed to 10% fetal calf serum DMEM supplemented with 0.5 mM isobutylmethylxanthine, 1 μM dexamethasone, and 10 μg/mL insulin. Subsequently, cells were treated with 10 μg/mL insulin in 10% FBS DMEM for an additional 24 h and then maintained with 10% FBS fresh media every other day for 8 days. With this protocol, more than 80% of the preadipocytes differentiate into adipocytes. 11
2-Deoxyglucose uptake assay
Glucose uptake assay using a 2-[N-(7-Nitrobenz-2-oxa-1,3-diazol-4-yl)Amino]-2-deoxyglucose (2-NBDG) was performed as previously described with some adjustments. Around 1 × 104 3T3-L1 adipocytes were seeded into each well of 96-well plates with POE, POE +10 μM compound C (CC, an 5′-AMP-activated kinase [AMPK] inhibitor), or POE +20 μM Wortmannin (WM, a PI3K inhibitor) for 24 h. After incubation, cells were left untreated or were stimulated with 100 nM insulin for 20 min at 37°C in Kreb's ringer phosphate buffer (128 mM NaCl, 4.7 mM KCl, 1.25 mM CaCl2, 1.25 mM MgSO4, and 10 mM NaPO4, pH 7.4). Glucose uptake was initiated by the addition of 10 μM 2-NBDG to each well. After 1 h, the supernatant was removed. Uptake of 2-NBDG was measured by a Multilabel Counter (Perkin Elmer, MA, USA) set at an excitation wavelength of 485 nm and emission wavelength of 535 nm.
Western blot analysis
To extract total protein from adipocytes, the cells were washed twice with ice-cold phosphate-buffered saline and harvested in lysis buffer (RIPA, 50 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1% Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], 1 mM phenylmethylsulfonyl fluoride [PMSF], 10 μg/mL aprotinin, 10 μg/mL leupeptin, 0.1 mM sodium orthovanadate [pH 7.4]). After sonication and centrifugation at 13,000 g for 30 min at 4°C, the protein content of the resulting supernatant was determined using a BCA Protein Assay Kit. The lysate containing 20 μg of protein was subjected to electrophoresis with 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Separated proteins were transferred electrophoretically to a pure nitrocellulose membrane, blocked with 5% skimmed milk solution for 1 h, and then incubated with the appropriate primary antibody (1:1000; Abcam, Cambridge, United Kingdom;) overnight at 4°C. After washing, the blots were incubated with goat anti-rabbit or goat anti-mouse IgG horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. Each antigen–antibody complex was visualized using ECL Western Blotting Detection Reagents and detected by chemiluminescence with a LAS-1000 plus instrument (FUJIFILM, Tokyo, Japan). Band densities were determined by an image analyzer (Multi Gauge V3.1; FUJIFILM Corporation, Valhalla, NY, USA) and normalized to β-actin for total protein and nuclear protein.
Isolation of PMs from 3T3-L1 adipocytes
3T3-L1 adipocytes were homogenized by sonication for 5 min at 3 kHz/130 W (UCD-130TM; Cosmo Bio Co., Tokyo, Japan) in ice-cold HES buffer (0.02 M HEPES, 0.25 M sucrose, and 2 mM EGTA; pH 7.4) and centrifuged at 700 g for 7 min to remove nonhomogenized cellular debris and nuclei from the homogenate. The harvested supernatant was further centrifuged at 760 g for 10 min to remove mitochondria. It was recentrifuged at 35,000 g for 60 min, and the resulting pellet was used as the PM fraction of the adipocytes. The supernatant was used as the cytosol fraction. These membrane and cytosol fractions were subjected to western blotting to detect GLUT4 protein. The protein concentrations of the cytosolic fraction and membrane pellet were quantified using the BCA Protein Assay Kit.
Statistical analyses
Data represent the mean ± standard error of three experiments. Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by post hoc Duncan's multiple-range tests. Statistical analyses were performed using the SAS 9.1 software (SAS Institute, Inc., Cary, NC, USA).
Results
Effect of POE on 2-deoxyglucose uptake
To examine the effect of POE on glucose uptake, we conducted a 2-deoxyglucose uptake test using 3T3-L1 adipocytes. Uptake of 2-deoxyglucose significantly increased upon treatment with POE in 3T3-L1 adipocytes (Fig. 1). When the cells were treated with 25, 50, and 100 μg/mL POE, glucose uptake was dose dependently increased 1.23-, 1.45-, and 1.67-fold compared with that of untreated control cells. The results showed that POE enhanced glucose uptake in 3T3-L1 adipocytes.
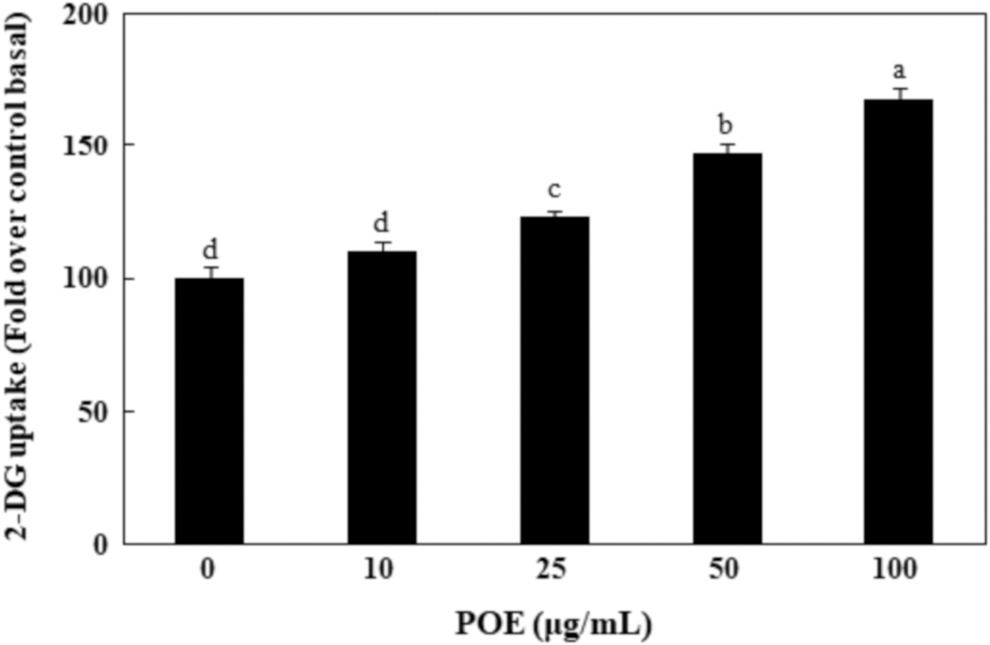
Effect of POE on glucose uptake in 3T3-L1 adipocytes. Differentiated 3T3-L1 adipocytes were treated with various concentrations of POE for 24 h before a 2-DG uptake assay. Each value is expressed as mean ± SD of triplicate experiments. a–dValues with different superscript letters were significantly different at P < .05 as determined by Duncan's multiple range test. 2-DG, 2-deoxyglucose; POE, Portulaca oleracea extract; SD, standard deviation.
Effect of POE on PI3K/Akt pathway expression
To establish the mechanism by which POE enhanced glucose uptake, the insulin signaling pathway was examined by determining the expression of IRS-1, PI3K, and AKT or PKCλ/ζ proteins. As shown in Figure 2, treatment with POE significantly increased IRS-1 phosphorylation and PI3K activation. At 100 μg/mL, POE treatment increased IRS-1 phosphorylation and PI3K activation to 151% and 166% of control values, respectively. Treatment of 100 μg/mL POE also markedly increased AKT phosphorylation to 149% of the value in untreated control cells. However, POE treatment did not increase PKCλ/ζ phosphorylation (Fig. 3). These results showed that POE promoted the phosphorylation of IRS-1 and AKT, and PI3K activation in the insulin signaling pathway.
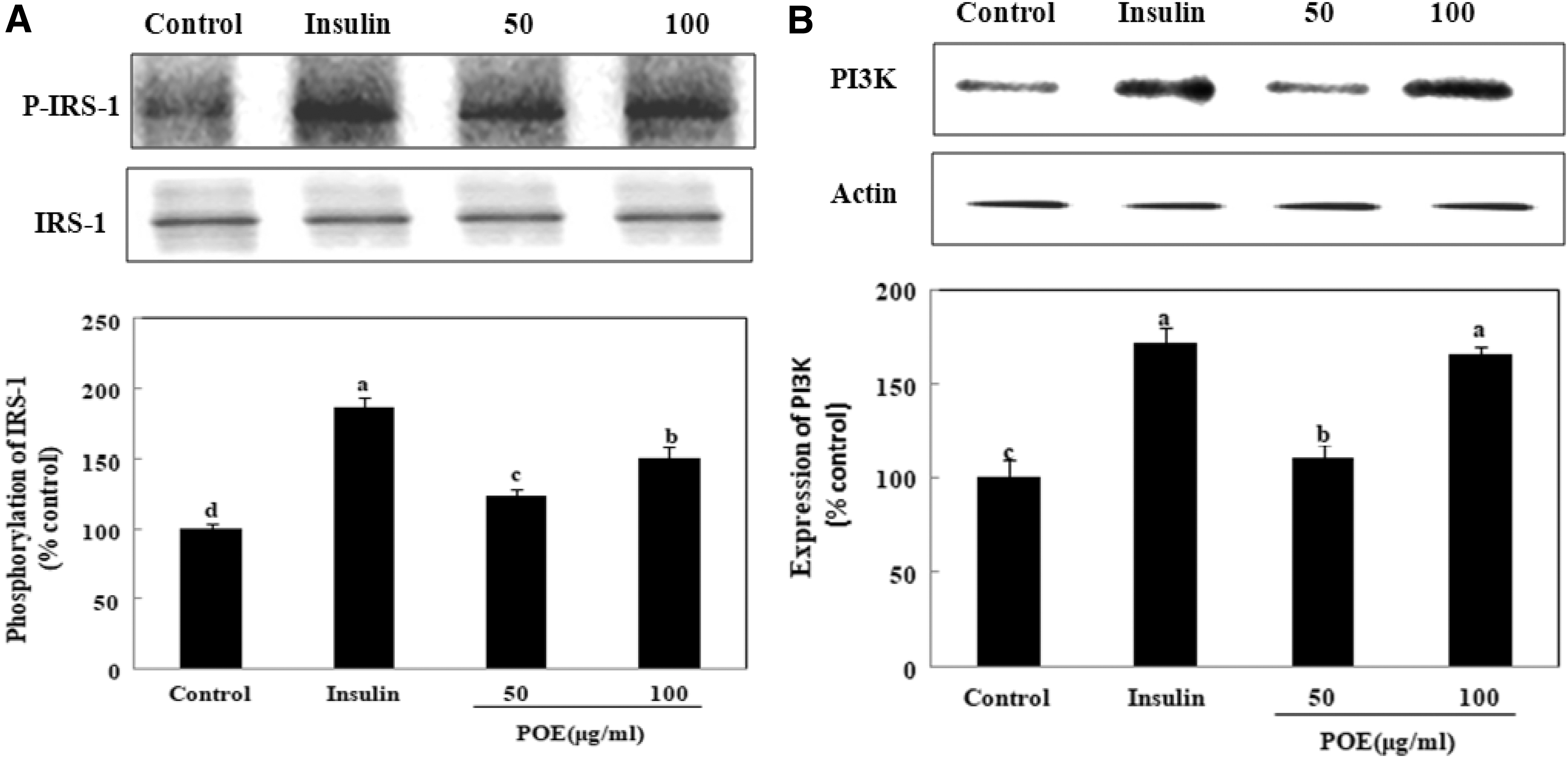
Effects of POE on IRS-1 and PI3K expression in 3T3-L1 adipocytes. Differentiated adipocytes were incubated with 50 or 100 μg POE extract or 100 nM insulin for 24 h, and then cells were lysed and subjected to immunoblot analysis.
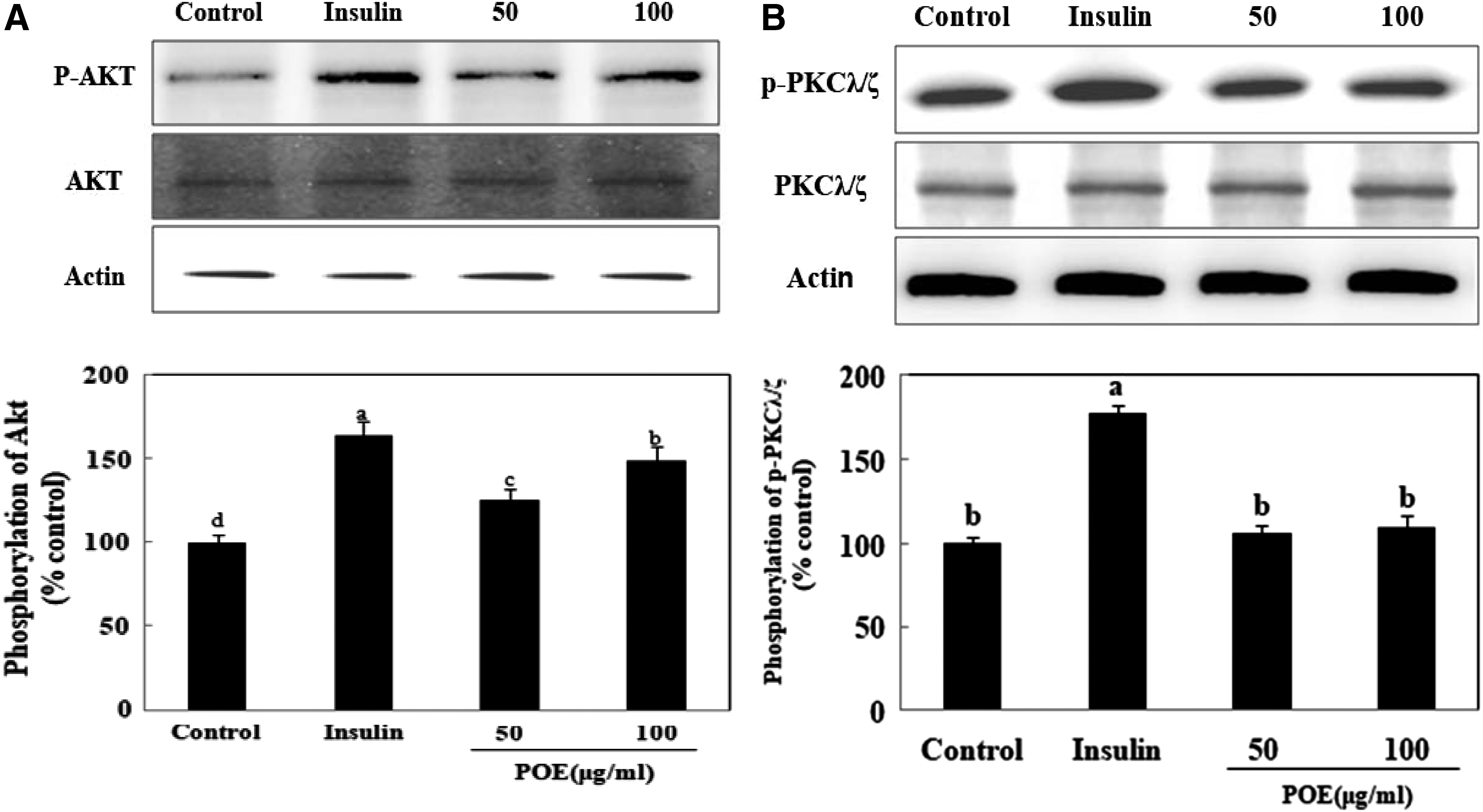
Effects of POE on Akt and PKCλ/ζ expression in 3T3-L1 adipocytes. Differentiated adipocytes were incubated with 50 or 100 μg POE or 100 nM insulin for 24 h, and then cells were lysed and subjected to immunoblot analysis.
Effect of POE on AMPK expression
Because AMPK phosphorylation stimulates glucose uptake by translocating GLUT4 to the PM, 12 the effect of POE on AMPK phosphorylation was investigated. As shown in Figure 4, POE significantly stimulated AMPK phosphorylation in 3T3-L1 adipocytes. Phosphorylated AMPK was significantly upregulated to 118% and 154% of the control value upon treatment with 50 and 100 μg/mL POE, respectively. This finding showed that POE may also increase glucose uptake by stimulating AMPK phosphorylation.
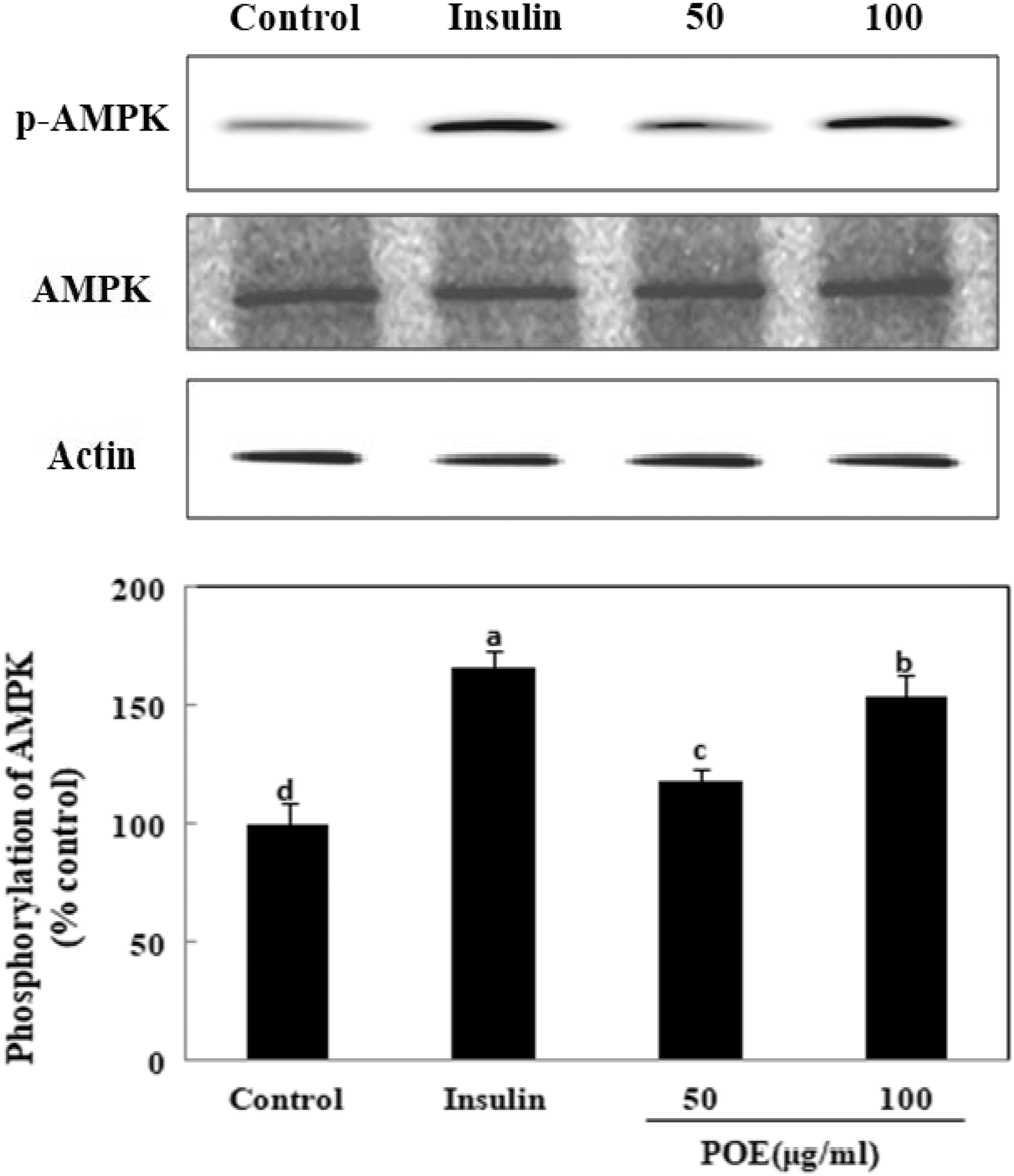
Effects of POE on AMPK expression in 3T3-L1 adipocytes. Differentiated adipocytes were incubated with 50 or 100 μg POE or 100 nM insulin for 24 h, and then cells were extracted and subjected to immunoblot analysis. Each value is expressed as mean ± SD of triplicate experiments. a–dValues with different superscript letters were significantly different at P < .05 as determined by Duncan's multiple range test. AMPK, 5′-AMP-activated kinase.
Effect of POE on PM-GLUT4 expression
PM-GLUT4 expression was examined to confirm the importance of GLUT4 in POE-stimulated glucose uptake. PM-GLUT4 expression was significantly increased upon POE treatment in 3T3-L1 adipocytes. At 100 μg/mL, POE significantly increased PM-GLUT4 expression to 285% of the value observed in untreated control cells (Fig. 5). However, PM-GLUT4 expression was significantly decreased to 130% of the value detected upon POE treatment by pretreatment with WM, a PI3K inhibitor. In addition, PM-GLUT4 expression was significantly decreased to 164% of the value detected upon treatment of POE by pretreatment with CC, an AMPK inhibitor. This finding showed that POE increased PM-GLUT4 expression and glucose uptake through activating PI3K/Akt and AMPK pathways.
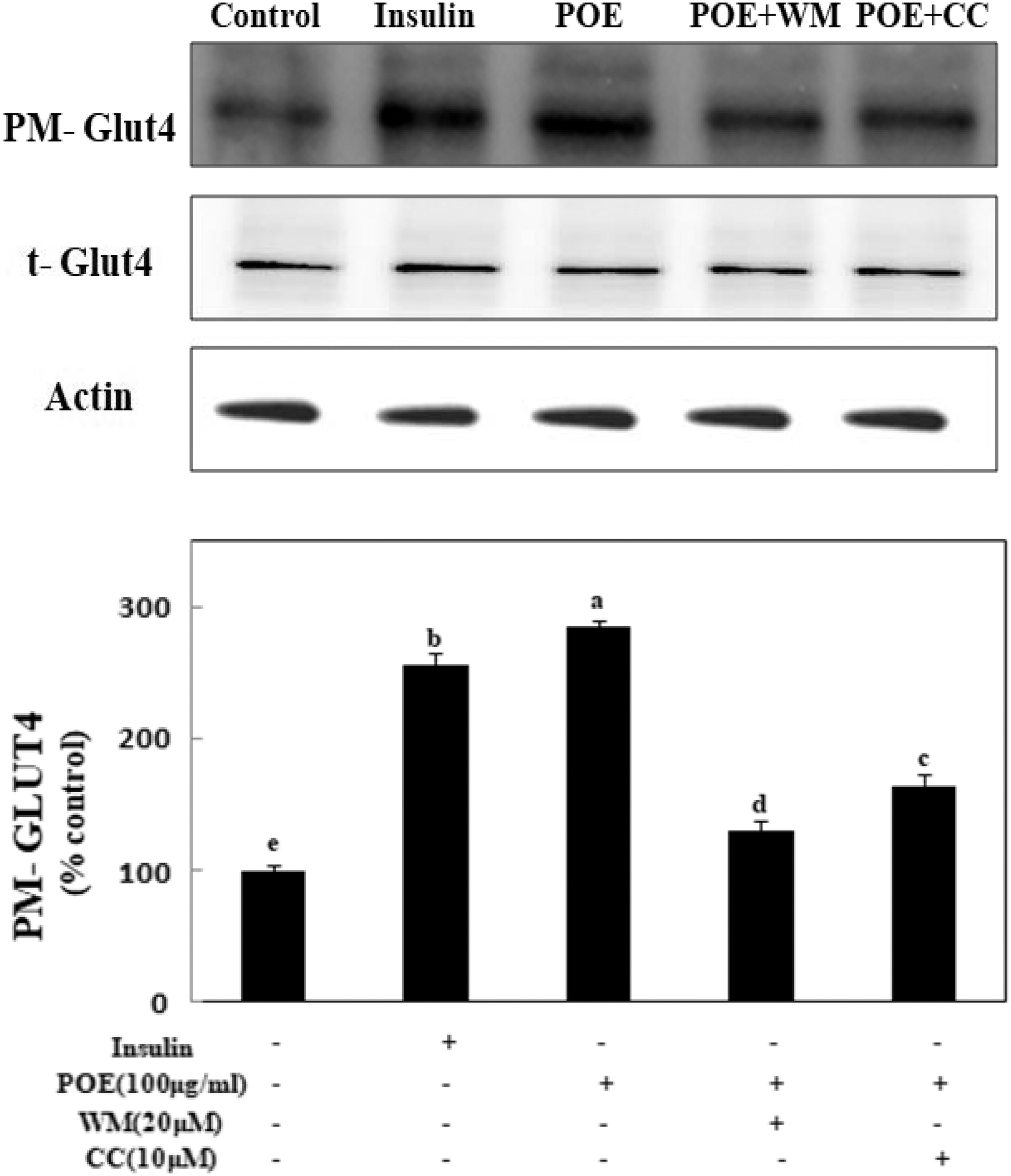
Effects of POE on GLUT4 protein expression in 3T3-L1 adipocytes. Differentiated adipocytes were incubated with 50 or 100 μg POE or 100 nM insulin for 24 h or in combination with 10 μM CC, 20 μM WM, and then cells were extracted and subjected to immunoblot analysis. Each value is expressed as mean ± SD of triplicate experiments. a–eValues with different superscript letters were significantly different at P < .05 as determined by Duncan's multiple range test. CC, compound C; WM, Wortmannin.
Effect of POE combined with CC and WM on glucose uptake
To investigate the effect of POE combined with CC and WM on glucose uptake, we performed a 2-deoxyglucose uptake assay in 3T3-L1 adipocytes. Uptake of 2-deoxyglucose was markedly increased by treatment with POE in 3T3-L1 adipocytes (Fig. 6). When the cells were treated with 100 μg/mL POE, glucose uptake was increased to 1.70-fold compared with that of untreated control cells. However, when the cells were treated with 100 μg/mL POE combined with 10 μM CC or 20 μM WM, glucose uptake was significantly decreased. This finding confirmed that POE increased glucose uptake through activating PI3K and AMPK pathway (Fig. 7).
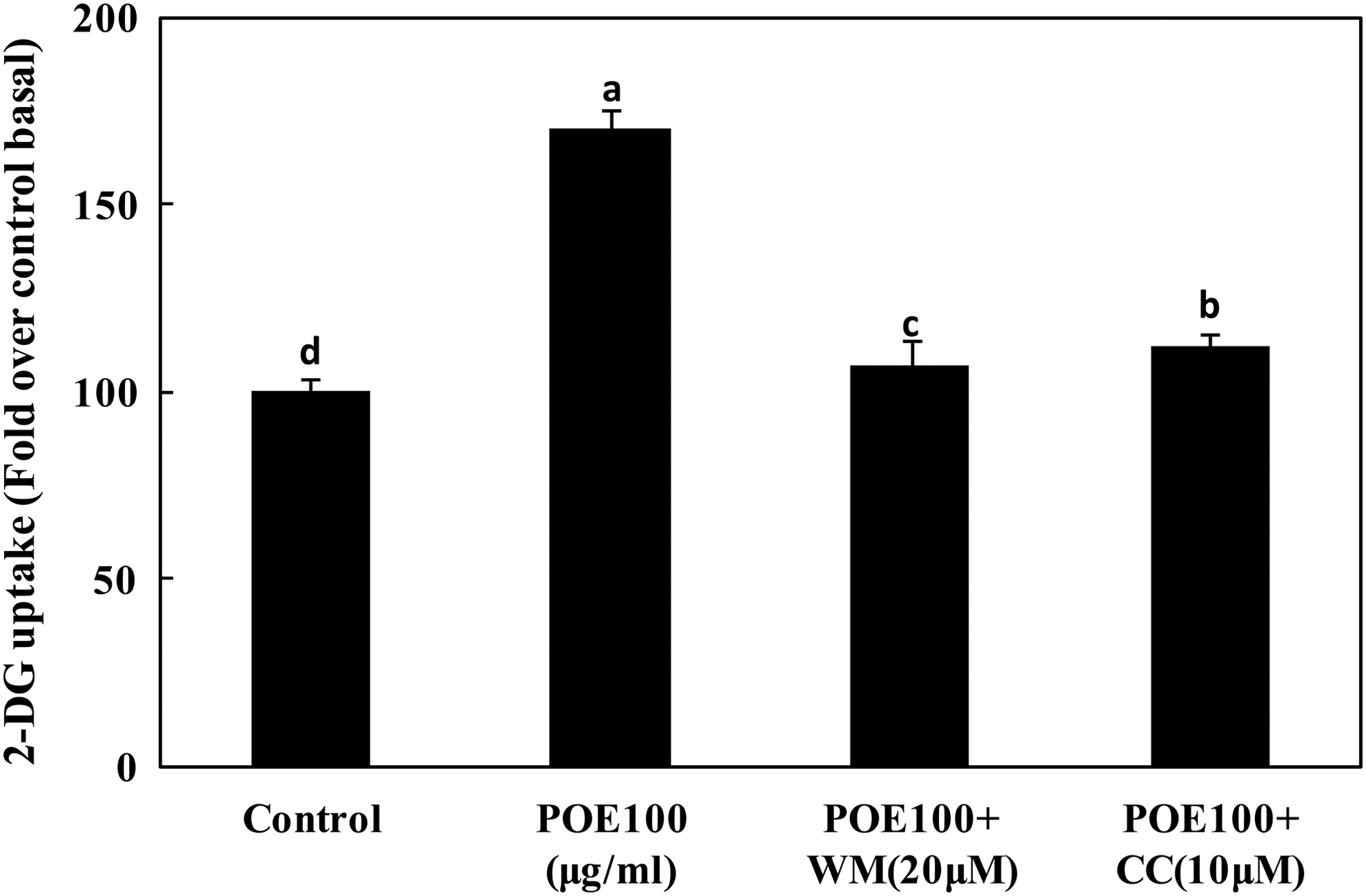
Effects of POE combined with CC and Wortmanin on glucose uptake in 3T3-L1 adipocytes. Differentiated adipocytes were incubated with 100 μg POE, 100 μg POE +10 μM CC, or 20 μM WM, and then 2-DG uptake was measured. Each value is expressed as mean ± SD of triplicate experiments. a–dValues with different superscript letters were significantly different at P < .05 as determined by Duncan's multiple range test.
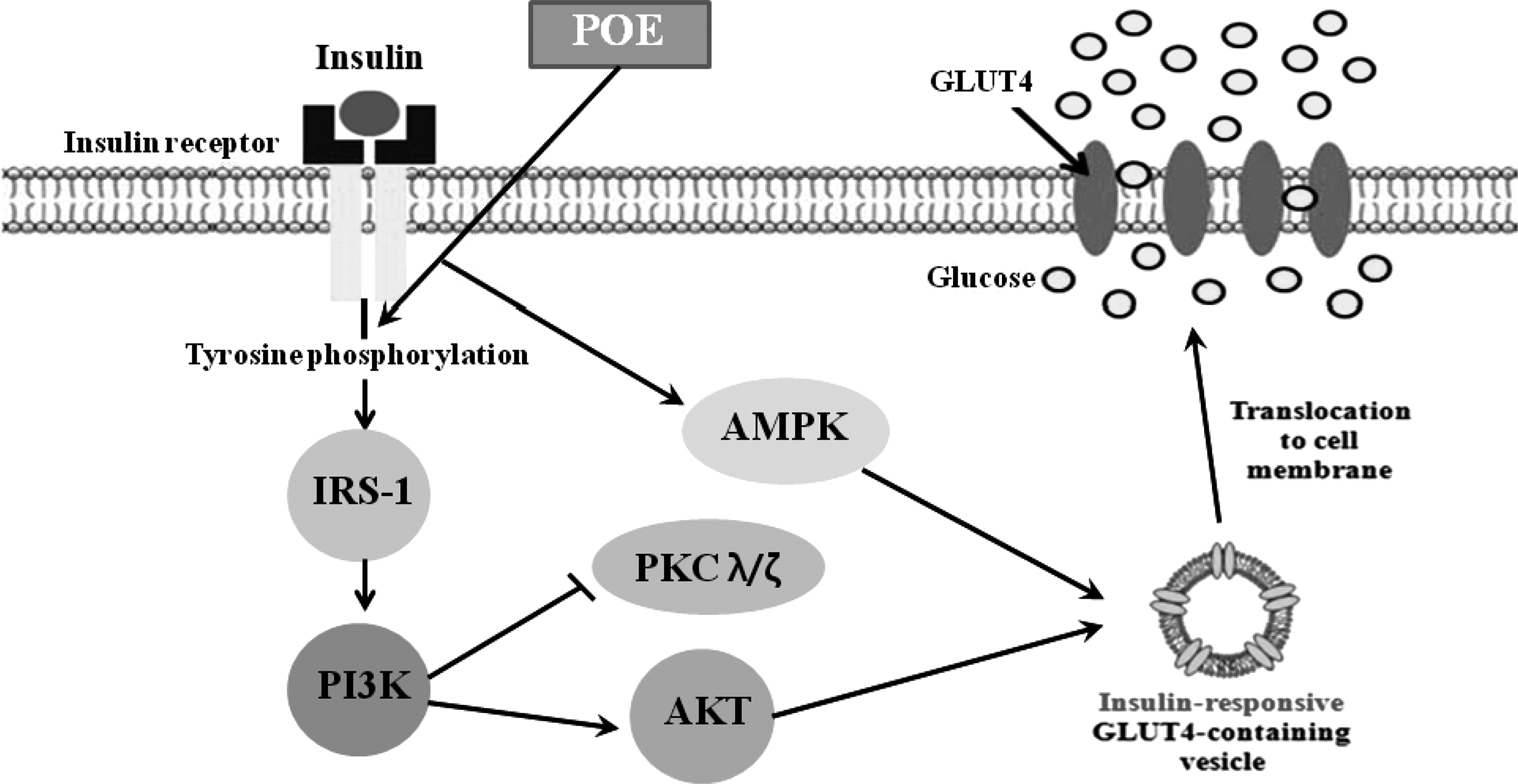
A proposed mechanism on glucose uptake of POE through PI3K/Akt pathway. POE enhanced glucose uptake, which was caused by increased GLUT4 expression at the plasma membrane through activating the PI3K/Akt pathway.
Discussion
P. oleracea is an annual plant widely distributed in Korea and China. This plant has long been used as a traditional remedy and its extract is known to show various biological activities, 5 –10,13,14 but its effect on glucose uptake was not previously investigated. Thus, this study was designed to measure the effect of POE on glucose uptake and investigate the cellular mechanisms underlying this effect in 3T3-L1 adipocytes. Hyperglycemia is a main symptom of type 2 diabetes, a chronic progressive metabolic disease. Hyperglycemia can exacerbate the damaged glucose uptake by interfering with the insulin signaling pathway in insulin target cells. 15 The control of glucose uptake is necessary for treating diabetes. Glucose uptake mainly occurs in insulin-sensitive tissues, such as skeletal muscle and adipose tissue. Insulin promotes glucose uptake by translocating GLUT4 to the PM through the insulin signaling pathway in 3T3-L1 adipocytes. 16 Stimulating glucose uptake might occur through an insulin signaling pathway-dependent mechanism in cells.
POE significantly stimulated glucose uptake in 3T3-L1 adipocytes. Stimulating glucose uptake into the cells is important for treating type 2 diabetes. Type 2 diabetes results from defective insulin sensitivity or impaired insulin secretion. Insulin maintains glucose homeostasis largely by stimulating glucose uptake into muscle and adipose tissues, which is a process mediated by GLUT4. GLUT4 plays an important role in glucose uptake in adipose tissue, which is essential for maintaining glucose homeostasis. Glucose uptake by GLUT4 can be activated by at least two major mechanisms, such as the PI3K and AMPK pathways. 16,17 In the PI3K pathway, insulin stimulates glucose uptake through insulin receptor-mediated phosphorylation of IRS-1 and intermediates that activate PI3K. PI3K activation is a major factor in the early insulin signaling pathway. This insulin signaling-mediated PI3K activation divides into two pathways: an Akt-mediated pathway and an atypical PKC-dependent pathway. 18 The activation of Akt or atypical PKCλ/ζ, downstream targets of IRS/PI3K, translocates GLUT4 to the PM and finally transfers glucose into the cells. 19
To elucidate the mechanism of POE action on glucose uptake, we investigated the activation of IRS/PI3K/Akt or atypical PKCλ/ζ in POE-treated 3T3-L1 adipocytes. The activations of IRS-1 and PI3K were significantly increased by POE treatment in the cells. Akt phosphorylation was also significantly increased, but the phosphorylation of atypical PKCλ/ζ was not observed. The phosphorylation of Akt by PI3K activation induces the translocation of GLUT4 from intracellular vesicles to the PM. 19 We observed that POE treatment also enhanced the translocation of GLUT4 to the PM. The expression of PM-GLUT4 may have been enhanced through activation of the IRS-1/PI3K/Akt pathway. These results suggested that POE might enhance glucose uptake through the PI3K/Akt pathway, but not the PKCλ/ζ-dependent pathway, in 3T3-L1 adipocytes.
P. oleracea is a well-known edible plant that is widely used as a medicinal herb. This plant contains various kinds of compounds such as triterpenoids, alkaloids, coumarins, and flavonoids. 20,21 The main active compounds of this plant are flavonoids. 22 Total phenol contents of P. oleracea ethanol extract were studied as 58.16 mg/g. Among them, the flavonoid content was studied as 20.08 mg/g. The content of flavonoid compounds in POE is reported to be higher than that of other kinds of compounds, and flavonoids were classified as the main compounds of POE. 23 Recent studies reported that flavonoids are involved in stimulating glucose uptake through insulin signaling pathways. 24,25 Flavonoids induce IRS-1 phosphorylation, which is an important protein in the insulin signaling pathway. Furthermore, these events could lead to the upregulation of PI3K/Akt signaling and stimulate glucose uptake. 26 Pentacyclic triterpenoids were also reported to have insulin-mimetic activity as insulin receptor activators of the PI3K signaling pathway. 27 The flavonoids and triterpenoids are active compounds of POE in the preliminary result from our study (data not shown). Thus, we assume that the flavonoids and triterpenoids contained in P. orelacea extract might stimulate glucose uptake through the IRS-1/PI3K/Akt pathway in 3T3-L1 adipocytes.
In addition to the PI3K pathway, the AMPK pathway is another intracellular signaling pathway that plays a major role in controlling glucose uptake in adipocytes. AMPK activation stimulates glucose uptake and GLUT4 translocation to the PM independently of insulin. 28 AMPK has been considered as a potential therapeutic target for treating type 2 diabetes due to its role as a central regulator of glucose metabolism. Activation of the AMPK pathway increases the glucose uptake by insulin-sensitive tissues to ameliorate insulin resistance. 29 To identify the effect of POE on glucose uptake by AMPK activation, we investigated the levels of phosphorylated AMPK. Treatment with POE significantly increased pAMPK levels, indicating AMPK activation, in 3T3-L1 adipocytes. Agents that increase pAMPK expression had an antihyperglycemic effect by facilitating GLUT4 translocation and glucose uptake. Metformin, one such agent with a beneficial effect on type 2 diabetes, increases glucose uptake through AMPK activation. 30 Studies reported that pachymic acid, a triterpenoid, increases AMPK activation and glucose uptake in 3T3-L1 adipocytes, 31 and polyphenols in black tea also activate AMPK. 32 These reports suggested that triterpenoid and polyphenol compounds play important roles in glucose uptake through AMPK and PI3K/Akt pathways. Thus, the effect of POE on glucose uptake seems to be partly due to the triterpenoids and polyphenols present in the extract.
In conclusion, POE increased glucose uptake through activating the PI3K/Akt and AMPK pathways in 3T3-L1 adipocytes. Activation of the PI3K/Akt and AMPK pathways by POE was confirmed using the PI3K inhibitor, WM, and the AMPK inhibitor, CC, which abolished the POE-mediated PM-GLUT4 expression. However, further studies are needed to elucidate which compound(s) in POE may be responsible for these effects.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT, and Future Planning (2017R1A2B4005323).
Author Disclosure Statement
No competing financial interests exist.