
Abstract
Photoaging is generally the result of chronic exposure to the sun and ultraviolet (UV) radiation, which causes skin damage. In this study, we developed a UVB-induced hairless mouse model to determine whether Lactobacillus acidophilus IDCC 3302 tyndallizate (ACT3302) can enhance photodamaged skin repair. Mice (6 weeks old) were divided into six groups containing normal, UVB-treated vehicle, and UVB-treated ACT3302 (1 × 105, 1 × 106, 1 × 107, and 1 × 108 cells). Epidermal thickness was increased by UVB, but the thickening was lessened by ACT3302 as was the transepidermal water loss (TEWL). However, ACT3302 increased capacitance and decreased TEWL. Skin tissue staining to evaluate skin collagen increases in the number of skin collagen bundles in UVB-treated ACT3302 mice. UVB irradiation increased matrix metalloproteinase (MMP) and proinflammatory cytokine expression and activated mitogen-activated protein kinases in hairless mice; these changes were also attenuated by ACT3302. We conclude that ACT3302 effectively suppressed wrinkle formation induced by UVB irradiation through MMP downregulation. Therefore, ACT3302 potentially prevents skin photoaging and wrinkle formation.
Introduction
A
Skin aging can be categorized into two types: intrinsic (chronological) and extrinsic (photoaging). 9 Extrinsic aging is caused by environmental oxidative factors, such as exposure to ultraviolet (UV) radiation, damaging collagen fibers, and excessive accumulation of abnormal elastic fibers. 10 UV irradiation induces changes in the physiological and biochemical characteristics of the skin by increasing epidermal thickness, skin damage, skin dehydration, wrinkle formation, and transepidermal water loss (TEWL). 11 Hyaluronan (HA) plays an important role in hydration, and the most dramatic histochemical change observed in aged skin in the extracellular matrix (ECM) is the marked disappearance of epidermal HA. 12
UV-irradiated and photodamaged skin has a reduced capacity for collagen synthesis and increased expression levels and activity of matrix metalloproteinase (MMPs), particularly MMP-1, MMP-3, and MMP-9. 13 MMP degrades ECM components, which are important general biological processes. 14 The expression of these MMPs was assessed by UV irradiation and it is generally known that skin wrinkles are alleviated when inhibiting their expression, and thus they are used as markers for wrinkling and skin inflammation. 14 In addition, increased levels of reactive oxygen species (ROS) induce collagen degradation through the upregulation of MMPs. 15
Exposure to UVB radiation significantly increases ROS concentrations, modulates cytokine expression, and promotes the expression of collagen-degrading MMPs. 16 Enzymatic antioxidants, such as superoxide dismutase (SOD) and catalase (CAT), are important endogenous antioxidants, which play an important role in human skin. 17 The inflammatory responses induced by UVB are manifested by an increase in skin inflammatory cell infiltration and increased prostaglandin synthesis. 18 The primary target of UV irradiation is keratinocytes, which initiate many responses to UV exposure by releasing proinflammatory cytokines. 19
In recent studies, many articles have reported that probiotic bacteria might be equally effective for protecting the skin from photoaging by affecting skin homeostasis by regulating systemic immune status, including protecting skin from photoaging as alternative treatments, by controlling the systemic immune status, including the release of regulatory cytokines. 20,21 In related studies, treating mice with probiotic bacteria before UVB irradiation inhibited various skin damages. 22,23 Other studies have showed that probiotic bacteria attenuated skin inflammation and allergy. 24,25 Also, probiotic bacteria containing Lactobacillus acidophilus tyndallizate have been shown to improve skin properties, such as skin hydration and body characteristics, in clinical research. 26,27
Based on these findings, it has been demonstrated that probiotic bacteria are effective in immunological and physiological regulators of skin conditions. This study demonstrates that orally administered L. acidophilus IDCC 3302 tyndallizate (ACT3302) protected against photoaging by decreasing MMP levels through the MAPK signaling pathway.
Materials and Methods
Microorganism
Lactobacillus acidophilus IDCC 3302 (GenBank accession number KP325412.1), the bacterial strain used in this study, was originally isolated from the feces of a Korean breast-fed infant after obtaining oral consent from the parents.
Tyndallization process
Bacterial cells were anaerobically cultured in yeast extract-based medium at 37°C overnight and then centrifuged at 12,500 g for 1 h using a continuous filter (Alfa Laval, Lund, Sweden). Thereafter, the pellets and the supernatant were concentrated five times using a vacuum concentrator (Dy-machine, Seoul, Korea) under reduced pressure at 80°C and then mixed with dry-sterilized cornstarch powder. The mix was then frozen and lyophilized to obtain ACT3302 powder.
Enumeration of total bacterial cell numbers and scanning electron microscopy
The total bacterial cell count of the ACT3302 suspension was determined using a Neubauer counting chamber (Marienfeld-Superior, Lauda-Königshofen, Germany) with 2.5 × 10−4 mm3 squares. The sample was diluted 103-fold with PBS, 25 small squares of cell particles were counted at a magnification of 400 × . Cell counts were calculated as follows: number of bacterial cells per mL = average number of cells counted per square × 4000 squares per mm3 × 103 mm3 per mL × 103 (dilution).
The cells were observed using high-resolution field emission scanning electron microscopy (FE-SEM, Merlin, Carl Zeiss AG, Germany) with and without tyndallization. The cell surface structures were viewed at an accelerating voltage of 10 kV for high-magnification imaging (25,000 × ).
Experimental animals and treatments
Male hairless mice (Hos/HR-1, 6 weeks old) were purchased from Japan SLC, Inc. (Sizuoka, Japan). The mice were housed in a climate control room maintained at 24°C and 50% humidity, and a 12-h light/12-h dark cycle. All mice were acclimatized for a week before the study and had free access to food and water. Mice were divided into six groups containing normal, UVB-treated vehicle, and UVB-treated ACT3302 (dosages: 1 × 105, 1 × 106, 1 × 107, and 1 × 108 cells) (n = 6 per group). The UVB-treated ACT3302 groups were orally administered 0.1 mL of ACT3302 for 5 days a week for 12 weeks. The vehicle group was orally administered drinking water, and the unexposed normal group animals did not receive any treatment. The Korea Institute of Oriental Medicine Institutional Animal Care and Use Committee (16-059) approved all experimental protocols.
UVB irradiation in mice
ACT3302 was orally administered after UVB irradiation on the dorsal skin was investigated. UVB irradiation was conducted for 12 weeks at 48-h intervals using a UVM-225D Mineralight UV Display Lamp (UVP, Phoenix, AZ, USA). The dose gradually increased from 60 mJ/cm2 per exposure at week one to 90 mJ/cm2 at week seven (1 minimal erythematous dose = 60 mJ/cm2).
Evaluation of TEWL and capacitance, and histological examination
TEWL was measured using Tewameter™ 300 (Courage and Khazaka Electronics, Cologne, Germany) and skin capacitance was measured using a corneometer (Courage and Khazaka Electronics, Cologne, Germany). The dorsal skin was fixed in 10% neutral buffered formalin, embedded in paraffin, and incised at 5-μm intervals to stain skin tissue. Hematoxylin and Eosin (H&E) and Masson's Trichrome were used for analysis. Epidermal thickness was measured by light microscopy using an eyepiece micrometer (Olympus, Japan).
ELISA assay for MMP, hyaluronic acid, and antioxidant enzyme activities
MMP-1, MMP-9, and HA were assayed in skin tissues using commercial ELISA Kits (R&D Systems, Minneapolis, MN, USA). The activities of SOD and CAT were also determined in skin tissue using commercial kits (Cayman Chemical Co., Ann Arbor, MI, USA). After all steps were completed, the levels of MMP-1, MMP-9, and HA were quantified by colorimetric analysis using plate reader (Molecular Devices). Also, SOD and CAT activities were determined using a plate reader.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from UVB-irradiated mouse skin tissue, using protocols recommended by the TRIzol reagent manufacturer (Invitrogen, Carlsbad, CA, USA). The final RNA pellet was dried and diluted with diethylpyrocarbonate (DEPC) to quantitate the RNA. A cDNA Reverse Transcription Kit was used for cDNA synthesis and quantitative real-time polymerase chain reaction (qRT-PCR) using TaqMan assays (Applied Biosystems, Foster City, CA, USA) specific for IL-1β, and IL-6 on a QuantStudio™ 6 Flex Real-Time PCR system (Applied Biosystems). Each sample was analyzed in triplicate and relative mRNA expression levels were calculated using the ΔΔCt method and normalized to the β-actin mRNA level in each sample.
Western blotting
Skin tissue was extracted using Precellys®24 (Bertin, USA) tissue homogenizer and were centrifuged at 18,000 g for 10 min. The protein content in the supernatant was determined using a Bio-Rad Protein Assay Kit (Bio-Rad). After extracting proteins from skin tissue, the protein concentration was measured and sample buffer was added to prepare a sample containing the same concentration of protein. A total of 20 μg of proteins were subjected to 10% SDS-PAGE and electrophoresis at 100 V, the gel containing proteins was absorbed onto nitrocellulose membranes. The membranes were then blocked by incubation with TBS-T (0.1% Tween 20) containing 5% BSA. Then, the primary antibody (anti-MMP-1, anti-MMP-9, anti-phospho-ERK, anti-phospho-MEK, anti-phospho-p38, anti-phospho-JNK, anti-MEK, anti-ERK, anti-p38, or anti-JNK) and then washed with TBS-T. Protein bands were visualized after incubation with conjugated secondary antibodies (goat anti-rabbit IgG or goat anti-mouse IgG Ab diluted 1:5000). Proteins were detected using enhanced chemiluminescence (ECL solution).
Statistical analysis
All data are expressed as mean±standard error of the mean (SEM). Results were analyzed by variance analysis (ANOVA) using Tukey's multiple comparison test to analyze differences, and P < .05 was considered to indicate statistically significant differences.
Results
SEM analysis of ACT3302
Figure 1 shows the effect of the tyndallization process. Cells of L. acidophilus IDCC 3302 tyndallizate contracted or fragmented in contrast to untreated L. acidophilus IDCC 3302.

FE-SEM images of nontreated (panels on the left-hand side) and tyndallized (panels on the right-hand side) Lactobacillus acidophilus IDCC 3302.
Effect of ACT3302 on the epidermis thickness and histological observations in UVB-irradiated hairless mice
Histological analysis of H&E-stained skin sections was used to confirm the antiphotoaging effect of ACT3302, and changes in collagen fibers were confirmed by Masson's Trichrome staining (Fig. 2A, B). Compared with the normal group, the UVB-treated group was found to have decreased amounts and densities of collagen fibers that were increased by administration of ACT3302. The skin epidermal thickness of the UVB-treated ACT3302 group, at a treatment concentration of 1 × 108, was 27.98 μm and was lower in a dose-dependent manner than the 51.65-μm thickness observed in the UVB-treated vehicle group (Fig. 2C). Thus, these results suggest that ACT3302 is an effective photoprotective agent that decreased the thickness of the epidermis and improved collagen integrity.
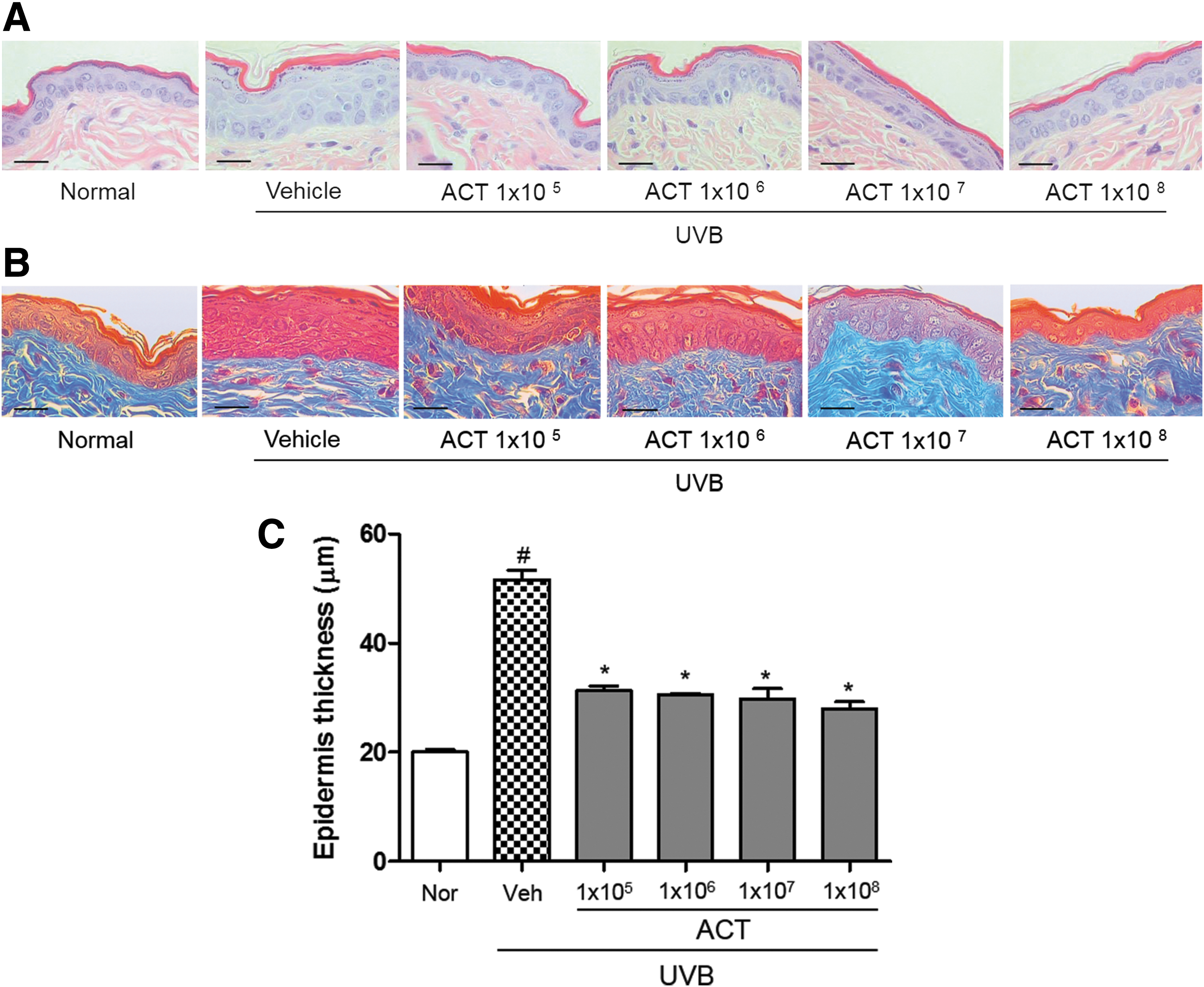
Effect of ACT3302 on UVB irradiation-induced epidermal thickening in hairless mouse dorsal skin.
Effect of ACT3302 on skin hydration
In this study, TEWL of the normal group was 1.58 g/m 2 /h, and in the UVB-treated vehicle group it was 3.92 g/m2/h. However, TEWL was attenuated in the UVB-treated group administered ACT3302 (Fig. 3A). In addition, it was confirmed that UVB reduced stratum corneum hydration, but treatment with ACT3302 significantly suppressed decreases in capacitance (Fig. 3B). The hyaluronic acid content in skin tissue was increased in the UVB-treated ACT3302 group compared with the UVB-treated vehicle administration group.
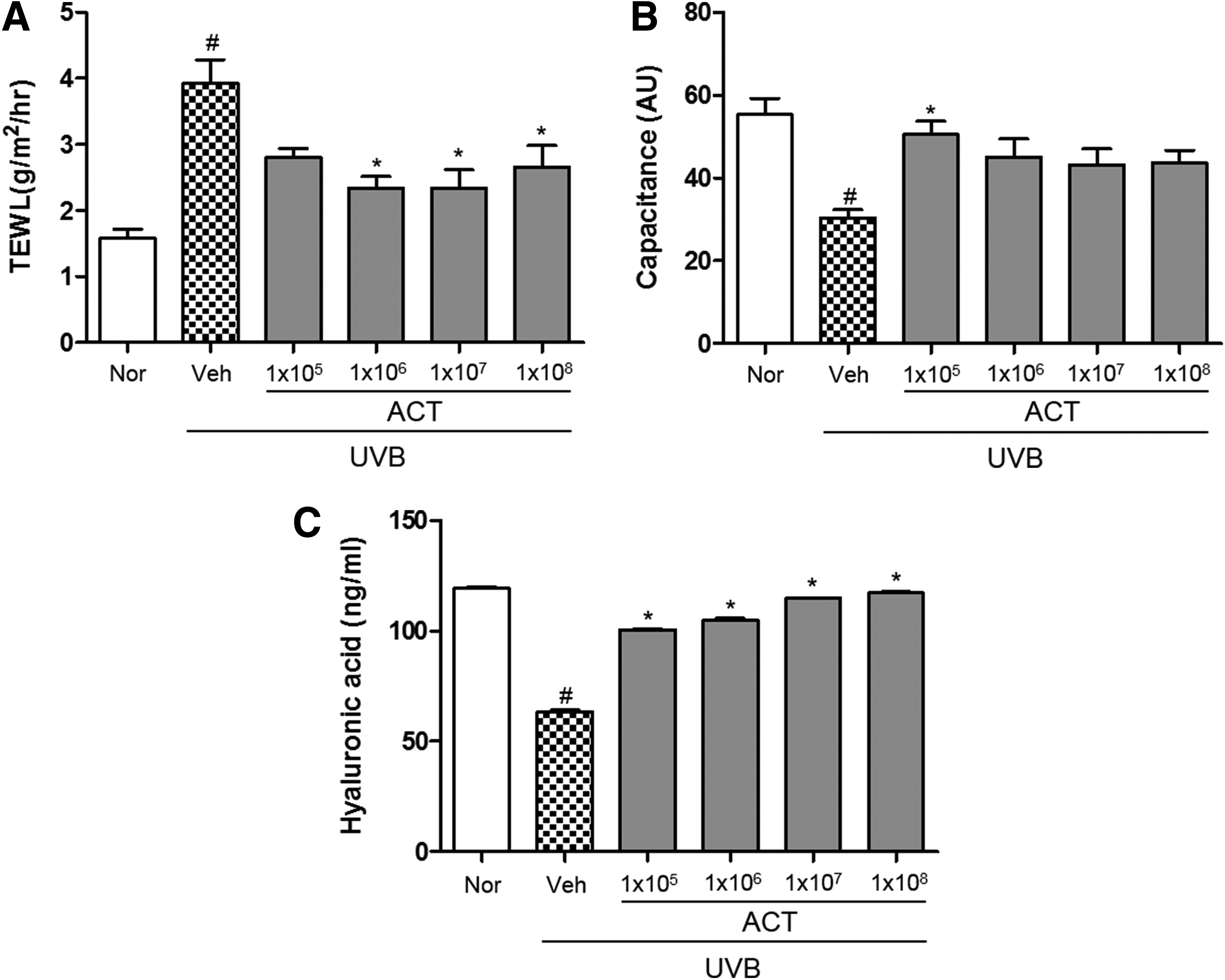
Effects of ACT3302 on UVB-induced skin hydration in hairless mice. Effects of ACT3302 on
ACT3302 inhibits UVB-induced MMP-1 and MMP-9 expression
ACT3302 decreased the expression of MMP-1 and MMP-9 in a dose-dependent manner compared with the UVB irradiation group (Fig. 4A, B). As shown in Figure 4C, the expression of protein levels of MMPs, including MMP-1 and MMP-9, was attenuated by ACT3302.
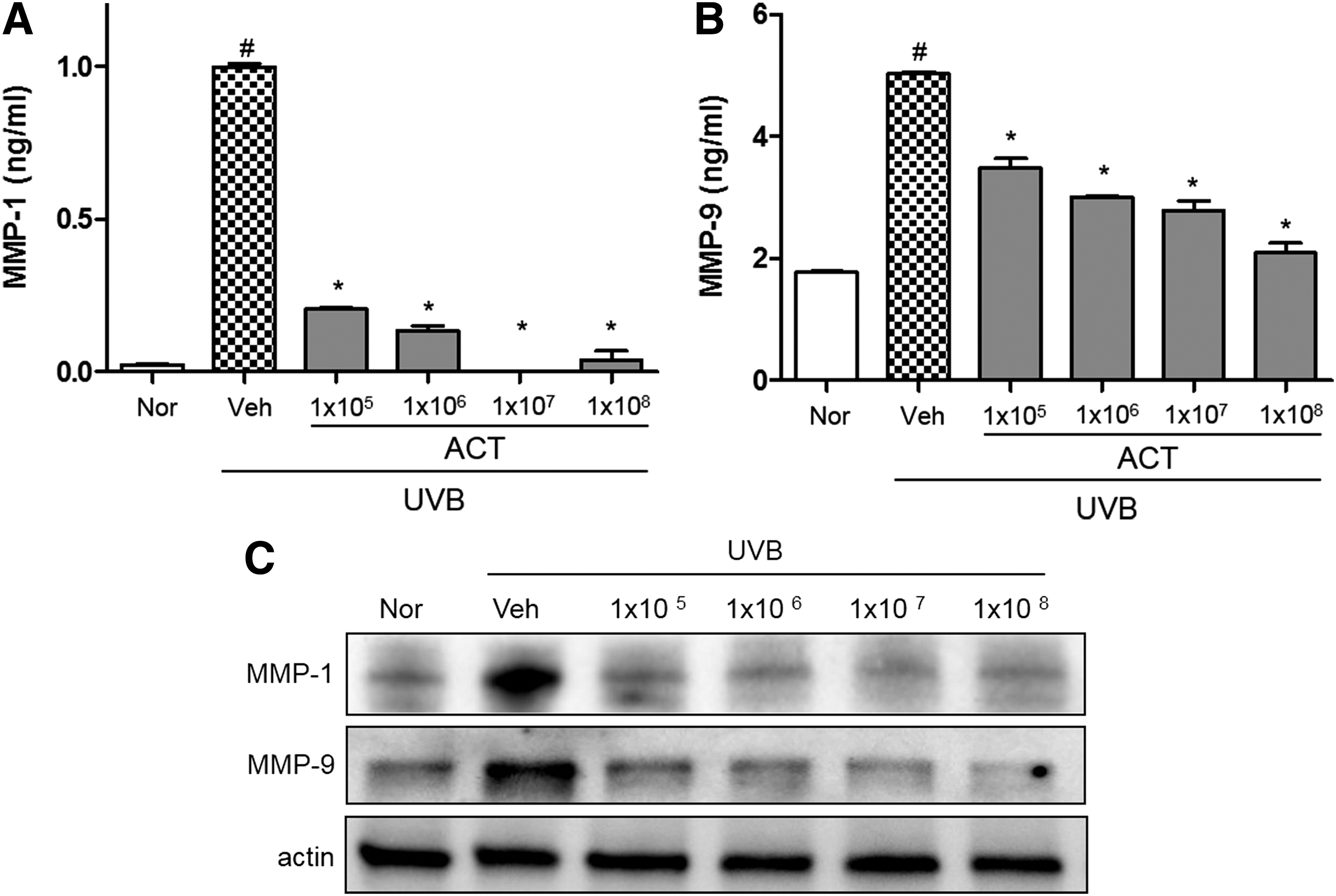
Inhibitory activity of ACT3302 on UVB-induced MMP-1 and MMP-9 protein expression in a photodamaged hairless mouse model in skin tissues.
Effects of ACT3302 on activity of antioxidant enzymes in UVB-induced hairless mice
In this study, radical-scavenging activity by antioxidant enzymes was observed in hairless mice after UVB irradiation. SOD activity was decreased in UVB-irradiated hairless mice and increased dose dependently by ACT3302 administration (Fig. 5A). In addition, CAT activity was increased by administration of ACT3302 (Fig. 5B). The antioxidant enzyme activity of ACT3302 was shown to further decrease UVB-induced oxidative stress by eliminating free radicals.
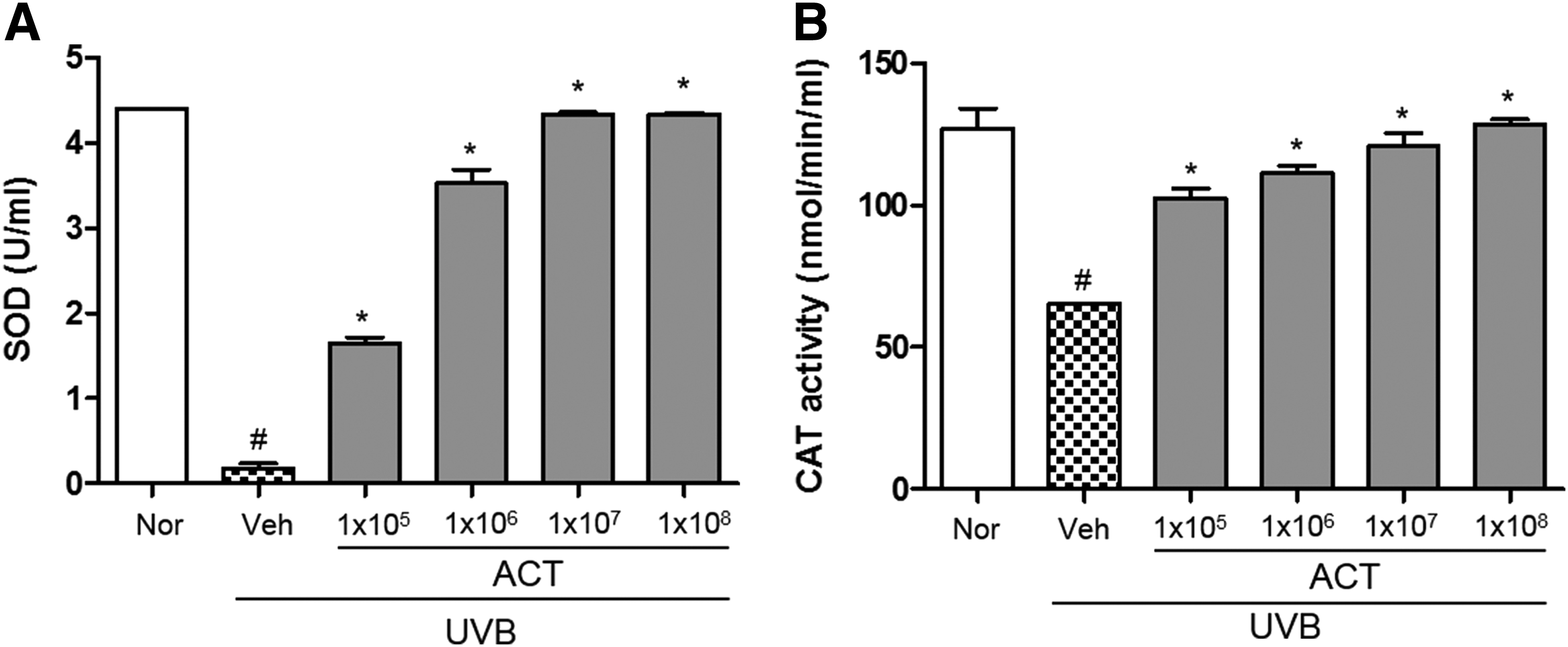
Effects of ACT3302 on the activity of
Effects of ACT3302 on the mRNA expression of inflammatory cytokines
Expression of proinflammatory cytokine mRNA was examined to evaluate the modulatory effect of ACT3302 on the production of proinflammatory cytokines in UVB-irradiated skin of hairless mice. As shown in Figure 6, levels of IL-1β and IL-6 mRNA were increased after exposure to UVB radiation, and were inhibited dose dependently by treatment with ACT3302. This result suggests that ACT3302 exhibits a photoprotective effect by reducing the inflammatory response to UVB irradiation.
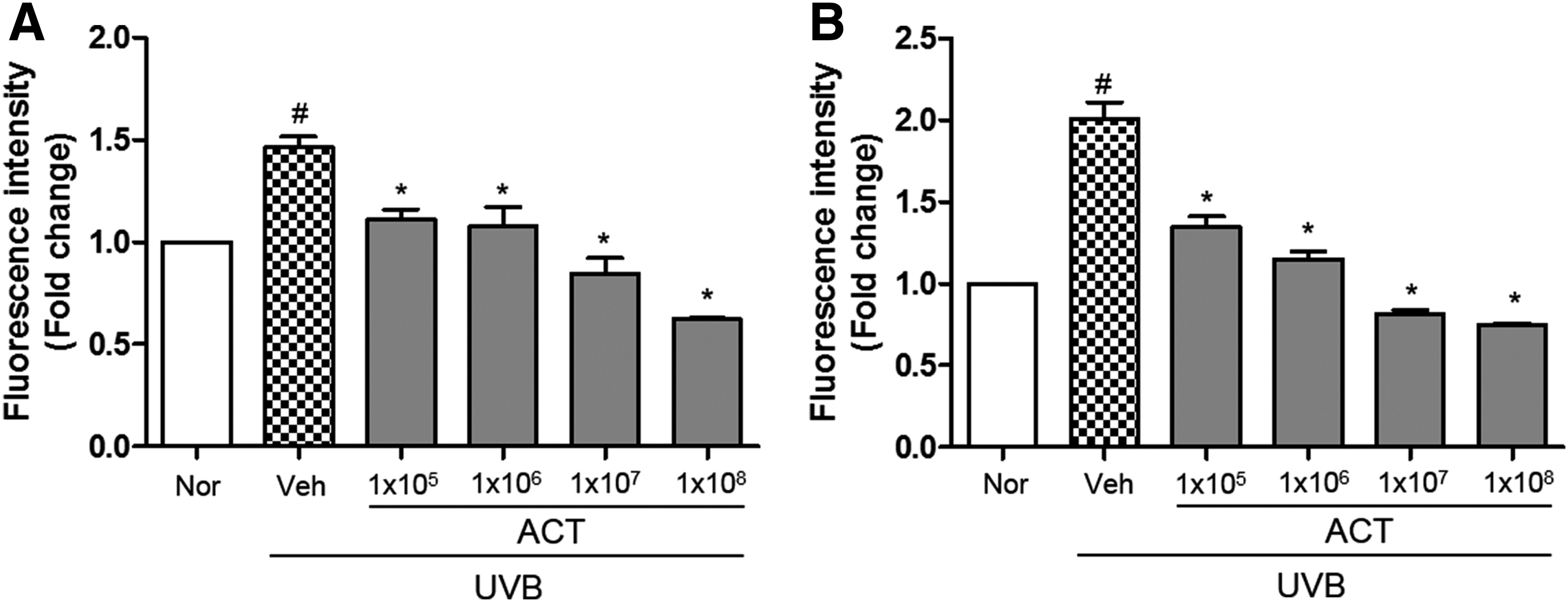
Effects of ACT3302 on proinflammatory cytokines in UVB-irradiated hairless mouse skin. The mRNA expression levels of
Effect of ACT3302 on MAPK phosphorylation in UVB-exposed hairless mice
We investigated the effect of UVB irradiation on the activation of MEK, ERK, p38, and JNK. As shown in Figure 7, UVB irradiation induced the phosphorylation of p38 and JNK, whereas ACT3302 inhibited this effect. In addition, ACT3302 suppressed the UVB-induced phosphorylation of MAPK kinase (Fig. 7). These data indicate that ACT3302 protected mouse skin from UVB.
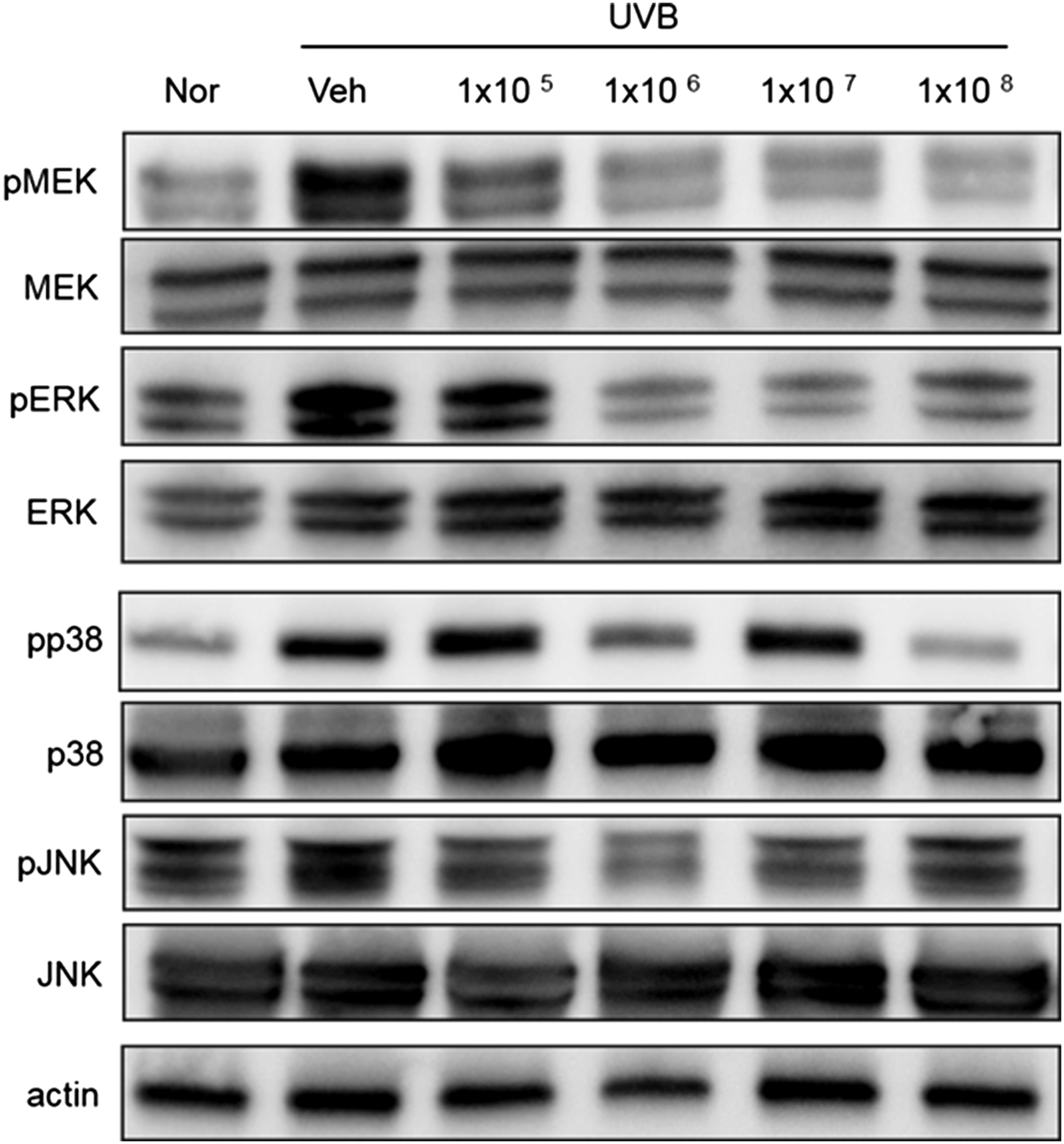
Effects of ACT3302 on the phosphorylation of MAPK in a UVB-irradiated mouse model. ACT3302 inhibited the phosphorylation of MEK, ERK, p38, and JNK.
Discussion
UV irradiation is a major reason for skin photoaging, which is characterized by skin wrinkles, as well as related skin diseases. 28 Epidermal thickening induced by UV irradiation is used as a parameter for inflammation and skin photoaging. 29 In the cosmetics and skin-care industries, TEWL measurement is used because it is directly related to the moisture barrier function of the skin. 30 As TEWL means water evaporation from skin in real time, skin barrier damage (UVB) increases TEWL. 31 UVB irradiation causes skin damage, resulting in skin dehydration and an increase in TEWL, a decrease in water content and a decrease in HA content. 32 Skin that is aged by UV irradiation is visually dry because increased TEWL impairs enzymatic functions. 30 Administration of ACT3302 effectively reduced skin damage and TEWL and increased skin hydration. Additionally, UVB irradiation reduced stratum corneum hydration and hyaluronic acid, but ACT3302 treatment blocked decreases in capacitance and hyaluronic acid.
UV irradiation increases the levels of ROS and antioxidant enzymes in the skin that increased ROS production changes the structure and function of multiple genes and proteins, resulting in skin damage. 33 UVB irradiation and environmental toxins are known to be related with the generation of ROS. 34 We examined whether the ROS-related mechanism of skin was involved in UVB-induced skin wrinkle formation.
Excessive expression of MMP by UV irradiation is an important mechanism for wrinkle formation through collagen degradation. 35 UVB irradiation exposure increases MMP levels by degrading collagen and reducing collagen synthesis, which damages skin connective tissue. 13 MMP expression induced by UVB radiation exposure leads to degradation of the ECM and thus contributes to the formation of wrinkles in the skin. 15 Treatment with ACT3302 before UVB irradiation decreased MMPs, including MMP-1 and MMP-9 expressions.
Exposure of keratinocytes to UVB induces cytokine secretion, such as IL-1, IL-6, IL-8, and IL-10, which is associated with a variety of skin inflammatory responses. 36 The qRT-PCR analyses showed that ACT3302 attenuated the expression of IL-1β and IL-6 induced by UVB.
MMP-1 is regulated by MAPKs and it has been shown that inhibition of MMP-1 expression is mediated by the inhibition of the MEK/ERK signaling pathway. 37 Activation of MEK/ERK signaling induced MMP-9 expression and is required for responses in epidermal keratinocytes that may be an important molecular target for modulation of MMP expression by antiphotoaging. 38,39 This study showed that ACT3302 attenuated the expression of UVB-induced MMP-1 and MMP-9, which were regulated by the MEK/ERK pathway.
In conclusion, ACT3302 inhibited the MMP-1 and MMP-9-related signaling pathway activated by UVB irradiation. ACT3302 also prevented UVB-induced skin thickening, wrinkle formation, and collagen fiber loss. Further investigation is needed to elucidate the detailed mechanism of MMP-1 and MMP-9 inhibition. These results suggest that ACT3302 could be a good photoprotective agent for skin care.
Footnotes
Acknowledgments
This research was supported by a grant funded by the Korea Institute of Oriental Medicine (K17300) and INNOPOLIS Foundation funded by the Ministry of Science, ICT & Future Planning, Republic of Korea (DDI-00664).
Author Contribution
Dae-Jung Kang and Sungwook Chae were responsible for project design. A-Rang Im and Byeonghun Lee were responsible for collecting and analyzing data and writing the article for submission.
Author Disclosure Statement
The authors of this article have no disclosures to report.