
Abstract
Nonalcoholic fatty liver disease is a serious liver disorder associated with oxidative stress. Black radish (Raphanus sativus L. var. niger) extract (BRE) can lower the risk of this disease. The hepatoprotective effect of BRE containing 3-(E)-(methylthio)methylene-2-pyrrolidinethione was evaluated in human hepatocyte carcinoma (HepG2) cells and in rat livers with carbon tetrachloride (CCl4)-induced hepatic injury. BRE was administered at 125, 250, 500, and 1000 μg/mL to the oleic acid-induced HepG2 cells. Male Sprague-Dawley rats were randomly divided into seven groups: the control group, BRE group, CCl4 group, and BRE + CCl4 group. BRE was administered orally at 125, 250, 500, and 1000 mg/kg/day once daily for 7 consecutive days, followed by a single oral treatment of 1.5 mL/kg CCl4. Inhibition of lipid accumulation, serum markers of liver injury, histological evaluations, levels of oxidative stress related enzymatic and nonenzymatic antioxidants in HepG2 cells and liver tissue were investigated. The protein expression of main liver P450 isoenzymes such as cytochrome p450(CYP)2E1, the expression of nuclear factor erythroid 2-related factor-2(Nrf-2) and heme oxygenase-1(HO-1) were also studied. BRE has an inhibitory effect on lipid accumulation and caused acute hepatotoxicity manifested by increased levels of lipid peroxidation, serum alanine aminotransferase, and aspartate aminotransferase with corresponding histopathological changes and high levels of oxidative stress. BRE treatment significantly increased the level of CYP2E1, Nrf-2, and HO-1 in a dose-dependent manner. Besides, 3-(E)-(methylthio)methylene-2-pyrrolidinethione significantly increased radical-scavenging effects and the expression of Nrf-2 in oleic acid-treated HepG2 cells. These results suggest that BRE treatment reduces lipid accumulation in oleic acid-induced steatosis of HepG2 cells, and has a hepatoprotective effect against CCl4-induced liver injury in rats, possibly through Nrf-2/HO-1-mediated antioxidant effects.
Introduction
C
Radish (Raphanus sativus L.) is a cruciferous plant that is widely consumed as a vegetable and is used in traditional medicine. 7 Recently, crude radish juice extract was shown to have gastroprotective effects in ethanol-induced gastric injury rats 8,9 and preventative against cadmium-induced immunotoxic and biochemical evaluations in rats 10 through its antioxidant effects. Furthermore, 4-(methylthio)-3-butenyl isothiocyanate, contained in radish, is known to decrease the severity of high fat diet-induced nonalcoholic fatty liver disease in rats, 11 suggesting that isothiocyanates play a role in hepatoprotection. Radishes contain a variety of bioactive components, including polyphenols, anthocyanins, polysaccharides, and glucosinolates, particularly in the roots and sprouts. 12
Black radish (R. sativus L. var. niger), which is cultivated in Mexico, Europe, and Asia, is a colored radish with antioxidant effects. 13,14 The juice and aqueous extract of black radish have pharmacological properties, including antilithiasic and hypolipidemic effects. 15,16 Both glucosinolate and its isothiocyanate metabolites induce antioxidation and detoxification in human hepatocyte carcinoma (HepG2) cells. 17
However, little is known about the hepatoprotective effects of black radish on CCl4-induced liver injury, the rat model of hepatotoxicity. In this study, we were to evaluate the effects of black radish extract (BRE) on rats with CCl4-induced liver injury, and to understand its pharmacological mechanism.
Materials and Methods
Cultivation and extraction of black radishes
Black radish seeds were obtained from Asia Seed Co., Ltd. (Seoul, Korea) and the plants were cultivated in the Seongsan area (Jeju-do, Korea).
The black radish flesh was washed, dipped in hot water (70–80°C) for 10 min, and dried by hot blast for 96 h. The dried material (400 g) was pulverized using a mill, and extracted with 70% aqueous ethyl alcohol at room temperature (RT) under stirring for 24 h. The combined resulting solutions were filtered, and then it was concentrated using a rotary evaporator at 37°C to produce a powdered extract (116.0 g).
Isolation and high performance liquid chromatography analysis
The 70% ethanol extract of black radish root was fractionated using Diaion® HP-20 (Supelco, PA, USA), and eluted using the method of decreasing polarity to obtain fractions from 100% distilled water, 50% methanol, and 100% methanol. The 50% methanol fraction (1 g) was subjected to Sephadex LH-20 column chromatography (GE Healthcare, PA, USA) with distilled water-methanol (4:6). Compound 1 (22.5 mg) was eluted first, followed by compound 2 (20.6 mg), and finally compound 3 (25.3 mg). To identify the chemical structures, we analyzed the isolated compounds using nuclear magnetic resonance (NMR) (AVANCE III: FT-NMR system, Bruker, MA, USA) and mass spectroscopy (Quattro micro API Mass, Waters Co., MA, USA). The three compounds were subject to high performance liquid chromatography (HPLC) analysis.
HPLC was analyzed on an Alliance 2695 System (Waters Co.) equipped with a 2998-photodiode-array detector. The separation was done on an Atlantis® C18 column (4.6 × 250 mm, i.d., 5 μm) (Waters Co.). The mobile phases were (A) water and (B) methanol using gradient elution: 100% A to 10% B in A for 5 min; 10% B in A to 80% B in A for 45 min; 80% B in A to 100% B for 5 min; and 100% B for 5 min.
Cell culture
The male HepG2 cell line was purchased from the Korean Cell Line Bank (Jongno-gu, Seoul, Korea) and maintained in HyClone™ Minimal Essential Medium with Earle's (MEM/EBSS) (GE Healthcare) supplemented with 10% fetal bovine serum (FBS; Alta Biologicals, CO, USA) and 1% penicillin-streptomycin (Gibco, Paisley, United Kingdom) in a 37°C incubator at 5% CO2. For the experiments, the cells were seeded in a 12-well cell culture plate (5 × 105 cells/well) and incubated for 16 h, the cells were incubated in DMEM with 0.5 mM nonesterified fatty acids (0.6 mM oleic acid) for a further 24 h to induce hepatosteatosis. To evaluate the cytoprotectiveness of BRE, the cells were incubated with different concentrations of BRE (125, 250, 500, and 1000 μg/mL) for 24 h in the DMEM containing 10% delipidated FBS (Cocalic Biologicals, Reamstown, USA).
Cell viability assay
The effect of BRE on cell viability was determined by using the MTT assay. Cells were seed on 96-well plate (1 × 104 cells/well), incubated for 24 h, and treated with BRE (125, 250, 500, and 1000 μg/mL) for 24 h. The cells were incubated with MTT (2 mg/mL) solution for 2 h. Then, the concentration of dissolved formazan was measured at an absorbance of 540 nm using a microplate reader (BioTek, State of Vermont, USA).
DPPH radical scavenging activity
DPPH radical scavenging activity was determined as previously described. 18 Briefly, mixtures containing equal volumes of sample and 0.4 mM DPPH solution were incubated at RT for 30 min. The absorbance was then measured at 517 nm. Ascorbic acid was used as the positive control.
Oil red O staining
Oleic acid induced HepG2 cells were fixed with 4% paraformaldehyde in phosphate buffered saline and stained for 20 min with 0.6% Oil Red O solution at RT to stain lipid droplets. After staining, the cells were washed to remove excess stain and visualized on a microscope (Nikon Eclipse TE2000, Tokyo, Japan). Oil Red O staining was semi-quantified using ImageJ software (NIH, Bethesda, MD, USA). The percentage of total area that was Oil Red O positive was determined using the following formula: percentage of Oil Red O area (%) = (area of blue pixels/total area) × 100. The results are shown as mean ± standard error of the mean.
Animals
Male Sprague-Dawley rats (200–300 g) were used for all experiments (OrientBio, Kyunggido, Korea). Animals were housed at a controlled temperature (24°C ± 2°C), humidity (55% ± 10%) with 12-h light/dark cycles, and fed a standard diet and water. All experimental procedures were conducted in accordance with the guidelines for the Care and Use of Laboratory Animals at Jeju National University (Permit number: 2016-0040).
Experimental design
Rats were randomly divided into seven groups (n = 6/each group). Distilled water was administered orally to normal controls. The black radish control groups were given a solution of BRE orally at a dose of 1000 mg/kg. BRE and vehicle-treated rats with CCl4-induced injury were administered BRE orally (125, 250, 500, and 1000 mg/kg) for 7 consecutive days before CCl4 injection.
CCl4 -induced hepatotoxicity
To induce liver injury, a 1:4 (v/v) mixture of CCl4 and sterile olive oil was administered orally (1.5 mL/kg). CCl4 was injected 3 h after the final administrations of BRE and vehicle. The rats were fasted for 24 h after CCl4 administration and then sacrificed. 6,19 At the time of sacrifice, blood samples and liver tissues were collected for serum chemistry and tissue examination, respectively.
Morphological analysis and immunohistochemistry
For histopathological analysis, liver tissue was fixed in 4% paraformaldehyde solution and routinely processed for paraffin embedding. To see lipid droplets in liver tissues, Oil Red O staining, as done in oleic acid induced steatosis in HepG2 cells, was done on fixed frozen sections of liver tissues. Sections (5 μm) of paraffin-embedded liver tissue were deparaffinized and then stained with hematoxylin and eosin (H&E). Immunohistochemistry was performed using the ABC elite kit (Vector Labs, Burlingame, CA, USA) according to the manufacturer s recommendations. For immunohistochemistry, the primary anti-CYP2E1 (1:400 dilution; Abcam, MA, USA) antibody was incubated with the liver sections for 1 h. The visualized reaction was developed using a diaminobenzidine (DAB) substrate kit (Vector Labs).
Serological analysis
Blood samples were allowed to coagulate at RT followed by centrifugation for serum fraction collection. To determine the extent of liver damage, serum levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), triglyceride, and total cholesterol were measured using a FUJI DRI-CHEM 4200 (FUJIFILM, Tokyo, Japan).
Lipid peroxidation products and antioxidant enzyme activities in the liver
Lipid peroxidation was evaluated by measuring the level of thiobarbituric acid-reactive substances (TBARS) in the homogenate and expressed as the concentration of malondialdehyde (MDA). Briefly, the liver was homogenized in ice-cold 1.15% KCl, and 500 μL of the homogenate supernatant was mixed with 3 mL 1% phosphoric acid (Sigma-Aldrich, St. Louis, MO, USA) and 1 mL 0.6% aqueous solution of 4,6-dihydroxy-2-mercaptopyrimidine (Alfa Aesar, Ward Hill, MA, USA). The TBARS was measured at 540 nm. For measure of antioxidant enzyme activities including superoxide dismutase (SOD) and catalase (CAT), the liver tissues were homogenized in 50 mM phosphate buffer (pH 7.4), the levels of SOD and CAT were measured using commercially available kits (Abcam, San Francisco, CA, USA) according to the manufacturer's instructions. Absorbance values were measured using a Biotrak II Visible Plate Reader (Amersham Biosciences, Piscataway, NJ, USA) at 570 nm.
Western blot analysis
HepG2 cells and liver tissue were lysed in radioimmunoprecipitation buffer. For the immunoblot assay, 40 μg of protein underwent 10% SDS-polyacrylamide gel electrophoresis and immunoblotted onto a nitrocellulose membrane (Schleich er and Schuell, Keene, NH, USA). The membrane was then incubated with rabbit polyclonal anti-CYP2E1 (Cat. ab28146, 1:1000 dilution; Abcam), rabbit polyclonal antinuclear factor erythroid 2-related factor (Nrf)-2 (Cat. SC-13032, 1:1000 dilution; Santa Cruz), and rabbit polyclonal anti-oxygenase-1 (HO-1) (Cat. SC-10789, 1:1000 dilution; Santa Cruz) for 2 h. Bound antibodies were visualized using a chemiluminescent substrate (Miracle-Star™; iNtRON Biotech, Gyeonggi, Korea). After imaging, the membranes were stripped and reprobed with anti-β-actin. The optical density (OD; per mm2) of each band was measured and the density of the band relative to the density of the β-actin band was compared using ImageJ software.
Statistical analysis
The data were presented as mean ± standard error. One-way analysis of variance followed by the Student–Newman–Keuls post hoc test were used for multiple comparisons. In all cases, P < .05 was considered to indicate significance.
Results
Isolation and quantification of compounds by HPLC analysis
Among the isolates, compounds 1 and 2 were diastereomers of 3-[ethoxy-(methylthio)methyl]-2-pyrrolidinethione, and compound 3 was 3-(E)-(methylthio)methylene-2-pyrrolidinethione (3 M2P) (Fig. 1A). The extract was analyzed by HPLC to quantify the level of 3 M2P, a compound previously reported to have been isolated from radish. 20 The calibration curve comprised of the concentration versus peak area of the marker compound and indicated that the method was linear across the range of 0.78–50 μg/mL. The calibration results indicated that the correlation coefficient (r 2) of the line of best fit was >0.9999. The concentration of this compound was 147.5 μg/g BRE (Fig. 1B).
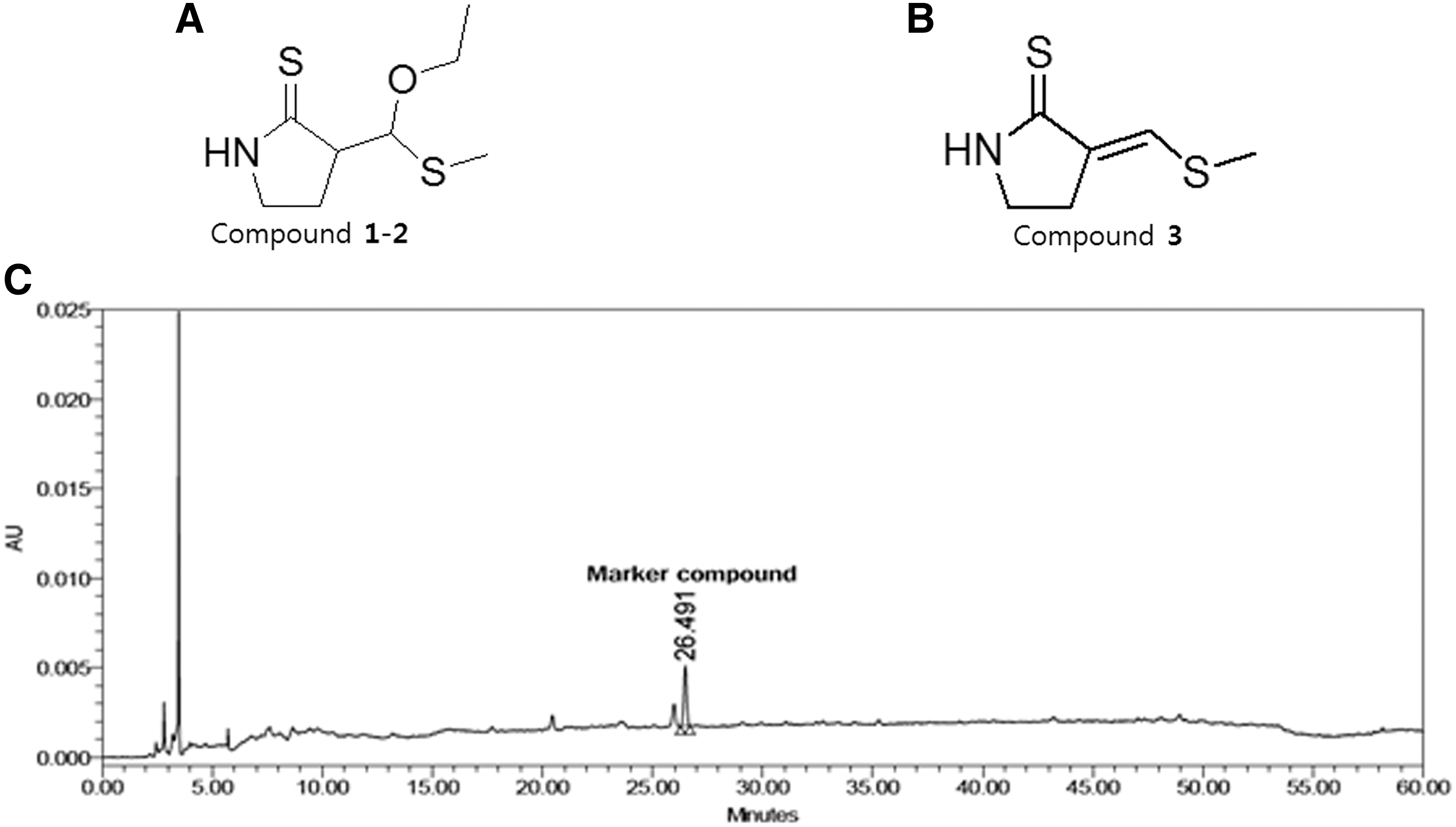
Chemical structures of compounds isolated from black radish root.
Effect of black radish on HepG2 cell viability
To measure the viability of BRE in HepG2 cells, we performed an MTT assay (Fig. 2A). HepG2 cells were treated with various concentrations (125, 250, 500, and 1000 μg/mL) of BRE for 24 h. The results showed that BRE at the range of various concentrations had no cytotoxicity on HepG2 cells and cell viability was approximately 100% compared to control cells. Thus, we used these concentrations of BRE in the following experiments.
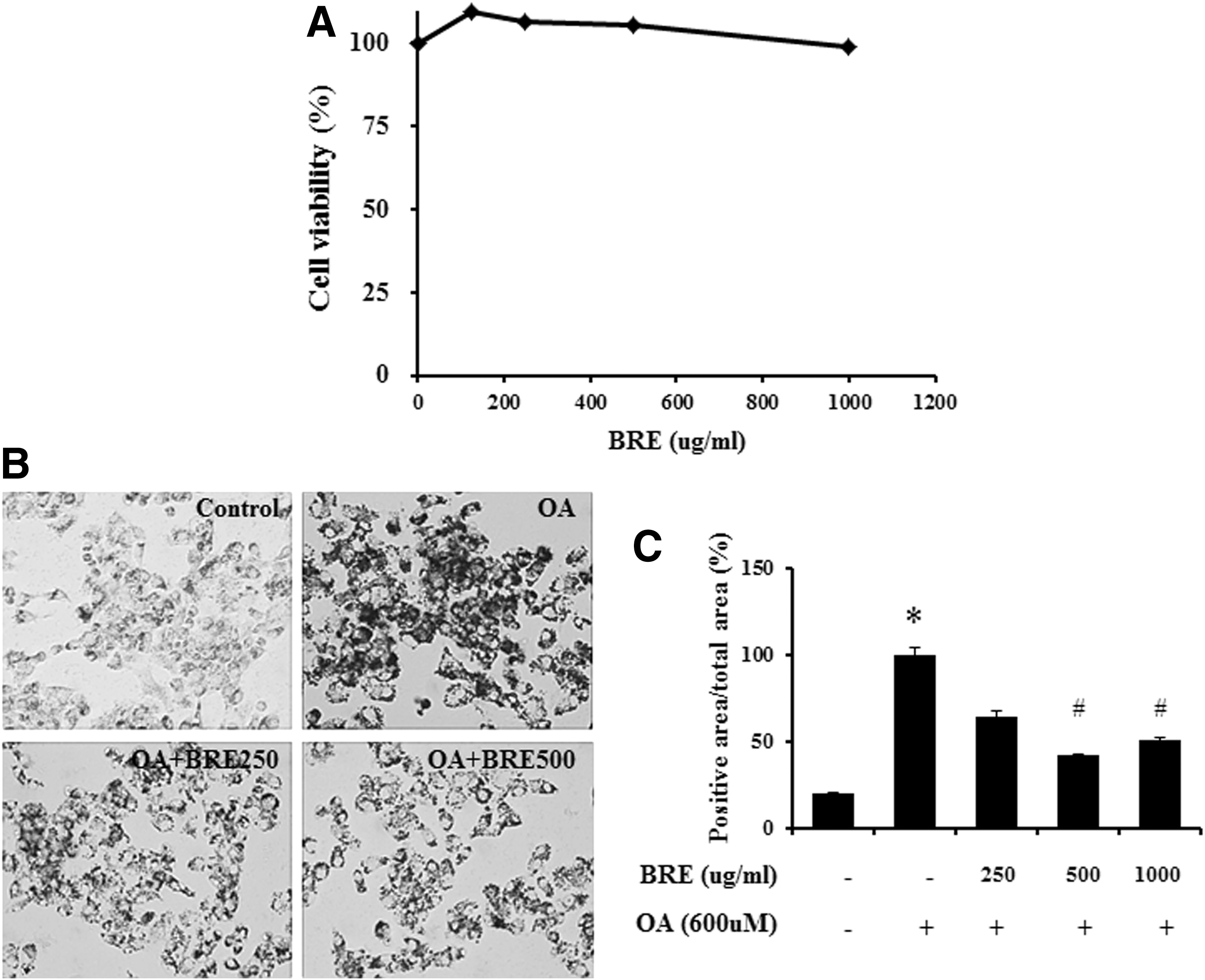
Effect of BRE on cytotoxicity and lipid accumulation in oleic acid-treated HepG2 cells.
Black radish pretreatment reduced lipid droplet accumulation in HepG2 cells and in rats with CCl4-induced liver injury
We confirmed that BRE treatment inhibited lipid accumulation in oleic acid-treated HepG2 cells and rat livers with CCl4-induced injury. BRE treatment at 500 and 1000 μg/mL attenuated lipid accumulation in oleic acid-treated HepG2 cells (Fig. 2A, B). The level of lipid accumulation was greatest in the vehicle-treated control group (arbitrarily set at 100%). In the BRE-treated group, lipid accumulation was significantly decreased in the groups treated with 500 μg/mL (42.3% ± 0.51%, P < .05) and 1000 μg/mL (50.4% ± 1.44%, P < .05) BRE compared with the vehicle-treated group (Fig. 2B). Similar results were observed in CCl4-treated rat livers (Fig. 3). Compared to pretreatment with vehicle, pretreatment with BRE significantly inhibited lipid droplet accumulation when used at doses of 500 mg/kg (47.5% ± 1.2%, P < .05) and 1000 mg/kg (48.0% ± 0.21%, P < .05) in livers of rats with CCl4-induced injury (Fig. 3F).
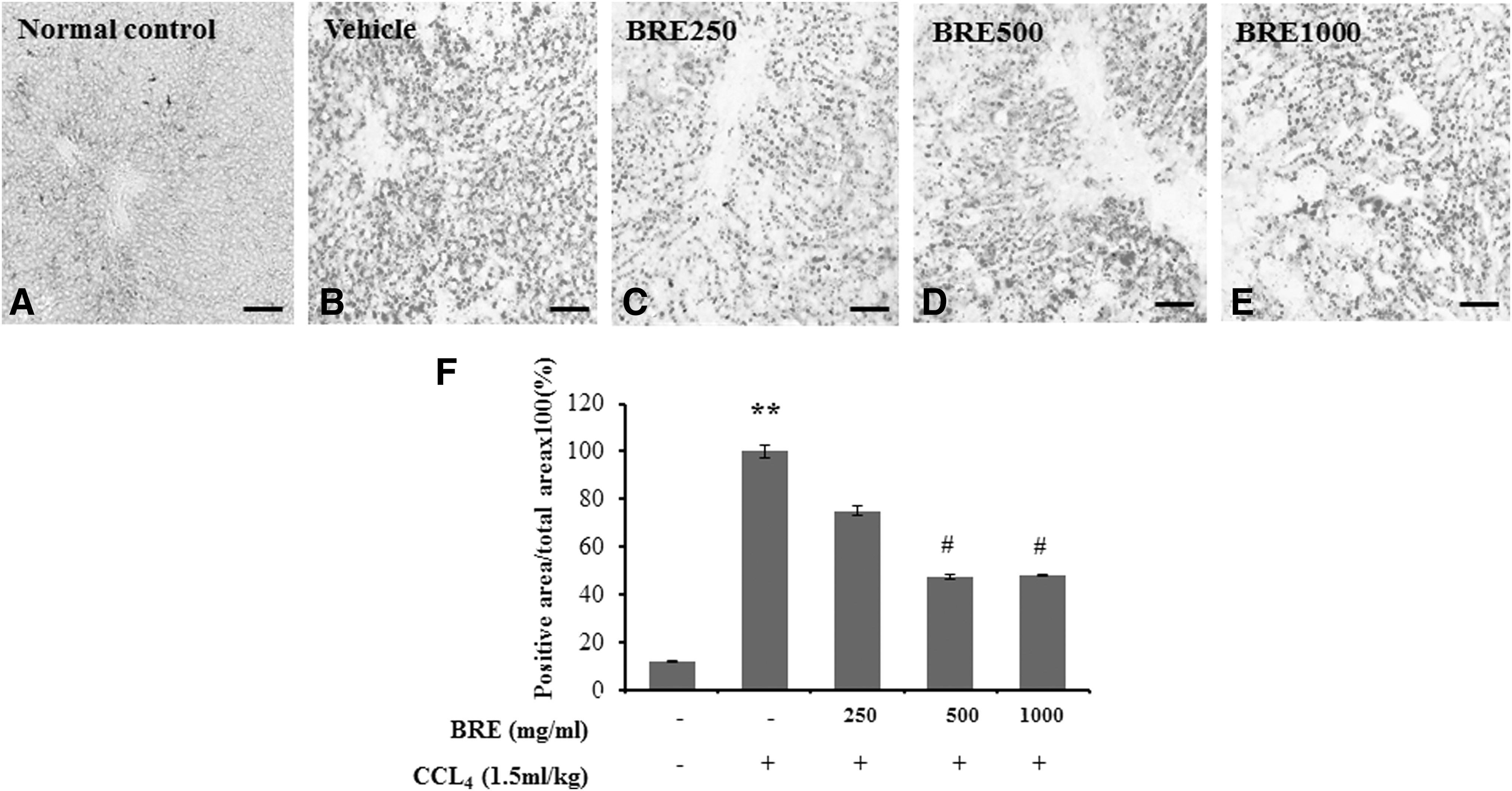
Effect of BRE on rat livers with CCl4-induced acute fatty changes. Liver sections were stained with Oil red O.
Black radish pretreatment reduced serum levels of ALT and AST in rat livers with CCl4-induced acute liver injury
Serum ALT and AST activities in controls were normal (Table 1). There were no significant differences in the levels of serum ALT or AST in normal rats treated with different doses of BRE (1000, 2000, and 4000 mg/kg), indicating that black radish does not have harmful effects on liver function in this model (Supplementary Table S1; Supplementary Data are available online at
Each value is represented as the mean ± SE.
P < .001 versus normal control group.
P < .05, ## P < .01, ### P < .001 versus vehicle-treated CCl4 group.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; TCHO, total cholesterol; GLU, glucose; BUN, blood urea nitrogen.
As shown in Table 1, biochemical analyses of serum enzymes were performed to verify the protective role of BRE during CCl4-induced hepatic injury. The activities of serum ALT and AST were significantly increased in vehicle-treated rat livers with CCl4-induced injury compared with normal controls (P < .001). However, in livers with CCl4-induced injury, pretreatment with 125, 250, 500, and 1000 mg/kg BRE significantly reduced the activities of serum ALT and AST (P < .05 and P < .001, respectively), which were significantly increased in the vehicle-treated rats (Table 1). There were no differences in the levels of total cholesterol, glucose, or blood urea nitrogen between the groups. These results suggest that black radish treatment protects the liver from CCl4-induced injury.
Effects of black radish treatment on antioxidant enzymes during CCl4-induced liver injury
To evaluate the antioxidant effects of black radish on CCl4-induced liver injury, the activities of antioxidant enzymes including SOD, CAT, and MDA were tested in liver tissues treated with black radish and CCl4 (Table 2). The treatment of vehicle to CCl4-treated rats showed significantly decreased CAT and SOD levels, and increased MDA levels compared with normal control rats. Conversely, the treatment of black radish before CCl4 treatment resulted in significant increases CAT and SOD levels, and decreased MDA levels compared with vehicle-treated CCl4-injured rats (Table 2).
Each value is represented as the mean ± SE.
P < .05, ** P < .01, *** P < .001 versus normal control group, # P < .05, ## P < .01 versus vehicle-treated CCl4 group.
BRE 125, black radish extract (125 mg/kg); MDA, malondialdehyde; SOD, superoxide dismutase; CAT, catalase.
Histopathological examination
There is evidence from histopathological studies that black radish protects against CCl4-induced liver injury (Fig. 4). In the normal control groups (Fig. 4A), the liver tissue showed a regular histological architecture, which included hepatocytes with a well-preserved cytoplasm, a prominent nucleus, and a central vein in the middle. CCl4-injured liver of rats revealed the extensive pericentral necrosis with loss of hepatic architecture and hemorrhage, infiltration of inflammatory cells in the parenchyma, and swelling of hepatocytes (Fig. 4B). The hepatic lesions of BRE-pretreated CCl4-injured rats were markedly reduced, compared with those in the vehicle-treated controls, and occurred in a BRE dose-dependent manner (Fig. 4C–F).
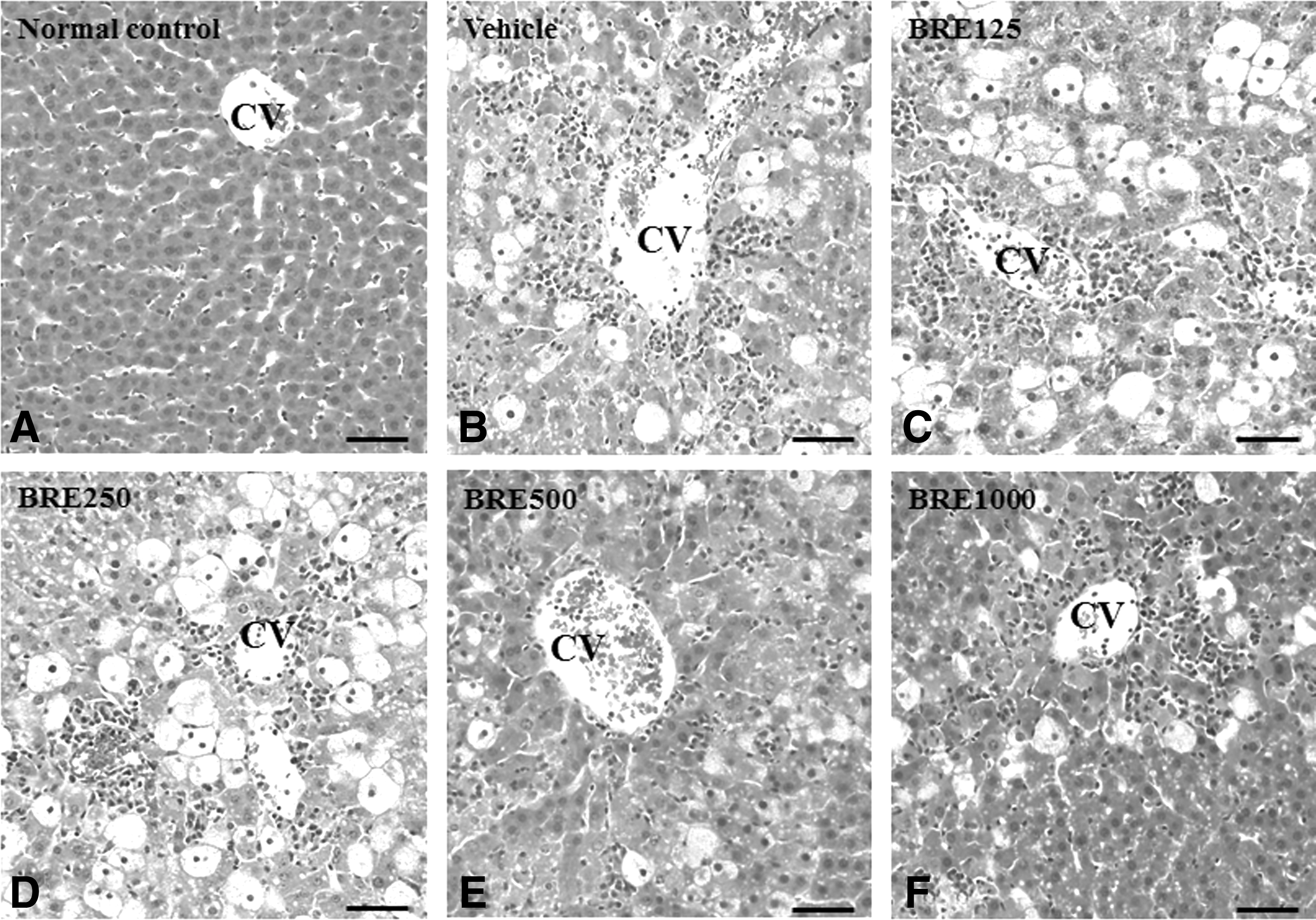
Histopathological results of livers from control and CCl4-injured rats with or without BRE pretreatment. Each panel shows a representative photograph from each group.
Black radish pretreatment enhanced CYP2E1 and HO-1 protein levels in livers of rats with CCl4-induced injury
Oxidative stress in liver injury is known to be important for CYP regulation 3 and HO-1, 5 and it has been confirmed that black radish can regulate the expression of CYP2E1 (Fig. 5A, C–F). The hepatic protein level of CYP2E1 was significantly decreased in the vehicle-treated group (relative OD, P < .05) compared with the normal control group. In contrast, pretreatment with BRE (250–1000 mg/kg, P < .05, respectively) significantly increased the level of CYP2E1 protein compared to pretreatment with vehicle (Fig. 5A). The protein level of HO-1 in the liver was lower in the vehicle-treated group than in the normal control group (P < .05; Fig. 5B). Pretreatment with 250 and 500 (P < .01) and 1000 mg/kg BRE (P < .05) significantly increased HO-1 levels compared to vehicle treatment (Fig. 5B).
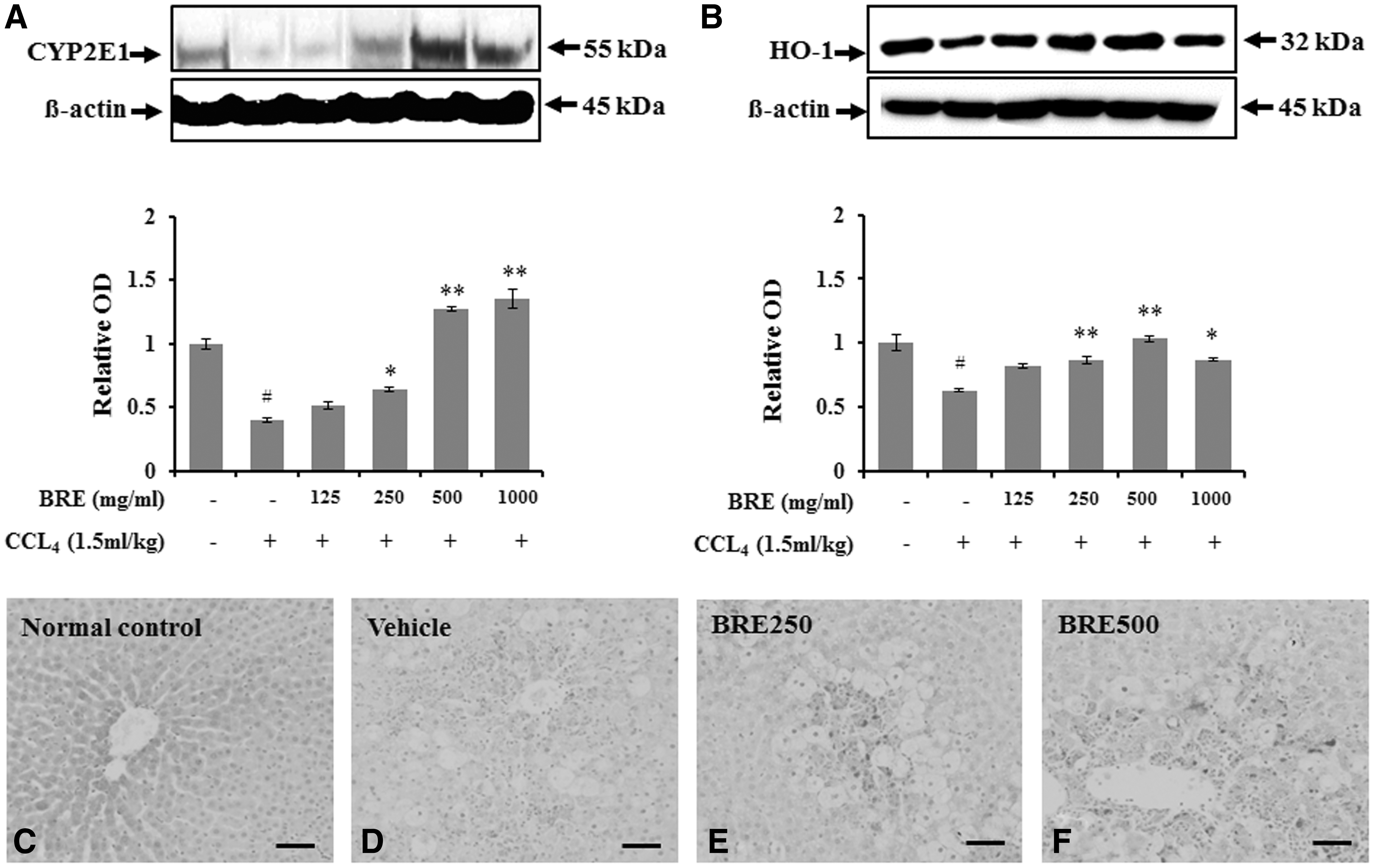
Western blot analysis of cytochrome p450 (CYP) 2E1 and HO-1 expression in CCl4-injured rats with or without BRE pretreatment. Representative immunoblots of
To determine the effect of black radish in expressed hepatocytes following CCl4-induced acute liver injury, the immunoreactivity of CYP2E1 in the liver was assessed. CYP2E1-positive regions were rare in the centrilobular area of normal livers (Fig. 5C). In vehicle-treated and CCl4-injured rats, CYP2E1 immunoreactivity was significantly decreased in the hepatic lobules (Fig. 5D). The degree of CYP2E1 immunoreactivity was markedly increased in the CCl4-injured liver tissue from rats pretreated with BRE (Fig. 5E, F).
Black radish pretreatment attenuated oxidative stress in HepG2 cells
Based on the antioxidant effects of BRE, the proteins CYP2E1, Nrf-2, and HO-1 in HepG2 cells were semi-quantitatively assessed by western blotting. CYP2E1 protein levels were significantly higher in the BRE-pretreated group than in the oleic acid control group (250 μg/mL, P < .05; Fig. 6A, B).
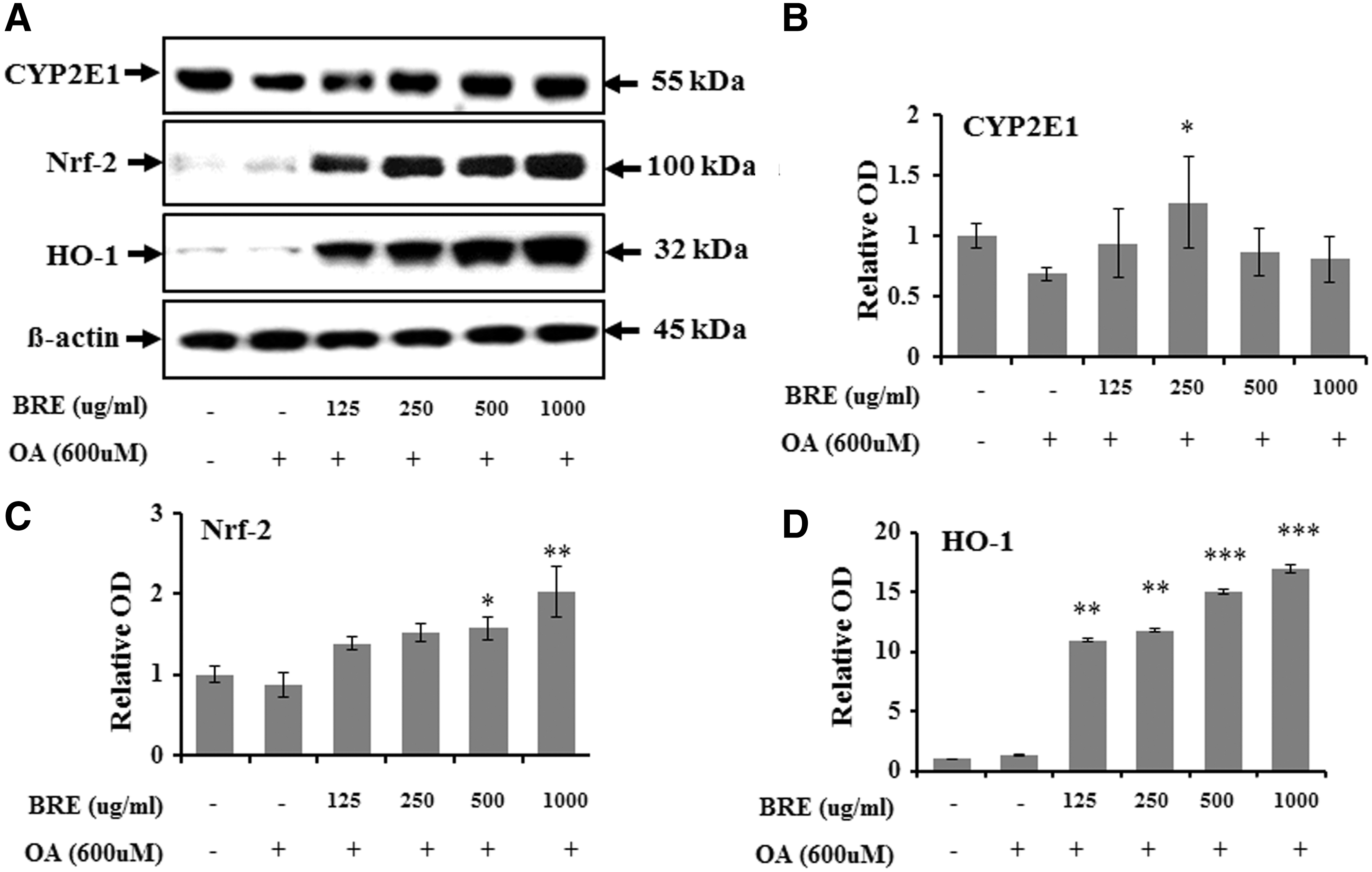
Western blot analysis of CYP2E1, Nrf-2, and HO-1 expression in oleic acid-treated HepG2 cells with or without BRE treatment.
The protein level of Nrf-2 and HO-1 in HepG2 cells was lower in the oleic acid-treated control group than the negative control group. However, pretreatment with BRE significantly increased Nrf-2 levels [500 μg/mL (P < .05); 1000 μg/mL (P < .01) vs. controls] (Fig. 6C) and HO-1 levels [125 and 250 μg/mL (P < .01); 500 and 1000 μg/mL (P < .001) vs. controls] (Fig. 6D) in oleic acid-treated HepG2 cells. Additionally, the radical scavenging activity of 3M2P, a compound isolated from black radish, on DPPH free radicals increased significantly with increasing BRE concentration in the different dosage treatments (Fig. 7A), and the level of Nrf-2 was significantly and dose-dependently higher in HepG2 cells pretreated with BRE than corresponding oleic acid-treated HepG2 cells (Fig. 7B).
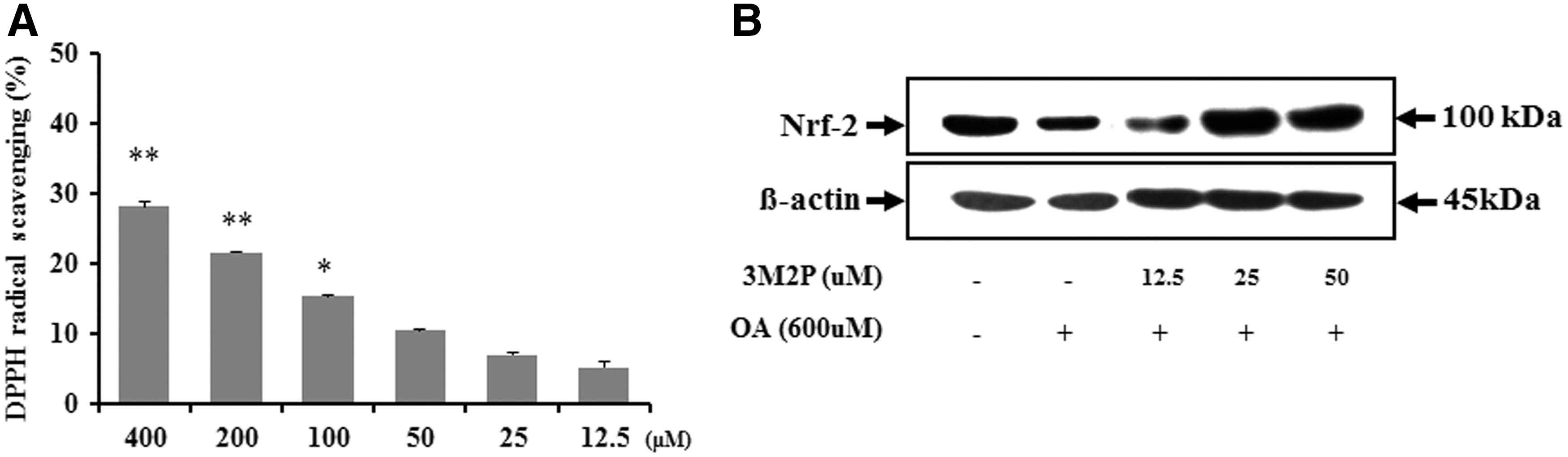
Radical scavenging activity and western blot analysis of Nrf-2 expressions in oleic acid-treated HepG2 cells with or without 3M2P treatment.
Discussion
This is the first time diastereomers of 3-[ethoxy-(methylthio)methyl]-2-pyrrolidinethione and 3-(E)-(methylthio)methylene-2-pyrrolidinethione (3M2P) were isolated from BRE. A previous study showed that silica gel chromatography of the dichloromethane extract of radish roots afforded 3M2P, a mixture of 4-methylthio-3-butenyl isothiocyanate and 4-(methylthio)butyl isothiocyanate, β-sitosterol, β-sitosteryl-3β-glucopyranoside-6′-O-palmitate, monoacylglycerols, and a mixture of α-linolenic acid and linoleic acid. 20 In our study, 3M2P isolated from BRE was the same compound previously isolated from radish extract. We postulated that 3M2P might be a marker compound in BRE.
In this study, we investigated whether BRE can inhibit accumulation of lipid droplets in rats with hepatic steatosis induced by oleic acid. BRE inhibited accumulation of lipid droplets in HepG2 cells without causing cytotoxicity. As for the cause of reduced accumulation of lipid droplets, we propose that BRE reduced accumulation of lipid droplets in oleic acid-induced steatosis of HepG2 cells and rat liver with CCl4-induced injury via reduction of CYP2E1 expression and activation of Nrf-2 and HO-1, leading to protection from oxidative stress caused by oleic acid-induced injury.
Antioxidant and cytoprotective molecules are known to protect hepatocytes against oxidative stress. 21 In this study, the hepatoprotective effects of black radish were further supported by a reduction in the levels of both ALT and AST in rats treated with BRE and CCl4 compared with vehicle-treated CCl4 controls. In addition, we analyzed the level of MDA, SOD, and CAT in livers with and without BRE pretreatment. The findings suggest that BRE pretreatment induces the downregulation of lipid peroxidation and the upregulation of SOD and CAT in the CCl4-injured livers, thus protecting the liver from CCl4-induced oxidative injury. These results agree with those of previous hepatoprotection experiments. 1,6
The expression and activation of CYP subfamily members, including CYP1A2, CYP2E1, CYP2B1, CYP2C, and CYP3A, were decreased by CCl4 injection in animal models, 3,22 indicating that the CYP subfamily is involved in hepatotoxicity. Therefore, CCl4 may cause the destruction of liver cell structure by reducing the level of CYP2E1. 22 In contrast, CYP2E1 expression was significantly higher in the BRE-pretreated group than the CCl4-injured group, and was BRE dose dependent. These results suggest that BRE mediates its protective effect by increasing CYP2E1 expression during CCl4-induced liver injury.
Next, we investigated the relationship between CYP2E1 expression and the degree of oxidative stress. Pretreatment with BRE protected against oxidative stress by restoring the levels of Nrf-2 and HO-1 in oleic acid-treated HepG2 cells, as in the various oxidative stress conditions in lung, liver, and nervous tissues. 6,23 In our study, Nrf-2 expression in the oleic acid-treated controls was lower than in normal controls with a concomitant increase in the level of HO-1 and its expression was significantly enhanced by BRE treatment. Our results showed that BRE treatment upregulated Nrf-2 expression with a concomitant increase in HO-1 expression in oleic acid-treated HepG2 cells. In addition, as a consequence of confirming the effect of 3M2P, we also confirmed that the increase in Nrf-2 expression was a result of BRE treatment.
Collectively, BRE treatment reduced lipid accumulation and ameliorated the subsequent inactivation of the CYP isoenzyme in rat livers with CCl4-induced liver injury. This may occur through the upregulation of antioxidant and radical scavenging enzymes, which are inversely related to ALT and AST levels. Overall, BRE may play a potential role as an Nrf-2 activator for nonalcoholic fatty disease.
Footnotes
Acknowledgments
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry, and Fisheries (IPET) through Agri-Bio Industry Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (Grant number: 316006-05-1-HD040).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.