
Abstract
Endoplasmic reticulum (ER) stress-related unfolded peptide accumulation is closely associated with the development of neurodegenerative diseases known as protein misfolding disorders. The antioxidative properties of kimchi, a traditional Korean fermented vegetable dish, have been well established. In this study, the neuroprotective effects of the kimchi methanol extract (KME) were examined in high-cholesterol diet (HCD)-fed mice. The animals were fed a HCD, with oral administration of either KME (KME group, 200 mg·kg bw−1·day−1, n = 10) or distilled water (Control group, n = 10) for 8 weeks. Compared with the levels in the control group, the reactive oxygen species, peroxynitrite, and lipid peroxidation levels in the brain were significantly decreased in the KME group (P < .05), whereas the glutathione level was increased (P < .05). In addition, the ER stress biomarkers, phospho-eukaryotic initiation factor 2 subunit α, glucose-regulated protein 78, X-box binding protein 1, inositol-requiring enzyme 1, and C/EBP homologous protein and the nuclear factor-kappaB-mediated inflammation were significantly reduced in the KME group (P < .05). In contrast, the expression levels of antioxidative enzymes regulated by nuclear factor erythroid 2-related factor-2 were elevated (P < .05). The amyloid-beta expression levels of the KME group were lower than that of the control group (P < .05). Moreover, the expression levels of Bcl-2-associated X, and caspases-3 and −9 were downregulated, with a concomitant upregulation of B cell lymphoma 2 (P < .05). Accordingly, KME provide neuronal cell protection via suppressing ER stress and caspase cascade signaling.
Introduction
R
Unfolded protein response (UPR) is initiated to restore the normal function of the ER by activation of membrane sensors such as protein kinase-like endoplasmic reticulum kinase (PERK), inositol-requiring enzyme 1 (IRE-1), and activating transcription factor 6 (ATF6) against unfolded peptide accumulation in the ER. As a consequence, the signal transduction mediated through UPR reset the gene expressions involved in protein folding, protein secretion, lipid synthesis, and protein degradation. 8 In fact, normalization to the function of secretory pathways in the ER of the neuronal cell could alter the pathological conditions. 2,9 Nevertheless, when UPR does not properly manifest its action, apoptosis will occur through upregulation of C/EBP homologous protein (CHOP) that is found in dead cells. 10
In HCD-fed mice, the cerebral cortex and hippocampus exhibit increased expression of caspase-3, which is a well-known biomarker for apoptosis. 11 Moreover, HCDs induce the formation of amyloid beta (Aβ) in the brain, which worsens the progression of neurodegeneration. 5 Consequently, HCD-induced neurotoxicity would cause a brain dysfunction via oxidative stress–inflammation-induced apoptosis. 5
Kimchi is a traditional Korean fermented vegetable food made with Korean cabbage, green onion, red pepper powder, garlic, ginger, and fermented fish sauce. 12 In our previous study, quercetin, capsaicin, ascorbic acid, and 3-(4′-hydroxyl-3′,5′-dimethoxyphenyl)propionic acid (HDMPPA) were identified in cabbage kimchi. 13 Besides these active compounds, lactic acid bacteria (∼109 CFU/mL kimchi extract) were also detected from the ripened kimchi. The health benefits of kimchi have been extensively studied, including its prevention of oxidative stress, 14 inflammation, 13 cancer, 15 and lipid-lowering effects. 16 In addition, kimchi was shown to protect against Aβ-induced memory loss and cognitive deficits, where the beneficial effects were mediated through alleviation of oxidative stress. 17 These effects are generally attributed to the antioxidative properties of kimchi. Moreover, kimchi decreased hepatic apoptosis in low-density lipoprotein receptor knockout mice (LDLr KO mice) via the suppression of ER stress and the upregulation of antioxidative enzymes regulated by nuclear factor erythroid 2-related factor-2 (Nrf2). 13 In this study, we investigated the inhibitory effects of a kimchi methanol extract (KME) and its mechanism of action on oxidative stress, inflammation, ER stress, and the caspase cascade in the brain of HCD-fed mice.
Materials and Methods
Preparation of kimchi and its methanol extract
Brined cabbage (84.9%) was mixed with kimchi condiments comprising red pepper powder (2.6%), garlic (2.5%), green onion (2.3%), ginger (0.5%), fermented fish sauce (3.0%), sugar (0.5%), and glutinous rice paste (3.7%). 12 The kimchi was fermented at 4°C in a kimchi refrigerator (R-K182PM; LG, Seoul, Korea) until the pH reached 4.3 ± 0.1, after which it was freeze-dried (SFDSM06; Samwon Freezing Engineering Co., Busan, Korea). Methanol extraction of the kimchi was carried out three times, using 10 volumes of 70% methanol for 24 h at room temperature each time. The KME was concentrated using a rotary evaporator (R-200; Buchi, Flawil, Switzerland), freeze-dried, and stored at −80°C for further use.
Animal study
LDLr KO mice (males, 5 weeks old) were purchased from Jackson Laboratories (Bar Harbor, ME, USA). After acclimation for 1 week, the animals were divided into two groups on the basis of body weight. They were raised individually under controlled conditions with a 12-h light-dark cycle, humidity of 50% ± 5%, and temperature of 23°C ± 1°C. Food intake was checked daily and the body weight was measured weekly. The mice were fed a HCD for 8 weeks, with administration of either distilled water as a vehicle (control group, n = 10) or KME at a dose of 200 mg·kg bw−1·day−1 (KME group, n = 10), based on the previous study. 17 The administration of 200 mg/kg bw to mouse is equivalent to 15 g kimchi consumption by 60 kg adult. The HCD containing 1.25% cholesterol was prepared according to the guidelines of Paigen's atherogenic rodent diet. 18 The amount of KME administered to the mice was based on a previous study showing its beneficial effects in overcoming Aβ-induced cognitive deficits in mice. 17 Upon completion of the 8-week KME treatment, the mice were anesthetized by CO2 asphyxiation after a 12-h fasting period. The brain was excised, rinsed with phosphate-buffered saline (PBS), immediately placed on liquid nitrogen, and then stored at −80°C for further use. The animal study was conducted in accordance with the guidelines for animal experiments and approved by the Pusan National University Institutional Animal Care and Use Committee (PNU-IACUC, Approval Number PNU-2016-1063).
Plasma lipid profile
Plasma triglyceride (TG) was determined using a kit (AM 157-K; Asan Pharm, Seoul, Korea) using a modified lipase-glycerol phosphate oxidase method. Total cholesterol (TC) concentration was determined using a commercial kit (AM 202-K, Asan Pharm).
Determination of cholesterol, lipid peroxidation, and oxidative stress markers in the brain
Brain homogenates were prepared in PBS (1:9, w/v). Cholesterol concentration was determined in the brain from lipids extracted from tissue homogenates by Folch's method. 19 For brain TC assay, the same kit used for the determination of plasma TC was used.
The thiobarbituric reactive substance (TBARS) level in the homogenates was also determined and expressed as the malondialdehyde concentration, 20 and the glutathione (GSH) concentration was measured using 5,5′-dithiobis(2-nitrobenzoic acid). 21
A postmitochondrial fraction was obtained through centrifugation of the brain homogenates at 3,012 × g for 15 min at 4°C, followed by a second centrifugation at 18,627 × g for 20 min at 4°C. The upper layer was used for the following experiments. The ROS level was measured using 2′,7′-dichlorofluorescein diacetate, 22 and peroxynitrite (ONOO−) level was determined using dihydrorhodamine buffer. 23 Changes in fluorescence of the reaction mixture were monitored for 30 min, at an excitation wavelength of 485 nm and an emission wavelength of 530 nm.
Western blot analysis
Proteins were extracted from the brain tissue by using lysis buffer and were then separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis according to a previously described method.
12
Protein expression was visualized by the enhanced chemiluminescence method, detected using the CAS-400 system (Core Bio, Seoul, Korea), and then calculated using ImageJ software (
The primary antibodies used in this study were as follows: α-tubulin (ab52866; Abcam, Inc., Cambridge, United Kingdom), β-actin (ab8226, Abcam, Inc.), Nrf2 (sc-13032; Santa Cruz Biotechnology, Santa Cruz, CA, USA), catalase (CAT, sc-34285; Santa Cruz Biotech.), heme oxygenase-1 (HO-1, sc-10789; Santa Cruz Biotech.), GSH S-transferase (GST, sc-138; Santa Cruz Biotech.), GSH peroxidase-1 (GSH-Px, sc-133160; Santa Cruz Biotech.), superoxide dismutase-1 (SOD, sc-11407; Santa Cruz Biotech.), nuclear factor-kappaB (NF-κB, sc-109; Santa Cruz Biotech.), cyclooxygenase-2 (COX-2, sc-1747; Santa Cruz Biotech.), inducible nitric oxide synthase (iNOS, sc-651; Santa Cruz Biotech.), Aβ (sc-58508; Santa Cruz Biotech.), phospho-eukaryotic initiation factor 2 subunit α (p-eIF2α; phospho S51, ab32157; Abcam, Inc.), glucose-regulated protein 78 (GRP-78, sc-1050; Santa Cruz Biotech.), X-box binding protein 1 (XBP-1, sc-7160; Santa Cruz Biotech.), IRE-1 (PA1-16927; Thermo Scientific, Waltham, MA, USA), CHOP (#2895; Cell Signaling Technology, Beverly, MA, USA), Bcl-2-associated X (Bax, sc-493; Santa Cruz Biotech.), caspase-3 (ab32351-100; Abcam, Inc.), caspase-9 (ab63488-100; Abcam, Inc.), and B cell lymphoma 2 (Bcl-2, sc-7382; Santa Cruz Biotech.).
The following horseradish peroxidase-conjugated secondary antibodies were purchased from Abcam, Inc.: rabbit anti-goat IgG H&L (ab6741), donkey anti-rabbit IgG H&L (ab6802), and rabbit anti-mouse IgG H&L (ab6728).
Statistical analysis
Statistical analyses were performed using SPSS version 23 (SPSS, Inc., Chicago, IL, USA). Data are presented as the mean ± SD. Statistical significances between two groups were analyzed by Student's t-test, and P < .05 was considered as being significant.
Results
Body weight gain, food efficacy, and brain weight
As shown in Table 1, the body weight gain, food efficacy ratio, and brain weight values between the two groups were not statistically different.
Data are the mean ± SD (n = 10 each group).
The control group was fed a HCD with oral administration of distilled water for 8 weeks. The KME group was fed a HCD with oral administration of the KME (200 mg·kg bw−1·day−1) for 8 weeks.
Total weight gains were divided by total food intakes.
Data between the two groups are not significantly different.
KME, kimchi methanol extract; HCD, high-cholesterol diet.
Lipid, ROS, ONOO−, TBARS, and GSH concentrations
As shown in Table 2, increased plasma TG (P < .01) and TC (P < .01) were reduced by kimchi by 31.80% and 13.95%, respectively, compared with the control group. Compared with the levels in the control group, TC concentrations in the KME group were significantly decreased by 14.01% (P < .05). The ROS, ONOO−, and TBARS concentrations of the KME group were significantly lower by 23.14%, 13.50%, and 19.67%, respectively, whereas the GSH concentration was increased by 118.60%, relative to the control levels (P < .05 for all).
Data are the mean ± SD (n = 10 each group).
See the footnote of Table 1 for the description of the control and KME groups.
Significant difference between the two groups was analyzed by Student's t-test; * P < .05 and ** P < .01.
TG, triglyceride; TC, total cholesterol; ROS, reactive oxygen species; ONOO−, peroxynitrite; TBARS, thiobarbituric reactive substance; GSH, glutathione; Flu, fluorescence.
Expression of Nrf2- and NF-κB-regulated enzymes and Aβ
As shown in Figure 1A, the expression level of Nrf2 in the KME group was higher than that in the control group (P < .05). Subsequently, the CAT, HO-1, GST, and GSH-Px (all regulated by Nrf2) levels in the KME group were significantly increased by 111.61%, 120.31%, 123.91%, and 125.23%, respectively, relative to the control group levels. As shown in Figure 1B, the NF-κB expression level was lower in the KME group (by 29.63%) than in the control group. As a result, the expression levels of COX-2 and iNOS (both regulated by NF-κB) were decreased by 10.07% and 15.84%, respectively, compared with the control group levels (P < .05). Aβ expression was significantly lower in the KME group (by 23.92%) than in the control group (Fig. 1C).
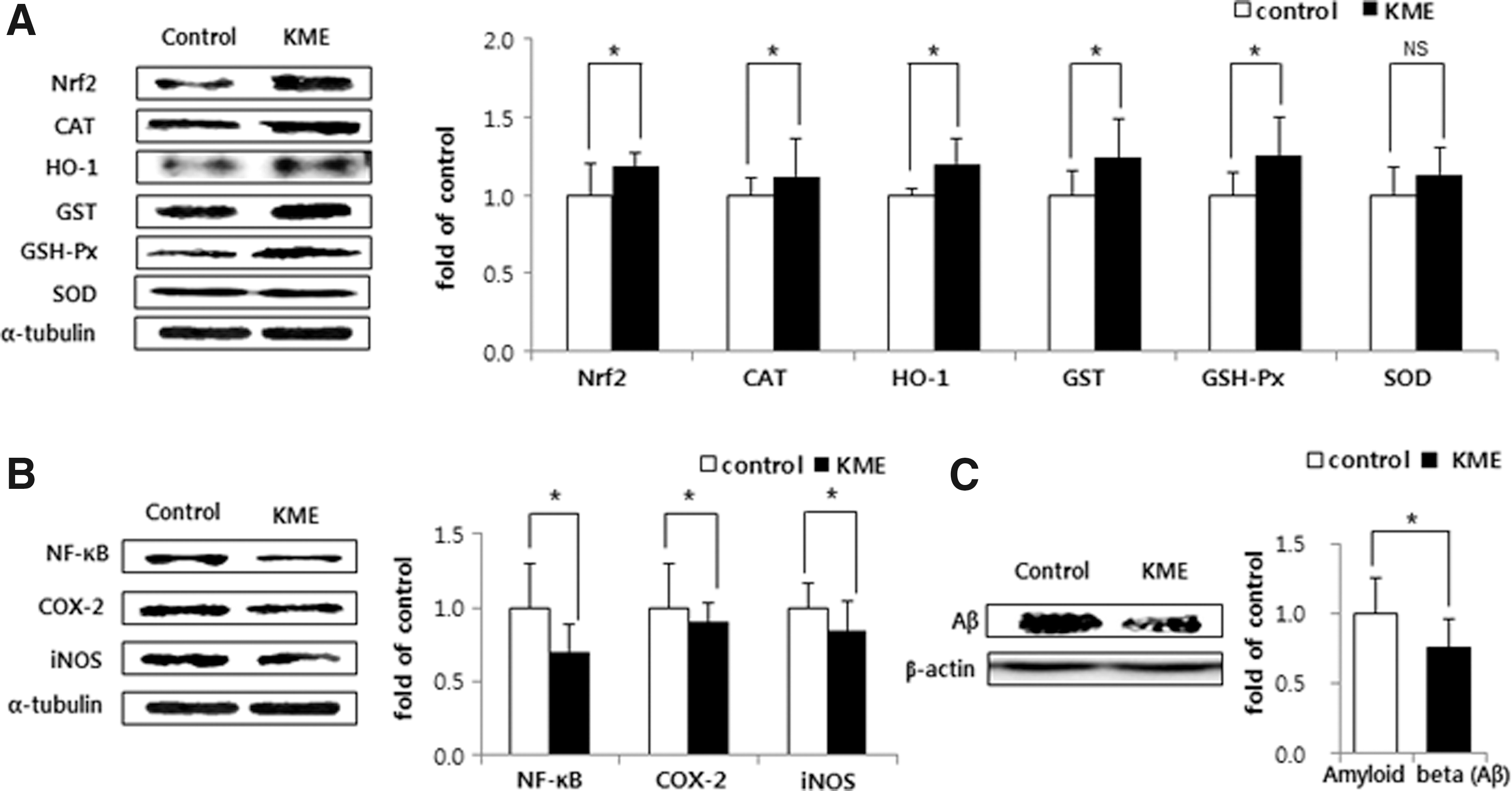
Protein expression levels in the brain of low-density lipoprotein receptor knockout mice fed a high-cholesterol diet for 8 weeks.
Expression of ER stress markers
As shown in Figure 2, the protein expression levels of ER stress markers such as p-eIF2α, GRP-78, XBP-1, IRE-1, and CHOP were significantly lower in the KME group by 24.87%, 33.19%, 21.20%, 20.59%, and 21.51%, respectively, than those in the control group.

Expression of endoplasmic reticulum stress-related factors; p-eIF2α, GRP-78, XBP-1, IRE-1, and CHOP in the brain of low-density lipoprotein receptor knockout mice fed a high-cholesterol diet for 8 weeks. See the footnote of Table 1 for the description of the control and KME groups. Data are the mean ± SD (n = 10 each group). Significant difference between the two groups was analyzed by Student's t-test; *P < .05 and **P < .01. p-eIF2α, phospho-eukaryotic initiation factor 2 subunit α; GRP-78, glucose-regulated protein 78; XBP-1, X-box binding protein 1; IRE-1, inositol-requiring enzyme 1; CHOP, C/EBP homologous protein.
Protein expression of caspase-3-related biomarker pathways
As shown in Figure 3, apoptosis markers that participate in the caspase cascade, such as Bax, caspase-3, and caspase-9, were downregulated by 18.28%, 16.55%, and 28.48%, respectively, whereas Bcl-2 expression was upregulated by 133.68%, relative to the control group levels (all P < .05).
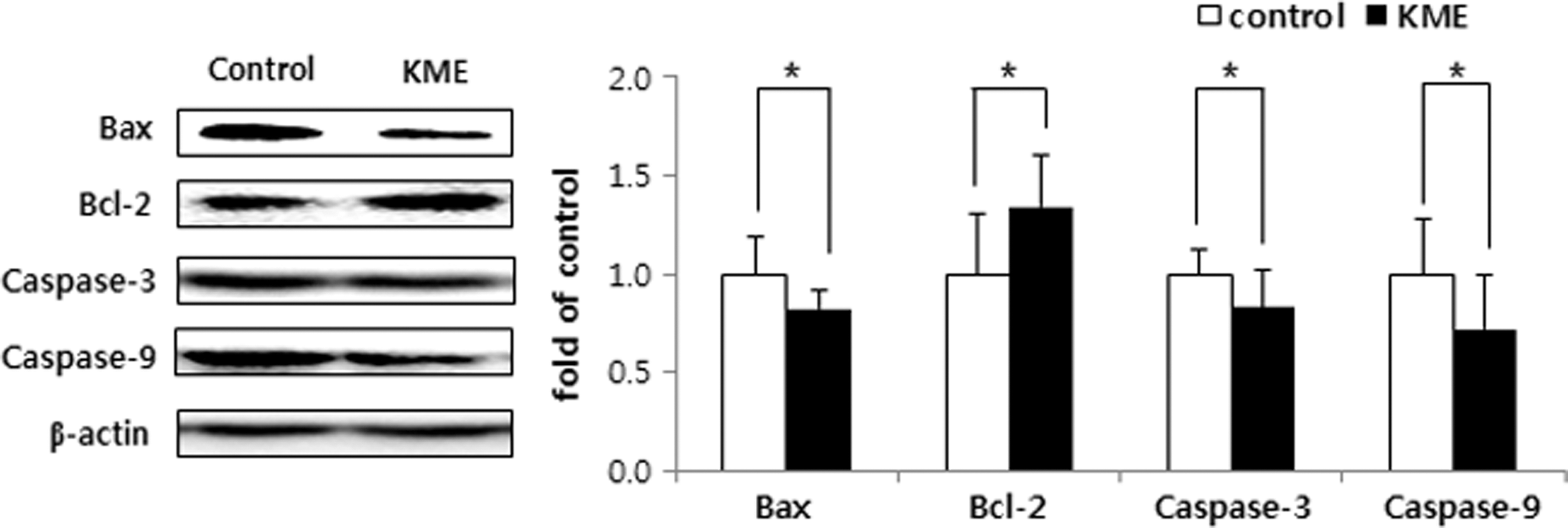
Expression of apoptosis-related factors; Bax, Bcl-2, caspase-3, and caspase-9 in the brain of low-density lipoprotein receptor knockout mice fed a high-cholesterol diet for 8 weeks. See the footnote of Table 1 for the description of the control and KME groups. Data are the mean ± SD (n = 10 each group). Significant difference between the two groups was analyzed by Student's t-test; *P < .05. Bax, Bcl-2-associated X; Bcl-2, B cell lymphoma 2.
Discussion
The brain is an organ in which oxidative stress could be easily elevated, because this organ consumes large amounts of oxygen (relative to the other organs) for energy production to conduct various brain functions. In addition, the brain contains a high concentration of polyunsaturated fatty acids, which are susceptible to lipid oxidation. 24 Furthermore, hypercholesterolemia is a known dietary factor that induces brain damage by apoptosis mediated through oxidative stress and ER stress. 5 Therefore, the antioxidative status of the brain is important to protect against neuronal cell damage from oxidative stress. 25 Furthermore, increased oxidative stress and Aβ accumulation in the brain are strongly associated with and has been a well-known cause of detrimental neuronal disorders including Alzheimer's disease, dementia, and Parkinson's disease. 26 In most tissues, including brain, oxidative balance is maintained by antioxidant material and enzymes such as SOD, CAT, HO-1, GST, and GSH-Px that are all regulated by Nrf2. 25,27 In brain injury, Nrf2 expression is diminished and the antioxidative enzymes are subsequently downregulated. 27 Therefore, upregulation of antioxidant status is important to prevent and/or retard the progress of neurodegenerative disorders. In the present study, KME was found to augment the Nrf2 expression in the HCD-fed mice, and its downstream protein expressions of CAT, HO-1, GST, and GSH-Px were upregulated. Consequently, the ROS and ONOO−levels and TBARS concentration elevated by HCD were diminished by KME administration, whereas GSH (an endogenous antioxidant) was increased. Moreover, the level of Aβ expression was significantly lower in the brain of KME-fed hypercholesterolemic mice. There are a lot of bioactive compounds in KME, as was confirmed by HPLC analysis. 13 Among these compounds, capsaicin has previously been found to inhibit neuronal degeneration in the mouse model of Parkinson's disease, 28 and ascorbic acid demonstrated protective effects on the neuronal cell in Alzheimer's disease. 29 Therefore, the current results demonstrated that bioactive compounds in KME might contribute to enhancement of the antioxidative status in the brain through the Nrf2 pathway.
Neuroinflammation is one of the immediate causes for the Aβ accumulation that impairs brain function and subsequently brings about cognitive deficits. 30 Hypercholesterolemia has been shown to increase NF-κB-mediated neuroinflammation in the brain of LDLr KO mice. 5,31,32 In this study, the HCD-induced elevation of NF-κB expression and that of its target enzymes, including iNOS and COX-2, were decreased upon KME administration. In the previous study, HDMPPA, a bioactive compound found in the cabbage kimchi, exhibited anti-inflammatory effects in microglial cells via inhibition of the NF-κB signaling pathway. 33 In addition, capsaicin, 28 ascorbic acid, 34 and quercetin 31 have revealed inhibitory effects on inflammation in the brain of different animal models. Based on these numerous reports, the anti-inflammatory effects of KME in the brain are expected, because quercetin, HDMPPA, ascorbic acid, and capsaicin are known components of KME. 13
ER stress occurs when unfolded peptides accumulate in the ER, which condition subsequently requires UPR to resolve the problem. UPR is initiated through membrane sensors such as PERK, ATF6, and IRE-1 followed by distinct signaling cascades 35 to upregulate genes involved in peptide folding and protein secretion. However, UPR works improperly, and then apoptosis occurs to determine the problem in the cell via production of CHOP. 10 In addition, ER membrane sensor IRE-1 activates JUN amino-terminal kinase to precede the apoptosis. Dysregulation of UPR signaling has been reported to play a critical role in the development of the neurodegenerative disease 36,37 that is recently classified as a protein misfolding disorder. 1,2 In this study, KME significantly suppressed ER stress via downregulation of ER stress markers such as GRP78, p-eIF2α, IRE-1, CHOP, and XBP-1. Our results are consistent with the previous studies. KME demonstrated inhibitory effects on the expression of hepatic ER stress markers in LDLr KO mice, in which GRP78, p-PERK, p-eIF2α, XBP-1, and CHOP expression levels were significantly reduced. 13 In addition, GRP78 and ATF6 were downregulated upon KME and HDMPPA treatment in 7-ketocholesterol-treated macrophages. 38 Furthermore, administration of quercetin, a bioactive compound found in the KME inhibited IRE-1 and PERK activation in the brain. 39 Similarly, the inhibition of oxidative stress decreased the risk of neurodegenerative disease through suppression of the ER stress and apoptosis. 40
Neuronal cell death underlies the symptoms of numerous human neurological disorders due to excessive apoptosis. 11 Mitochondrial-dependent apoptosis occurs under the condition of an unbalanced expression of proapoptotic (Bax, caspase-9, and caspase-3) and antiapoptotic (Bcl-2) factors. The caspase cascade is initiated through caspase-9 activation, as a result of the dissociation of Bax and Bcl-2. 41 In HCD-fed mice, caspase-3 expression was elevated in the cerebral cortex and hippocampus. 11 In this study, compared with those in the control group, the expression levels of Bax, caspase-9, and caspase-3 were significantly decreased in the KME group with a concomitant increase of Bcl-2 expression, indicating that neuronal cell death induced by HCD might be prevented by KME administration. These antiapoptotic effects of KME might be attributed to the bioactive compounds. Capsaicin, 42 quercetin, 41 and vitamin C, 43 which are found in KME, attenuated apoptosis by inhibiting oxidative stress and the caspase-3 pathway in the neurons of rats.
In conclusion, this study demonstrated potential benefits of KME on the protection of neuronal cells by attenuation of ER stress and apoptosis in the brain of HCD-fed mice. We did not have histological data on neuronal cell death; however, Aβ expression in the brain was reduced and the caspase cascade signaling pathway was significantly suppressed. The neuroprotective activity of KME might be a result of its bioactive compounds contributing to enhancement of the antioxidative and anti-inflammatory status in the brain. Further advanced studies on the protective effects of the bioactive compounds in KME are needed; in particular, on their improvement of memory loss and cognitive deficits manifested in neurodegenerative diseases.
Footnotes
Acknowledgment
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2015R1D1A1A01057267).
Author Disclosure Statement
No competing financial interest exists.