
Abstract
The inflammatory cytokine tumor necrosis factor α (TNFα), upregulated in the obese condition, promotes protein degradation and is implicated in obesity-related skeletal muscle atrophy and age-related sarcopenia. Quercetin, a flavonoid, elicits antioxidative and anti-inflammatory activities. In this study, we investigated the effect of quercetin on TNFα-induced skeletal muscle atrophy as well as its potential mechanism of action. In this study, we observed that quercetin suppressed expression of TNFα-induced atrophic factors such as MAFbx/atrogin-1 and MuRF1 in myotubes, and it enhanced heme oxygenase-1 (HO-1) protein level accompanied by increased nuclear translocation of nuclear factor erythroid 2-related factor 2 (Nrf2) in myotubes. The HO-1 inhibitor ZnPP suppressed the inhibitory actions of quercetin on TNFα-induced atrophic responses and degradation of IκB-α in myotubes. Moreover, quercetin supplementation to high-fat diet-fed obese mice inhibited obesity-induced atrophic responses in skeletal muscle, accompanied by upregulation of HO-1 and inactivation of nuclear factor-kappa B (NF-κB), and the quercetin actions were attenuated in Nrf2-deficient mice. These findings suggest that quercetin protects against TNFα-induced muscle atrophy under obese conditions through Nrf2-mediated HO-1 induction accompanied by inactivation of NF-κB. Quercetin may be used as a dietary supplement to protect against obesity-induced skeletal muscle atrophy.
Introduction
O
Heme oxygenase-1 (HO-1) is the rate-limiting enzyme in the conversion of heme into biliverdin, releasing free iron and carbon monoxide, and plays a critical role in the prevention of oxidative stress, inflammation, and metabolic dysregulation. 13 Nuclear factor erythroid 2-related factor 2 (Nrf2), a basic leucine zipper transcription factor, is a major regulator of HO-1 expression that protects against oxidative damage and inflammation. It was previously reported that induction of HO-1 was able to suppress inflammatory cytokine expression in ob/ob mice and diabetic rats, 14 –16 reduces obesity-induced inflammation by polarizing macrophages in obese adipose tissue and liver, 17,18 and improves insulin resistance in skeletal muscle of high-fat-fed mice. 14,19 Hemin, an HO-1 inducer, was shown to reduce expression of inflammatory cytokines genes/proteins as well as oxidative stress in skeletal muscle of high-fat-fed mice, accompanied by inhibition of obesity-induced adipose inflammation. 17,19 Moreover, HO-1 induction has been shown to attenuate immobilization-induced skeletal muscle atrophy. 20 Given that inflammation is a key component of obesity-induced skeletal muscle atrophy, HO-1 induction may be a useful strategy for protecting against inflammation-related muscle loss/atrophy under obese conditions.
Quercetin (3,3,4,5,7-pentahydroxyflavone) is an abundant flavonoid present in various vegetables and fruits such as onions, tomatoes, and apples. 21 Quercetin elicits antioxidant and anti-inflammatory activities, and these properties are closely associated with its beneficial actions against various metabolic diseases such as insulin resistance, cardiovascular diseases, and cancer. 22 Interestingly, recent studies have demonstrated that quercetin protects against oxidative stress-induced muscle atrophy/damage both in vitro and in vivo. 23 –25 We previously reported that quercetin protects against obesity-induced inflammatory responses in adipose tissue and skeletal muscle by inhibiting inflammatory signaling and inflammatory mediator production, 3,26 accompanied by reduction of obesity-induced skeletal muscle atrophy. 3 However, the mechanisms by which quercetin protects against muscle atrophy under obese conditions, in terms of HO-1 induction, remains unclear. In this study, we demonstrated that quercetin protects against TNFα-induced atrophic responses in myotubes and obese skeletal muscle through Nrf2-mediated HO-1 induction.
Materials and Methods
Cell culture and treatment
The murine myoblast cell line C2C12 (5 × 105 cells/mL) was grown at 37°C in 5% CO2 in DMEM (Life Technologies, Grand Island, NY, USA) containing 10% FBS (Life Technologies) and 1% penicillin/streptomycin (Life Technologies). At a confluence of 95–100%, the medium was replaced with differentiation medium (DMEM plus 2% horse serum [Life Technologies]), which was changed after 2 days. Differentiated myotubes were treated for 2 days with 100 ng/mL of TNFα (PeproTech), quercetin (Jena Bioscience, Germany), or both drugs in combination.
Animal experiment
Six-week-old (C57BL/6, male) mice were purchased from Orient Ltd. (Orient, Busan, Korea). After a 1-week adaptation period, mice were randomly divided into three dietary groups (n = 6 per group). Mice were fed (1) a regular diet (13% of calories as fat from soybean oil; Harlan Teklad, Madison, WI, USA), (2) a high-fat diet (HFD) (60% of calories as fat from lard and soybean oil; Research Diets, New Brunswick, NJ, USA), or (3) an HFD supplemented with 0.05% (w/w) quercetin (Que) for 9 weeks. Animals were sacrificed by CO2 asphyxiation, and their muscles were dissected. Nrf2-deficient mice with a B6/JSL background were purchased from The Jackson Laboratory (Bar Harbor, ME, USA) and bred in a specific pathogen-free animal facility at the University of Ulsan. Male Nrf2-deficient mice and their wild-type (WT) littermates at 8 weeks of age were individually housed in plastic cages in a specific pathogen-free animal facility with a 12-h light:12-h dark cycle. Mice were fed an HFD (Research Diets) or an HFD supplemented with 0.05% (w/w) quercetin for 9 weeks and were given free access to food and water. All animal care and procedures were conducted according to the protocols and guidelines approved by the University of Ulsan Animal Care and Use Committee (LNY-14-010). Mice were maintained under specific pathogen-free conditions at 22°C ± 2°C with a 12-h light:12-h dark cycle.
Myotube diameter measurements
Microscopic images of myotubes were captured using a digital camera mounted on an Olympus CK2 microscope (Olympus Optical Co., Ltd., Japan). Myotube diameter from five random fields at 100 × magnification was measured using AxioVision AC software (Zeiss). Mean diameters were calculated as the means of three approximately equidistant measurements taken along the lengths of 15 myotubes.
DNA-binding activity of Nrf2 or NF-κB
Nuclear proteins were extracted using a nuclear extraction kit (Active Motif, Carlsbad, CA), and their concentrations were determined using a BCA Protein Assay kit (Pierce). Nuclear extracts were used to quantify DNA-binding activity of Nrf2, and the p65 subunit of NF-κB using ELISA-based TransAM kits (40096 and 50296; Active Motif) according to the manufacturer's instructions.
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from myotubes or muscle tissue samples with TRI reagent (Life Technologies, Carlsbad, CA, USA). Two microgram aliquots of total RNA were reverse-transcribed to cDNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA). Real-time polymerase chain reaction (PCR) amplification of cDNA was performed using a SYBR Premix Ex Taq kit (TaKaRa Bio, Inc., Forster, CA, USA) with a Thermal Cycler Dice (TaKaRa Bio, Inc., Japan). All reactions were performed according to the same schedule: 95°C for 10 sec, 45 cycles at 95°C for 5 sec, and 60°C for 30 sec. The results were analyzed using Real Time System TP800 software (TaKaRa Bio, Inc.), and all values were normalized to levels of the housekeeping gene. The primers used in the analysis are listed in Table 1.
MuRF1, muscle ring-finger protein 1.
Western blot analysis
Tissues and cells were homogenized in lysis buffer containing 150 mM NaCl, 50 mM Tris-HCl, 1 mM EDTA, 50 mM NaF, 10 mM Na4P2O7, 1% Igepal, 2 mM Na3VO4, 0.25% protease inhibitor cocktail, and 1% phosphatase inhibitor cocktail. Homogenates were centrifuged at 12,000 g for 20 min at 4°C. Samples containing 10–25 μg of total proteins were subjected to Western blot analysis using polyclonal antibodies specific for Nrf2 (Santa Cruz, CA, USA), HO-1 (Enzo Live Science, Inc.), MuRF1 (Santa Cruz), atrogin-1 (Santa Cruz), IκB-α (Santa Cruz), lamin A/C (Santa Cruz), and α-tubulin (Abcam). Protein bands were detected using an enhanced chemiluminescence Western blotting detection kit (Perkin Elmer, Waltham, MA). Protein intensities were quantified by densitometry using ImageJ software.
Statistical analysis
The results are presented as mean ± standard error of the mean of values obtained from repeated experiments. Statistical analysis was performed using Student's t-test or one-way analysis of variance (ANOVA) followed by Tukey's test with Prism 5.0 software (Graphic Pad, San Diego, CA, USA). Differences were considered to be significant at P < .05.
Results
Effects of quercetin on TNFα-induced atrophic responses in myotubes
TNFα production is elevated in obesity-induced inflammatory states and is implicated as a major mediator of muscle atrophy. 27 Here, we first examined the effect of quercetin on TNFα-induced muscle atrophy in C2C12 myotubes by analyzing changes in the morphological appearance of TNFα-treated C2C12 myotubes. TNFα treatment significantly reduced myotube diameter (Fig. 1A), whereas quercetin completely restored TNFα-induced reduction of myotube diameter (Fig. 1A). Subsequently, we examined whether or not quercetin alters the expression of MAFbx/atrogin-1 and MuRF1, which are markers of muscle atrophy. For this, myotubes were treated with TNFα, and atrophic markers were determined by real-time PCR and/or Western blot analyses. Quercetin significantly downregulated transcription levels of MAFbx and MuRF1 (Fig. 1B) as well as reduced protein expression levels of MAFbx and MuRF1 in TNFα-treated C2C12 myotubes (Fig. 1C). These results indicate that quercetin reduces TNFα-induced muscle atrophy/loss under obese conditions.
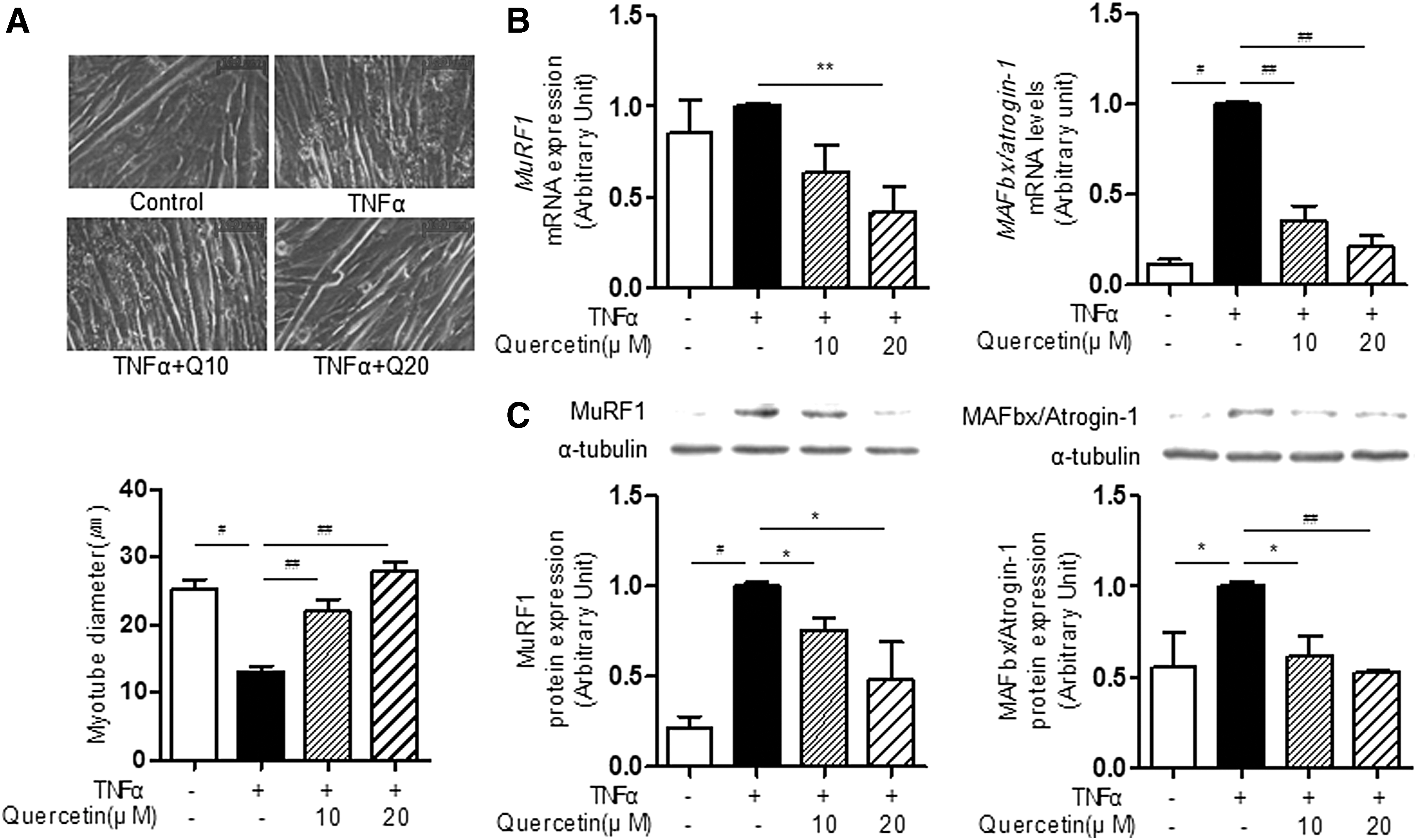
Effect of quercetin on TNFα-induced atrophic response in myotubes. Myotubes differentiated for 2 days were treated with 100 ng/mL of TNFα in the presence or absence of quercetin for 48 h.
Effects of quercetin on Nrf2 activation and HO-1 induction in myotubes
HO-1 is a major component in the defense against oxidative stress and inflammatory responses and is regulated by activation of Nrf2, a basic leucine zipper transcription factor. It has been shown that HO-1 can be triggered by certain electrophilic compounds such as polyphenols and plant-derived components. 28 In this study, we first examined the effect of quercetin on HO-1 induction in myotubes and found that quercetin treatment to myotubes significantly upregulated expression levels of HO-1 protein in a dose-dependent manner (Fig. 2A). Along with this, we observed that quercetin increased Nrf2 protein levels in the nuclear fraction of myotubes (Fig. 2B), indicating that quercetin may have the potential to induce HO-1 through Nrf2 activation.
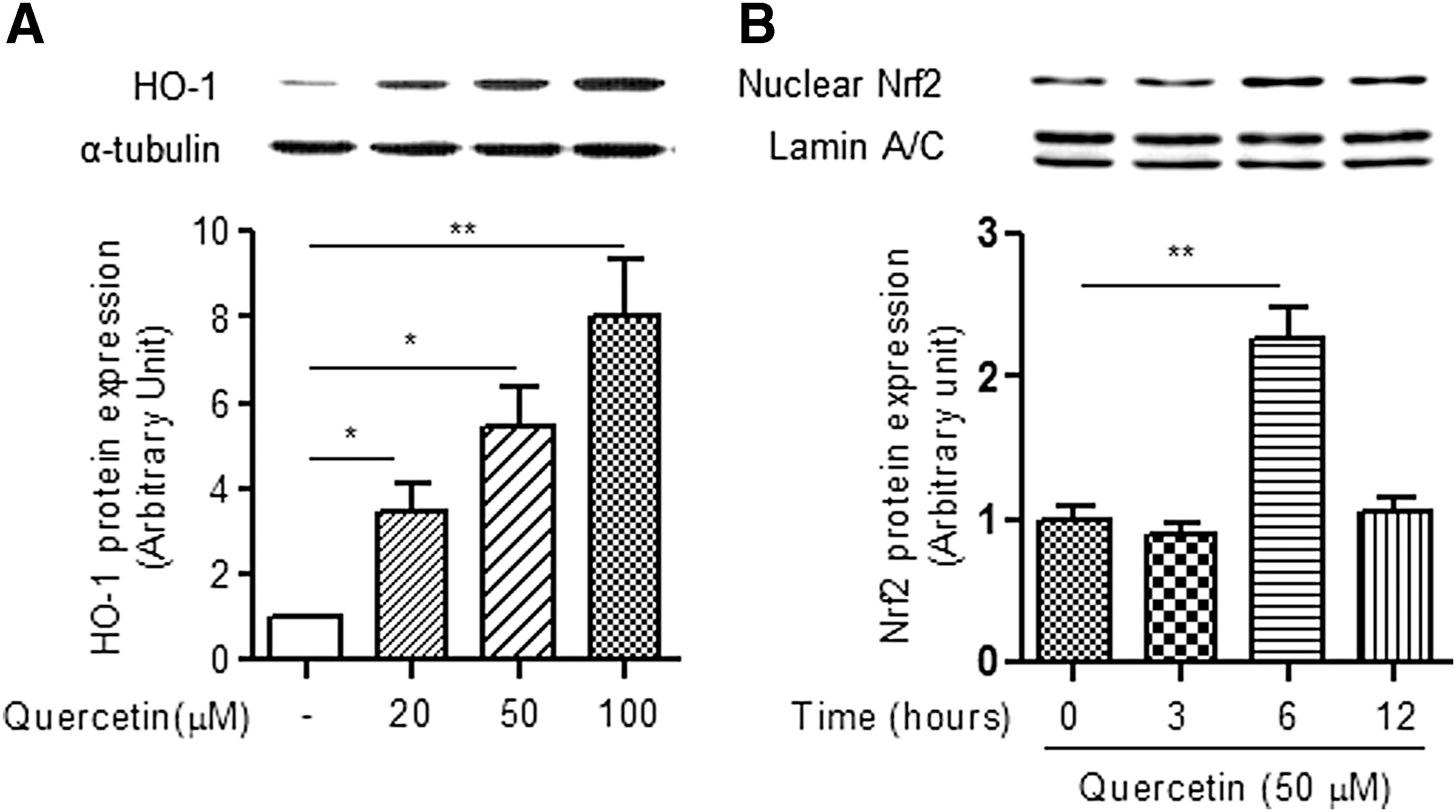
Effect of quercetin on HO-1 and Nrf2 expression in myotubes. Differentiated myotubes (4 d) were exposed to the indicated concentrations (0, 20, 50, 100 μM) of quercetin or for the indicated times (0, 3, 6, 12 h).
Effects of HO-1 inhibitor on the beneficial actions of quercetin
Using the HO-1 inhibitor protoporphyrin IX zinc (II) (ZnPP), we further examined the direct association of HO-1 induction with the beneficial actions of quercetin against TNFα-induced atrophic factors. We confirmed that quercetin suppressed IκB-α degradation, which regulates genes encoding proinflammatory cytokines such as TNFα, suggesting a decrease in NF-κB activity in cells. Moreover, the HO-1 inhibitor ZnPP abrogated the inhibitory effect of quercetin on TNFα-stimulated IκB-α degradation (Fig. 3A). Along with this, we observed that quercetin abrogated expression of the TNFα-induced atrophic factor MuRF1 in myotubes as well as restored reduction of myotube diameter (Fig. 3B, 3C). More importantly, the HO-1 inhibitor ZnPP significantly inhibited the protective actions of quercetin against TNFα-induced reduction of MuRF1 expression and myotube diameter (Fig. 3B, C).
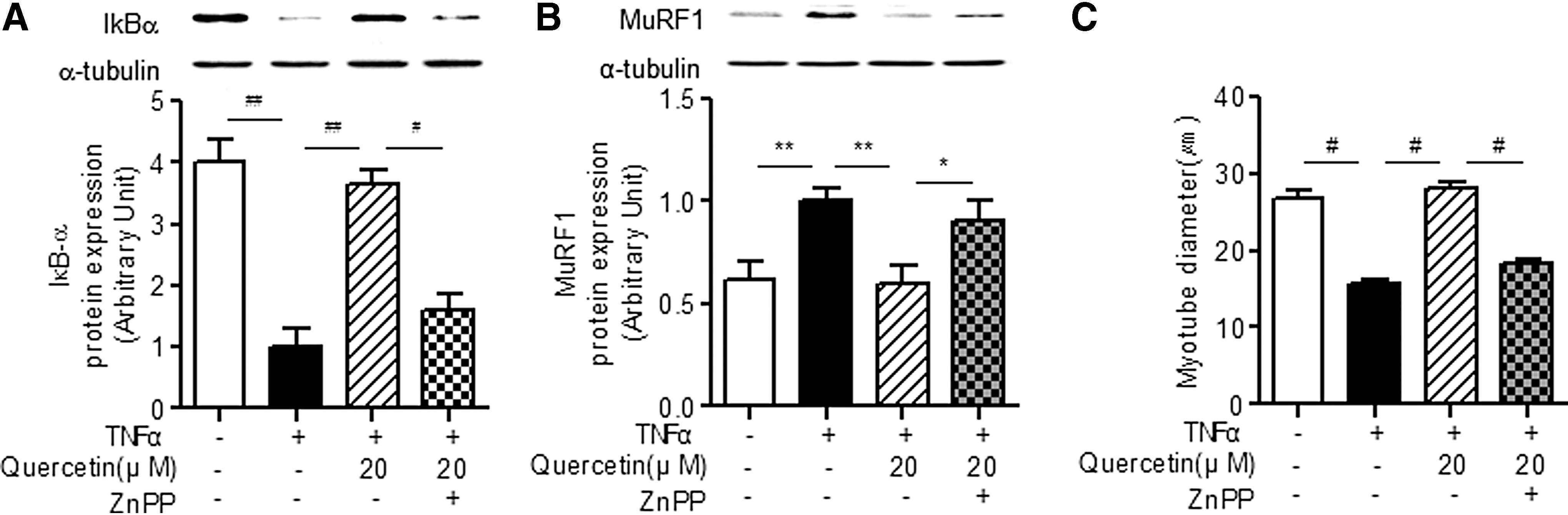
Effect of HO-1 inhibitor on quercetin-mediated antiatrophic action in TNFα-treated myotubes. Differentiated myotubes were treated with 100 ng/mL of TNFα plus 20 μM quercetin and/or 0.1 μM ZnPP for 30 min.
Effects of quercetin supplementation on HO-1 induction and NF-κB activation in skeletal muscle of obese mice fed an HFD
Next, we examined whether or not quercetin supplementation promotes HO-1 induction in skeletal muscle of obese mice fed an HFD in vivo as well as whether or not the effect is associated with inhibition of NF-κB activation. As shown in Figure 4A, HO-1 protein expression was significantly upregulated in skeletal muscle of quercetin-supplemented obese mice compared with that of control obese mice. Subsequently, we observed that quercetin supplementation enhanced Nrf2 activation and suppressed NF-κB activation in skeletal muscle of obese mice fed an HFD (Fig. 4B, C). At the same time, quercetin supplementation downregulated MuRF1 expression in skeletal muscle of obese mice fed an HFD (Fig. 4D).
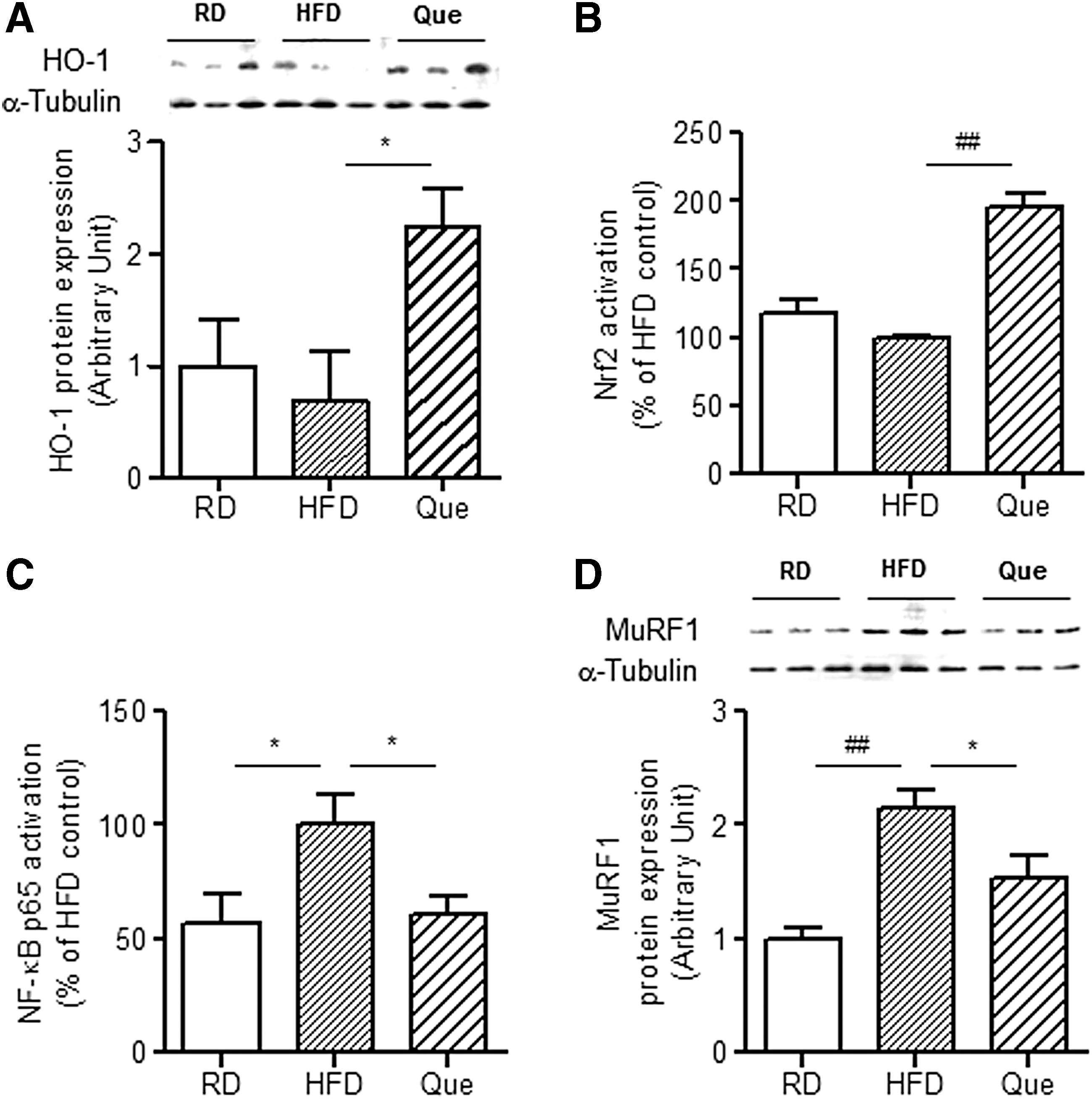
Effect of quercetin supplementation on HO-1 induction and NF-kB activation in skeletal muscle of obese mice fed an HFD. C57BL/6 mice were fed an RD, an HFD, or an HFD supplemented with 0.05% quercetin (Que) for 9 weeks.
Effects of Nrf2 deficiency on quercetin actions in skeletal muscle of obese mice fed an HFD
Using Nrf2-deficient obese mice fed an HFD, we further examined whether or not HO-1 induction by quercetin is mediated via the Nrf2 pathway. As shown in Figure 5A, quercetin-induced upregulation of HO-1 protein expression was inhibited in the skeletal muscle of Nrf-2-deficient obese mice compared with that of the WT obese control, and no Nrf2 activation by quercetin was observed in the Nrf2-deficient obese mice (Fig. 5B). Furthermore, quercetin-mediated NF-κB inactivation was significantly reduced in the skeletal muscle of Nrf2-deficient obese mice (Fig. 5C). Moreover, we confirmed that the protective action of quercetin against MuRF1 expression was diminished in Nrf2-deficient obese mice (Fig. 5D).
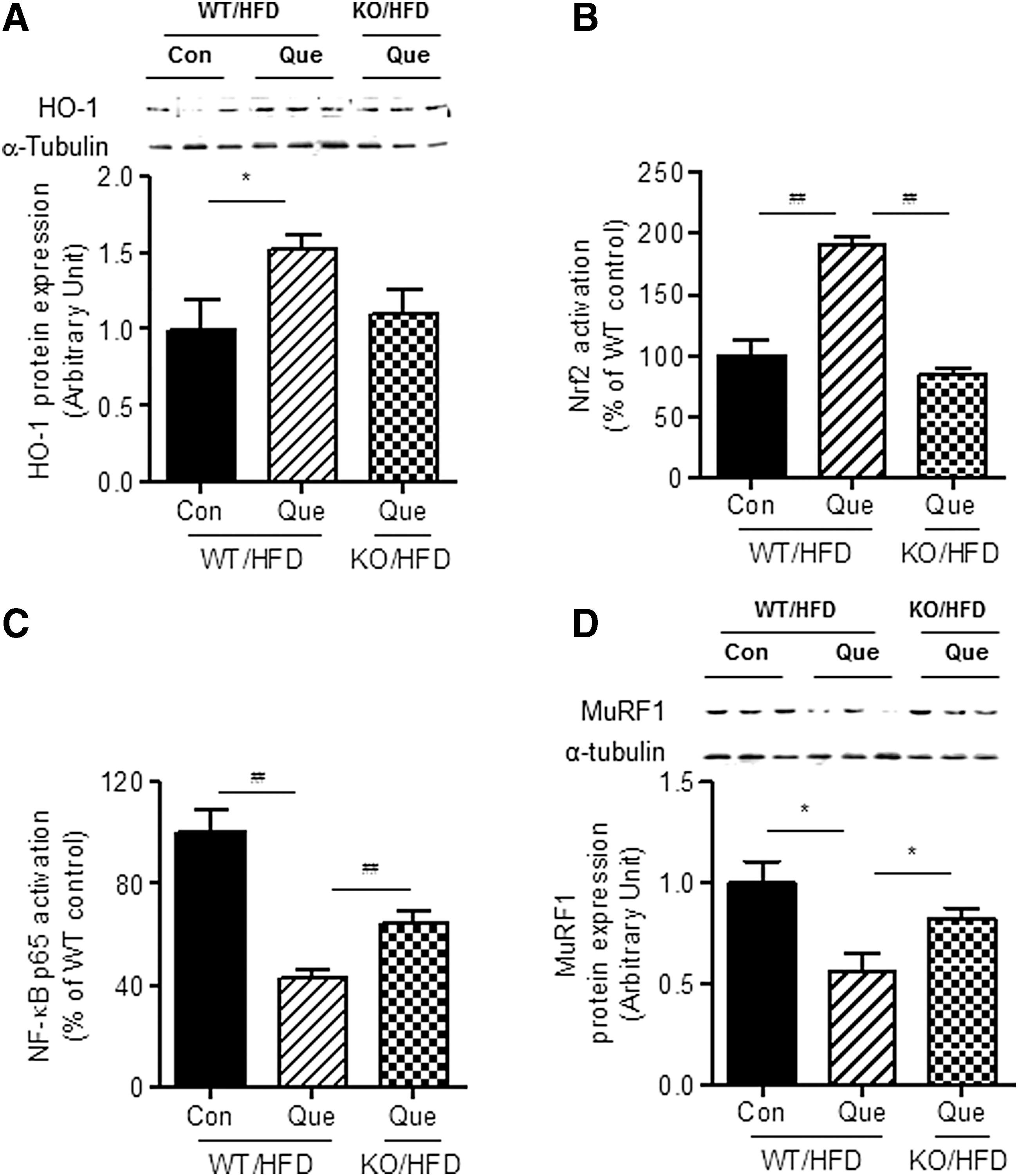
Effect of Nrf2 deficiency on quercetin action in skeletal muscle of obese mice fed an HFD. WT and Nrf2-deficient mice were fed an HFD or an HFD supplemented with 0.05% quercetin for 9 weeks.
Discussion
TNFα is a potent activator of NF-κB, which is one of the most important inflammatory signaling pathways, and it is closely linked to loss of skeletal muscle mass in various inflammatory conditions such as obesity. 27,29 Activation of NF-κB in skeletal muscle enhances degradation of muscle-specific ubiquitin ligases such as MuRF1 and MAFbx/atrogin-1 as well as inhibits regeneration of myofibers after injury/atrophy. 29 These reports strongly suggest that TNFα and/or NF-κB could be important targets for protection against muscle atrophy. Indeed, it has been shown that the anti-TNFα agent infliximab, which suppresses NF-κB activation, reverses inflammatory sarcopenia in patients with Crohn's disease. 30 In this study, quercetin inhibited TNFα-induced upregulation of MuRF1 and MAFbx/atrogin-1 at both the transcript and protein levels in myotubes, accompanied by suppression of TNFα-induced IκB-α degradation, implying inactivation of NF-κB. The antiatrophic action of quercetin was also confirmed in skeletal muscle of HFD-fed obese mice. Taken together, this indicates that quercetin may elicit its antiatrophic activity under obese conditions by inhibiting TNFα-induced inflammatory signaling.
It has been reported that NF-κB activation acts as a repressor of Nrf2 transcriptional activity, 31 which regulates expression of HO-1 and antioxidant genes that protect cells from various inflammatory injuries. 32 Conversely, the Nrf2/HO-1 pathway inhibits NF-κB-mediated transcription by preventing degradation of IκB-α, 31 suggesting that factors modulating Nrf2 activation and/or HO-1 induction have the potential to protect against TNFα-mediated muscle atrophy. Indeed, the HO-1 inducer hemin has been shown to reduce immobilization-induced skeletal muscle atrophy as well as suppress protein degradation. 20 In this study, we observed that quercetin per se markedly upregulated expression of HO-1 protein in myotubes, accompanied by Nrf2 activation. Along with this, induction of HO-1 by quercetin supplementation was associated with inhibition of both NF-κB activation and MuRF1 protein expression in HFD-fed obese mice. Together, these results indicate that quercetin by itself is a potent inducer of Nrf2-mediated HO-1 induction in myotubes, leading to suppression of NF-κB activation.
Of note, ZnPP, a specific inhibitor of HO-1 activity, significantly suppressed the ability of quercetin to inhibit TNFα-induced IκB-α degradation in addition to expression of the ubiquitin ligase MuRF1 as well as myotube diameter. The antiatrophic action of quercetin in TNFα-treated myotubes, accompanied by HO-1 induction and NF-κB inactivation, was confirmed in skeletal muscle of obese mice fed an HFD. These findings suggest that activation of the Nrf2/HO-1 pathway by quercetin may inhibit NF-κB activation, leading to reduction of the TNFα-induced atrophic response in myotubes and skeletal muscle of obese mice. To determine whether or not quercetin-induced HO-1 induction depends on the Nrf2 pathway, we examined whether or not Nrf2 deficiency abolishes the action of quercetin on HO-1 induction and/or the atrophic response in obese mice fed an HFD. We observed that the antiatrophic effect of quercetin supplementation was partially reduced in Nrf2-deficient HFD-fed obese mice compared with WT obese control mice. Moreover, inactivation of NF-κB and induction of HO-1 by quercetin were decreased in Nrf2-defieicnt obese mice. These results indicate that the Nrf2 pathway may be partially associated with quercetin-mediated HO-1 induction and antiatrophic activity, and there may be another mechanism responsible for HO-1 induction by quercetin.
It has been previously reported that peroxisome proliferator-activated receptors (PPARs) are important modulators of HO-1 activity. 33 It is likely that HO-1 is a direct PPAR target gene, whose transcription is regulated by PPARγ and/or PPARα via PPAR response elements. 33 Ligands for PPARγ or PPARα are known to elicit anti-inflammatory and antiatrophic effects, 34,35 indicating that PPAR ligands may have the potential to upregulate HO-1. Indeed, several plant-derived polyphenols such as curcumin and resveratrol, which elicit PPAR ligand activities, have been shown to upregulate HO-1 expression 28,36,37 and protect against muscle damage. 38 Moreover, activation of PPARs by curcumin or resveratrol is mediated through the NF-κB pathway. 39 Given that quercetin has PPARγ and/or PPARα ligand activity, 40,41 induction of HO-1 by quercetin may be at least partially associated with activation of PPARs.
In summary, quercetin reduces atrophic responses in myotubes and skeletal muscle under obese conditions, accompanied by induction of HO-1 and inactivation of NF-κB. The protective effect of quercetin was abrogated by an HO-1 inhibitor in TNFα-treated myotubes and Nrf2 deficiency in skeletal muscle of HFD-fed obese mice. These findings suggest that the antiatrophic effects of quercetin may be mediated via Nrf2/HO-1 pathway accompanied by inactivation of NF-κB (Fig. 6). Quercetin may be used as a dietary factor to protect against obesity-induced skeletal muscle atrophy.

Schematic presentation of the beneficial effect of quercetin on TNFα-induced muscle atrophy via induction of HO-1. Quercetin reduces levels of MAFbx/atrogin-1 and MuRF1 and thus protects against TNFα-induced muscle atrophy under obese conditions. The antiatrophic action of quercetin is mediated through Nrf2-mediated HO-1 induction accompanied by inactivation of NF-κB.
Footnotes
Acknowledgments
This work was supported by a grant from the Korea Food Research Institute (KFRI E0160500-02) and the Priority Research Centers Program (2014R1A6A1030318) of the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology.
Author Disclosure Statement
No competing financial interests exist.