
Abstract
The high propensity of influenza viruses to develop resistance to antiviral drugs necessitates the continuing search for new therapeutics. Peanut skins, which are low-value byproducts of the peanut industry, are known to contain high levels of polyphenols. In this study, we investigated the antiviral activity of ethanol extracts of peanut skins against various influenza viruses using cell-based assays. Extracts with a higher polyphenol content exhibited higher antiviral activities, suggesting that the active components are the polyphenols. An extract prepared from roasted peanut skins effectively inhibited the replication of influenza virus A/WSN/33 with a half maximal inhibitory concentration of 1.3 μg/mL. Plaque assay results suggested that the extract inhibits the early replication stages of the influenza virus. It demonstrated activity against both influenza type A and type B viruses. Notably, the extract exhibited a potent activity against a clinical isolate of the 2009 H1N1 pandemic, which had reduced sensitivity to oseltamivir. Moreover, a combination of peanut skin extract with the anti-influenza drugs, oseltamivir and amantadine, synergistically increased their antiviral activity. These data demonstrate the potential application of peanut skin extract in the development of new therapeutic options for influenza management.
Introduction
I
Because of their diversity, natural products have gained interest as potential sources of antivirals. Antiviral activities of naturally occurring nutrients may be of special interest not only because of their availability but also due to the possibility of their incorporation as part of a diet to combat viral infections. To date, some extracts and food constituents have demonstrated antiviral effects: Vigna angularis (red bean) extract against rabies virus, 4 Litchi chinensis (Lychee fruit) extract against betanodavirus, 5 and whey acidic proteins against human immunodeficiency virus (HIV). 6 In addition, extracts from Alchemilla mollis (Lady's mantle tea), 7 Aspalathus linearis (Rooibos tea), 8 Panax ginseng (Red ginseng), 9 purified constituents such as methylglyoxal from manuka honey, 10,11 lactoferrin from bovine milk, 12 catechins from green tea, 13 and a dihydrochalcone from seagrass Thalassodendron ciliatum (Forsk.) den Hartog 14 have been shown to possess potent anti-influenza virus activity. These studies highlight the possibility of using crude extracts and purified compounds from natural sources, especially foods, to control viral infections.
The peanut, Arachis hypogaea L., is a globally important commercial crop used for the production of oil and as an important source of protein. It recently gained attention as a functional food. 15 The consumption of peanuts is associated with many health-promoting attributes, including protection against heart disease, 16 body weight management, 17 anticancer, 18 and antidiabetes properties. 19 These health benefits are attributed to the trans-fatty acids, minerals, vitamins, fibers, and bioactive compounds present in the peanut. 20 The peanut skin, a byproduct of the peanut industry, contains high levels of polyphenols, which exert potent antioxidant, 21 anti-inflammatory, 22 hypolipidemic, 23 and antibacterial 24 effects. Despite these studies indicating that peanut skins are a potentially rich, safe, and inexpensive source of functional ingredients that could be helpful for combating microbial infections, their antiviral activity remains to be studied. In this study, we aimed to investigate the anti-influenza activity of peanut skins. We found that crude ethanol extracts of peanut skins possess potent inhibitory activity against influenza type A and B viruses. Moreover, the extract showed a synergistic effect when combined with approved anti-influenza drugs, oseltamivir and amantadine. Our work accentuates the biological importance of peanut skins and highlights their possible application in the management of viral infections.
Materials and Methods
Cells, viruses, chemicals, and peanut skin
Madin Darby Canine Kidney (MDCK) cells, a kind gift from Dr. Kyosuke Nagata (Tsukuba University, Japan), were maintained in minimum essential medium (MEM) purchased from Wako Pure Chemical Industries, Ltd. (Tokyo, Japan) and supplemented with 5% fetal bovine serum from Life Technologies (Scoresby, Australia), 100 U/mL penicillin, and 100 μg/mL streptomycin (Nacalai Tesque, Inc., Kyoto, Japan) at 37°C in 5% CO2. Influenza viruses A/WSN/33 (H1N1), A/Puerto Rico/8/34 (H1N1), A/Virginia/ATCC2/2009 (H1N1), A/Aichi/2/68 (H3N2), and B/Lee/40 were prepared as described. 25 Oseltamivir phosphate (F. Hoffmann-La Roche, Basel, Switzerland) and Zanamivir (LTK Laboratories, St. Paul, MN, USA) were dissolved in phosphate-buffered saline (PBS) and dimethyl sulfoxide (DMSO), respectively. Amantadine (Sigma-Aldrich, Tokyo, Japan) was dissolved in water and gallic acid (Nacalai Tesque) in ethanol. Resveratrol (Merck Millipore, Billerica, MA, USA) was dissolved in DMSO. Peanut skin samples were obtained as previously described 26 and are summarized in Table 1.
N/A, not available.
Preparation of peanut skin extracts
Peanut skins were pulverized into a fine powder using a blender. Ten grams of the powder were defatted with 50 mL of hexane for 1 h, the solvent was discarded and the process was repeated twice. The same procedure was repeated using ethyl acetate. Finally, the residue was extracted overnight with 100 mL of ethanol. The solvent was then evaporated to obtain dry material. The defatting and extraction processes were performed at 30°C under continuous shaking. The dried material was reconstituted to 5 mg/mL in ethanol and used for the determination of total polyphenol and resveratrol content, antiviral activity, and cytotoxicity.
Determination of polyphenol and resveratrol contents
The Folin–Ciocalteu assay was used to determine the polyphenol content of the extracts as described previously. 27 In brief, 20 μL of peanut skin extract or gallic acid serial dilutions were added to a 96-well microtiter plate, followed by the addition of 100 μL Folin–Ciocalteu reagent (Nacalai Tesque). The mixture was incubated at room temperature (RT) for 5 min, and 80 μL of 7.5% sodium carbonate was added. Controls of reagent blank and sample dilutions without reagents were included. After a 2 h incubation at RT, optical density was measured at 760 nm with an Infinite M200 plate reader (TECAN, Mannedorf, Switzerland). A calibration curve was prepared using gallic acid, and it was used for the calculation of the polyphenol content in the samples as a gallic acid equivalent. The resveratrol content of the peanut skin extracts were quantified using the liquid chromatograph–tandem mass spectrometry (LC/MS/MS) method by the Japan Food Research Laboratories (Tokyo, Japan). 28 In brief, samples (10 mL) were injected onto a HPLC system equipped with an Inertsil ODS-2 column (5 μm, 150 × 2.1 mm). Separation was performed using a mobile phase composed of AcOH in H2O and ACN-MeOH at 35°C. The typical flow rate was maintained at 0.2 mL/min. Mass spectrometry analysis was performed using electro spray ionization and negative detection mode. Scan range was from m/z 142.8 to 226.9. The amount of resveratrol was determined from an average of two independent measurements.
Cellular toxicity and antiviral activity assay
Cellular toxicity and antiviral activity were evaluated as described. 29 In brief, MDCK cells (3 × 104 per well) seeded in 96-well tissue culture plates were treated with serial dilutions of extracts in MEM vitamin (MEM supplemented with 1% of 100 × MEM vitamin). Influenza virus solution of 100 TCID50 (50% tissue culture infective dose) was added per well for the antiviral activity assay. After 48 h incubation, cell density was determined by crystal violet (CV) staining. The percentage of relative cell density was calculated by comparing the optical density of the treated wells to those of the untreated controls. The 50% cytotoxic concentration (CC50) and 50% inhibitory concentration (IC50) were calculated by linear regression analysis using Microsoft Excel software. The selectivity index was determined from the CC50 to IC50 ratio.
Virus titration
Virus titer in the culture supernatant of cells infected with the virus in the presence of extract was determined using a TCID50 assay as described. 30 In brief, 150 μL of 100-fold dilution of culture supernatant were transferred to 96-well microtiter plates and serially diluted 10-fold in MEM vitamin. A hundred microliters of the dilutions was added to MDCK cells seeded in 96-well tissue culture plates. After incubation for 72 h, cell density was determined by CV staining and TCID50 per virus dilution calculated using Reed and Muench method.
Plaque formation assay
The plaque formation assay was performed as described with some modifications. 10 In brief, MDCK cell monolayers in six-well tissue culture plates were washed with serum-free medium and infected with 300 plaque-forming units of the A/WSN/33 virus at 37°C in 5% CO2 for 1 h. After washing with MEM vitamin, cells were overlaid with MEM containing 0.8% agarose, 0.1% bovine serum albumin, 1% 100 × MEM vitamin solution, and 0.03% glutamine. After incubation at 37°C for 72 h, the plaques were visualized by fixing the cells with acetic acid-ethanol (1:1) and staining with 0.5% amido black 10B. Plaques were counted by visual examination and the results represented as a ratio of plaque number in the presence of extract or zanamivir to that of untreated controls.
Combination of peanut skin extract and anti-influenza drugs
An analysis of the combination of peanut skin extract with anti-influenza drugs was performed as previously described. 7 In brief, MDCK cells in 96-well tissue culture plates were infected with 100 TCID50 of A/WSN/33 virus per well in the presence of peanut skin extract (0.08, 0.31, and 1.25 μg/mL) mixed with twofold serial dilutions of either oseltamivir (0.008–2 μg/mL) or amantadine (0.37–47 μg/mL). After 48 h incubation at 37°C, cell density was determined by CV staining, and the percentage of inhibitory activity was calculated relative to the cell density in untreated controls. The effect of the combination treatment was analyzed by calculating the combination index at 50% inhibitory activity using the method described by Chou and Talay, 31 in which drug combinations are assessed using combination index {for drugs A & B (CI) = Ac/Ae+Bc/Be}, where Ac and Bc are the concentrations of A and B when used in combination, and Ae and Be are the concentrations able to produce an effect of the same magnitude if used alone. If CI is <1, the effect of the combination is synergistic, whereas if CI = or >1, the effect is additive or antagonistic, respectively.
Statistical analysis
The results in graphs and tables represented as the mean ± standard deviation were calculated from three independent experiments performed in duplicate. To test statistical significance, each experiment was performed in three replicates and repeated three times. The difference between test samples and untreated controls was evaluated using the Student's t-test. A P value of <.01 was considered statistically significant.
Results
Polyphenol and resveratrol content, antiviral activity, and cellular toxicity of peanut skin extracts
Peanut skin samples used in this study were produced in different regions of the world and their skins were prepared by roasting, blanching, or pickling (Table 1). Ethanol extracts of the skin samples were prepared and evaluated for total polyphenol content, resveratrol content, antiviral activity, and cellular toxicity. 26 The dry weight of ethanol extract per 10 g of raw material and the percentage of total polyphenol content are indicated in Table 2. Apart from sample H, all extracts had a polyphenol content of 31–34%. Unlike the skins of the other samples, which were removed by roasting or blanching, the skin of sample H was removed by pickling. In addition, the amount of dry material from sample H was considerably lower than that from the rest of the samples, possibly due to the loss of phytochemicals during sample processing.
Extract obtained from 10 g peanuts skins.
Polyphenol content was determined by Folin–Ciocalteu method as gallic acid equivalent.
50% IC against A/WSN/33 virus.
50% CC for MDCK cells.
SI, the ratio of CC50 to IC50
CC, cytotoxic concentration; IC, inhibitory concentration; MDCK, Madin Darby Canine Kidney; N/A, not applicable; ND, not determined; SI, selective index.
Analysis of anti-influenza activity showed that all samples could inhibit virus replication with IC50 values of 1.0–1.5 μg/mL, except for sample H, which presented an IC50 value of 13.9 μg/mL. In addition, the range of cellular toxicities of the samples was close, with CC50 values of 5.4–9.1 μg/mL, while that of sample H was lower (>50 μg/mL). These data depicted a correlation between polyphenol content, antiviral activity, and cellular toxicity, suggesting that polyphenols are the major active components in the peanut skin extracts. As a control, we used commercially prepared peanut skin extract powder (sample I). Sample I had polyphenol contents, antiviral activity, and cytotoxicity similar to those of the other samples. In addition, resveratrol, one of the polyphenols in peanut skin, which has been reported to have anti-influenza activity, 32 was tested. Resveratrol exhibited anti-influenza virus activity with an IC50 value of 5.0 μg/mL. Quantification of resveratrol revealed that the extracts had varying amounts which did not correlate with polyphenol content, antiviral activity, or cellular toxicity implying that other polyphenols were also responsible for the activities observed (Table 2). Since sample A is well characterized and IC50 was not very different from other samples, it was selected for further analyses.
Peanut skin extract suppress viral cytopathic effects and virus production
We have established the CV assay, 29 as a rapid method for evaluation of antiviral activity based on cytopathic effect (CPE), where infected cells detach from the bottom of the tissue culture plates. This assay was used to investigate the inhibitory effects of peanut skin extract (sample A) and oseltamivir in low and high infection titer. The experimental condition of low infection titer (0.003 TCID50/cell for 48 h incubation, Fig. 1A) allows multiple cycles of virus replication and mimics the spread of virus during the incubation period in vivo, whereas high infection titer (1 TCID50/cell for 24 h treatment, Fig. 1B) reflects single cycle of replication in infected cells. In Figure 1A, oseltamivir showed good antiviral activity as expected, since oseltamivir inhibits the release of virus particles, thereby suppressing the spread of infection. Sensitivity of oseltamivir drastically decreased when the virus was infected with high titer (Fig. 1B). In contrast, CPE was suppressed by peanut skin extract concentrations of 1.56 and 3.13 μg/mL in both low and high infection titer (Fig. 1A, B). The virus titer in the supernatant was markedly reduced (<1 × 103 TCID50/mL) by 3.13 μg/mL of peanut skin extract treatment in both low and high infection titer, and 41.0 μg/mL oseltamivir treatment at low infection titer (Fig. 1C, D).
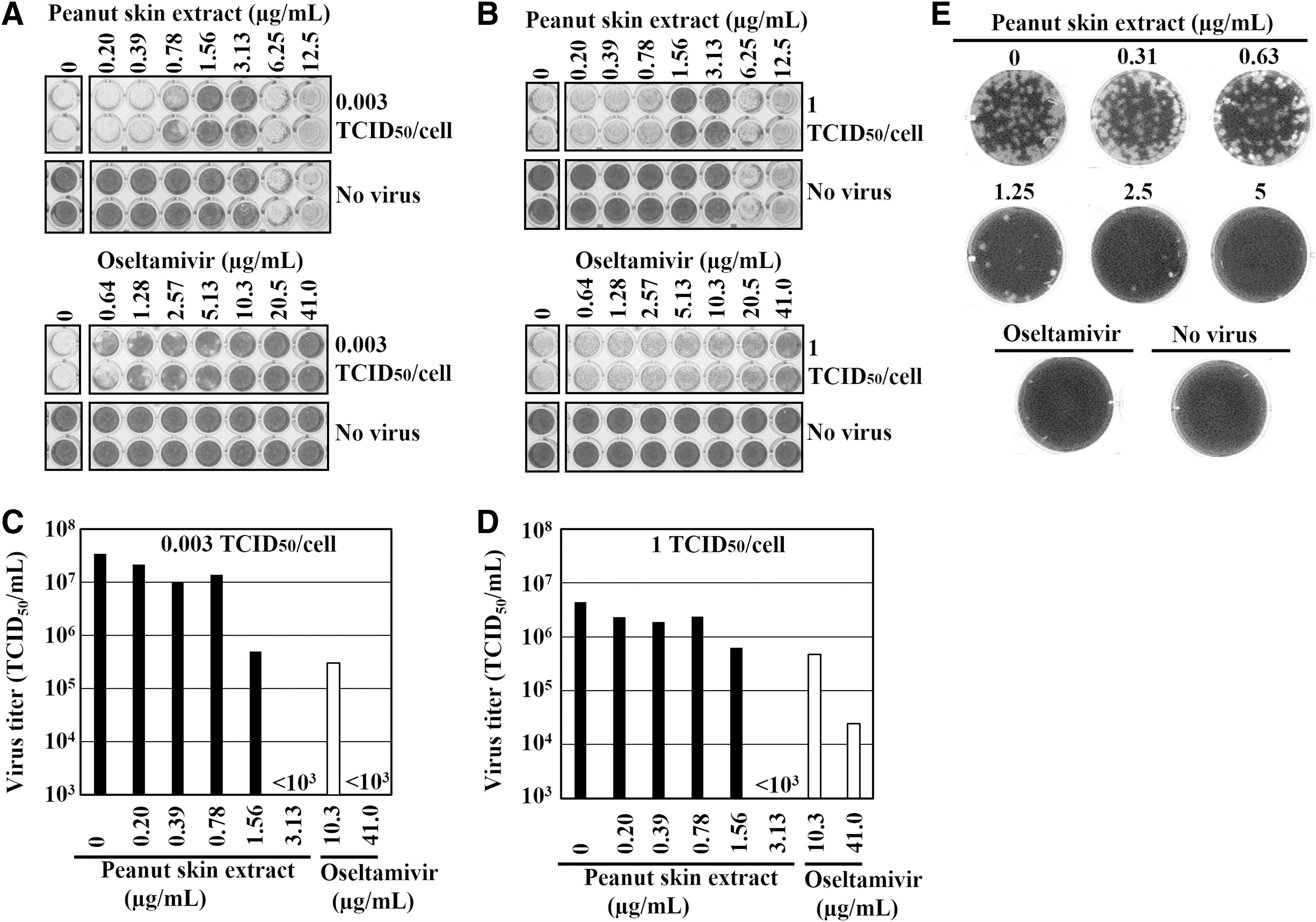
Next, plaque inhibition assay, 33 a gold standard for the evaluation of antiviral activity was performed (Fig. 1E). The number of plaques were dose-dependently decreased with concentrations higher than 1.25 μg/mL of peanut skin extract, which is in good agreement with IC50 value obtained by CV assay (Table 1, sample A). These results demonstrate that peanut skin extract suppresses CPE and virus production, and that CV assay with low infection titer (0.003 TCID50/cell) can be used for evaluation of antiviral activity instead of plaque inhibition assay.
Inhibitory effects of peanut skin extract on different stages of viral replication
We investigated the stage of influenza virus replication inhibited by peanut skin extract by performing different virus and/or cell treatments (Fig. 2). Treatment of cells with extract before virus infection did not suppress plaque formation when compared with those of the untreated control. In addition, neither incubation of the virus with extract before infection nor treatment of the cells after virus infection inhibited plaque formation. However, simultaneous addition of virus and extract to cells significantly reduced plaque formation, suggesting that the extract inhibits the early stages of virus infection. On the contrary, zanamivir completely suppressed plaque formation when added to the cells after virus infection. These results indicate that peanut skin extract targets a different stage of viral replication from that inhibited by zanamivir.
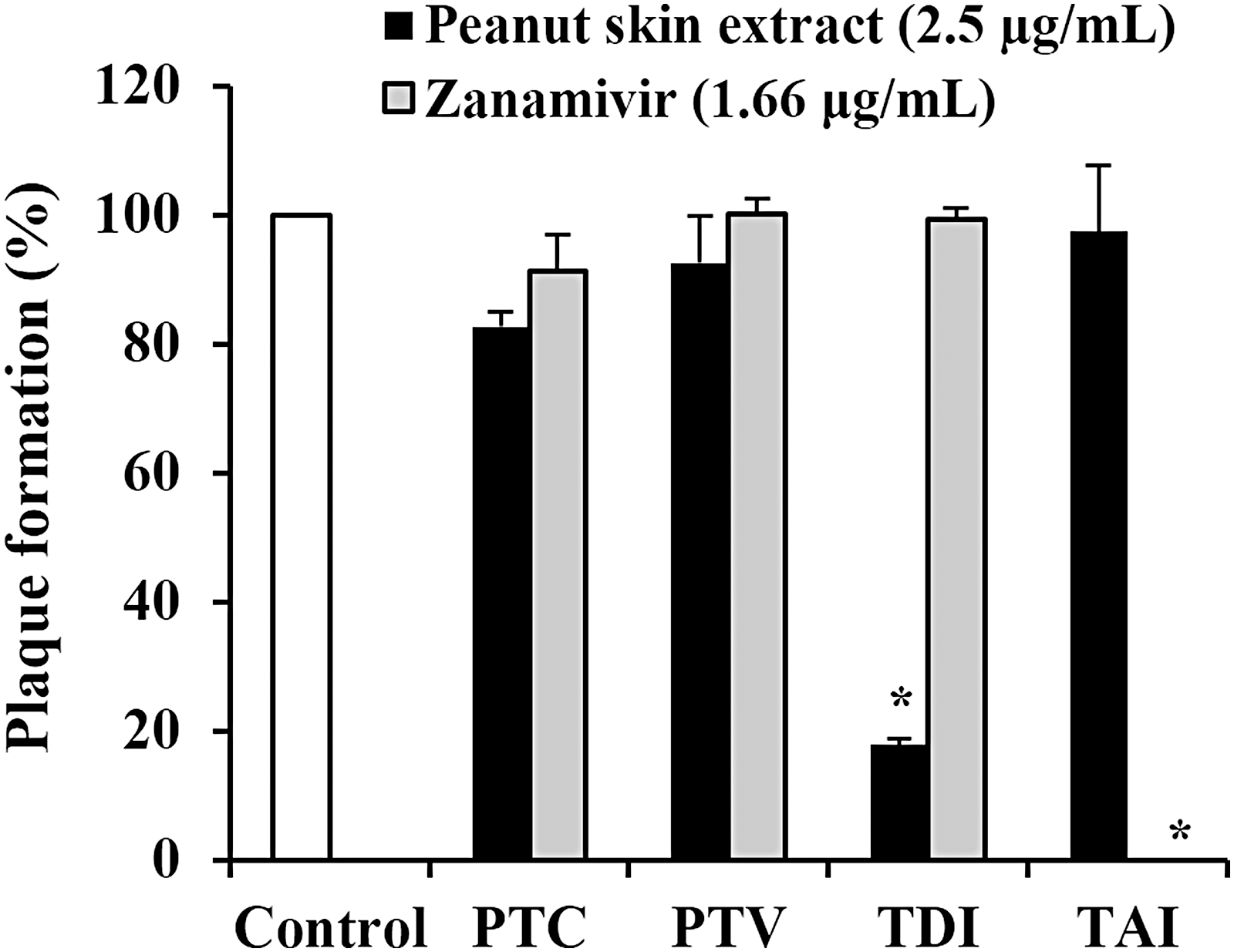
Effect of peanut skin extract on plaque formation. Plaque formation assays were carried out as described in Materials and Methods section with the following treatment protocols for the MDCK cells and A/WSN/33 virus; Pretreatment of cells (PTC): cells were treated with the sample and incubated at 37°C for 1 h. After removing the sample, cells were washed and infected with virus. Pretreatment of virus (PTV): virus was mixed with sample and incubated at RT for 1 h. The pretreated virus was then used to infect cells. Treatment during infection (TDI): the sample and virus were simultaneously added to cells and incubated at 37°C for 1 h. Treatment after infection (TAI): cells were infected with virus and then overlaid with agarose containing the sample. The results are presented as the percentage of plaques formed in each treatment relative to the plaques formed in untreated controls. The results are represented as the mean ± SD obtained from three independent experiments performed in triplicate. Asterisk indicates statistical significance (P < .01, Student's t-test). RT, room temperature; SD, standard deviation.
Combination of peanut skin extract and antiviral drug
Influenza virus infected cells were cotreated with peanut skin extract and antiviral drugs, oseltamivir and amantadine. As demonstrated in Figure 3, the presence of the extract improved the inhibitory activities of both drugs. In Table 3, the IC50 of oseltamivir and amantadine with 1.25 μg/mL of peanut skin extract treatment decreased 23.5-fold and 38.7-fold, respectively. The combination indices of both oseltamivir and amantadine with various concentrations of peanut skin extract was <1 indicating synergistic effects. These results imply that peanut skin extract could enhance the antiviral activity of the currently approved anti-influenza drugs.
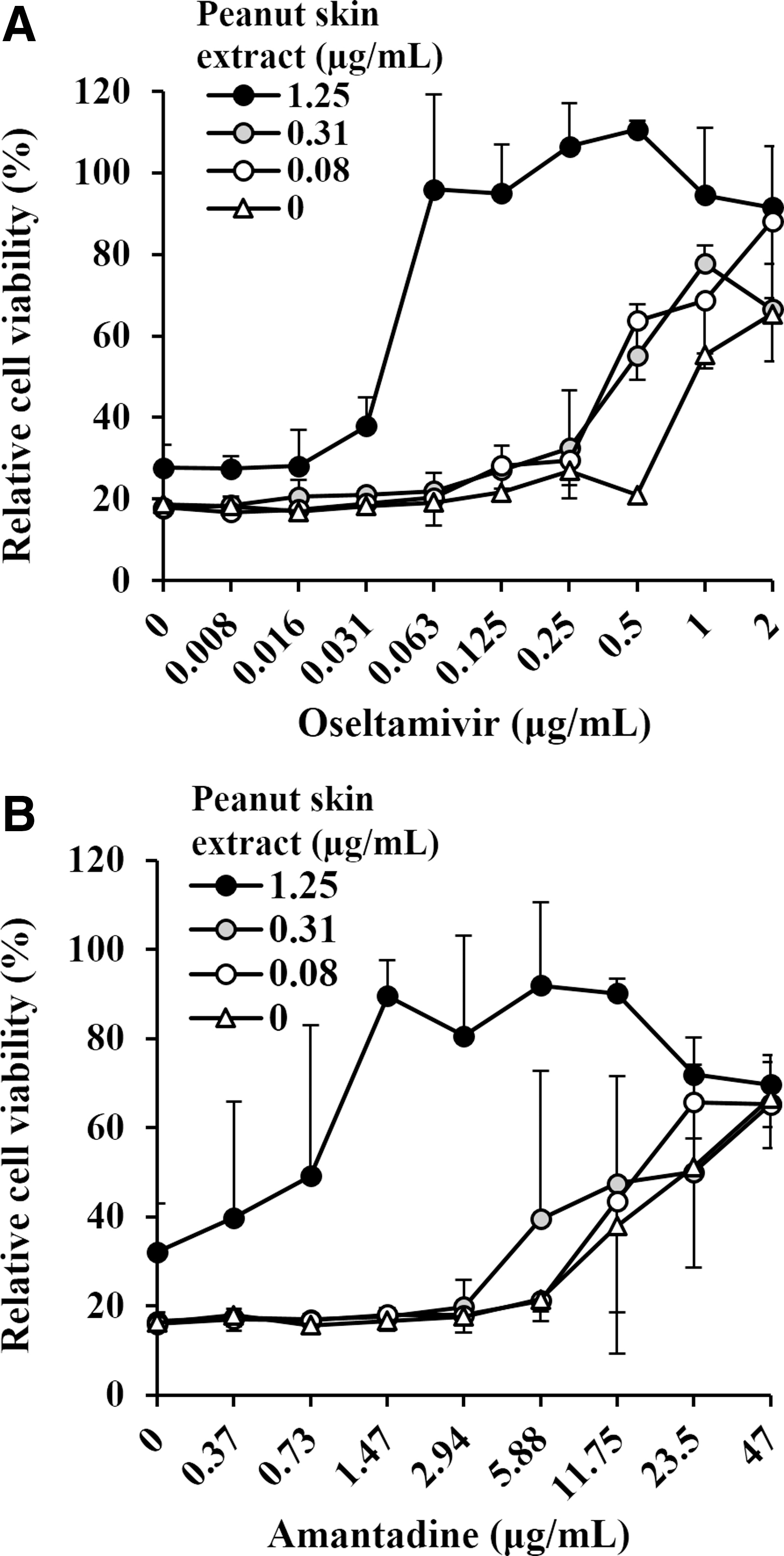
Effects of cotreatment with the peanut skin extract and anti-influenza drugs. MDCK cells were infected with the A/WSN/33 virus in the presence of
IC50: 50% inhibitory concentration, IC50 of peanut skin extract alone = 1.5 μg/mL.
Combination index as determined by Chou and Talay method, if it is <1 is considered synergistic, if = 1 or >1 is additive and antagonistic, respectively.
Sensitivity of various influenza viruses to peanut skin extract
The antiviral activity of peanut skin extract against type A and type B influenza viruses was investigated (Table 4). We found that the extract could inhibit the replication of all viruses tested, including A/WSN/33 (H1N1), A/Puerto Rico/8/34 (H1N1), A/Virginia/ATCC2/2009 (H1N1), A/Aichi/2/68 (H3N2), and B/Lee/40. When compared to oseltamivir, the extract had a close-range inhibitory activity for all viruses, with an IC50 range of 1.3–3.2 μg/mL. Moreover, the extract showed significantly higher activity than oseltamivir when A/Virginia/ATCC2/2009 (H1N1), a clinical isolate of the 2009 H1N1 pandemic flu and type B viruses, was used. These data indicate a broad spectrum of activity of peanut skin extract against influenza viruses, including oseltamivir-insensitive strains.
IC50: 50% inhibitory concentration.
Clinical isolate of A(H1N1)pdm09 influenza.
Discussion
The emergence of new influenza viruses and the spread of drug-resistant viruses highlight the urgent need to find new therapeutics. We investigated the anti-influenza activity of crude extracts prepared from the skins of peanuts obtained from different areas of the world. We found that peanut skins processed by pickling had a far lower polyphenol content than that of peanut skins processed by either roasting or blanching. Previous studies indicated that heat treatment of peanut skins increases their polyphenol content and antioxidant activity through the formation of Maillard products, and that ethanol extraction improved the recovery of polyphenols from roasted skins. 21,34 Our data showed that extracts with a high polyphenol content presented a high anti-influenza viral activity. Phenolic acids (coumaric, ferulic, chlorogenic, p-hydrobenzoic acids), phenolics (catechins, proanthocyanidins), and stilbenes (resveratrol) have been isolated from peanut skins. 22,35,36 Among these constituents, chlorogenic acid, 37 catechins, 13 proanthocyanidins, 38 and resveratrol, 32 isolated from other sources, have been reported as anti-influenza virus compounds. Resveratrol is also well known to have antiviral activities against human viruses such as HIV, 39 herpes simplex virus, 40 and other animal viruses 41 and is undergoing clinical trials for various human diseases. 42 We quantified the resveratrol content in the peanut skin extracts, (Table 2), and found that the resveratrol content correlated with neither polyphenol content nor antiviral activity of the extracts. In fact, sample H, which had the lowest polyphenol content and antiviral activity, contained the highest amount of resveratrol. It is thus conceivable that other polyphenolic constituents may be responsible for the anti-influenza virus activity. For instance, sample A (runner type) contained 6 μg/g of resveratrol, which is equivalent to 0.55 μg/g of resveratrol in dried peanut skins. This is in good agreement with a previous report that peanut skins from the runner and Virginia types contain ∼0.65 μg/g of resveratrol, 36 which is only 0.0016% of the total polyphenol content. Thus, which polyphenolic compound(s) of peanut skins contribute the antiviral activity remains to be determined.
The currently circulating influenza A viruses have lost their sensitivity to M2 ion channel inhibitors. 43 Therefore, neuraminidase inhibitors are recommended for prophylaxis and treatment of influenza virus infections. However, there are numerous reports of widespread resistance to oseltamivir among seasonal H1N1 viruses 2 and the 2009 H1N1 pandemic virus. 44 Combination chemotherapy is regarded as one possible way to curb the rapid emergence of drug resistance in the treatment of cancer, HIV, tuberculosis, and malaria, 45 and it has also been explored for influenza treatment. 46 Furthermore, during the outbreak of severe acute respiratory syndrome in 2002, the combination treatment of Chinese herbal medicine and drugs such as ribavirin and corticosteroids, among others, is reported to have alleviated clinical symptoms in patients, 47,48 indicating the possible application of herbal products to curb the spread of pandemic diseases.
The anti-influenza virus activity of amantadine and oseltamivir synergistically increased in the presence of peanut skin extract, which could be due to the enhanced effect achieved by targeting different replication stages of the influenza virus. Indeed, we found that anti-influenza activity of oseltamivir is largely affected by infection titer, whereas that of peanut skin extract is not (Fig. 1A, B), and the simultaneous treatment of virus and extract reduced plaque formation (Fig. 2). These results suggest that the extract inhibits the early stages of virus infection. More experiments are, however, required to clearly define the mechanism of action of the extract. A low dose (0.08 μg/mL) of peanut skin extract ∼85-fold lower than the CC50 value was enough to suppress influenza virus propagation in the presence of antiviral drugs (Table 3). Moreover, the peanut skin extract inhibited the replication of both types A and B influenza viruses. Thus, its combination with currently approved anti-influenza drugs presents a possible broad application for influenza management, especially against the 2009 H1N1 pandemic and type B viruses, both of which exhibited reduced sensitivity to oseltamivir. Therefore, the daily consumption of peanut skins as a supplementary food could be a new regimen to help prevent the circulation of pandemic influenza.
In conclusion, the ethanol extract of peanut skins showed anti-influenza activity and demonstrated its potential application in the management of influenza in combination with currently approved drugs. Because peanut skins are available worldwide at a low cost, and extracts contain high levels of polyphenols, they offer an inexpensive source of functional ingredients for protection against viral infections. Further studies are required to determine the in vivo antiviral activity of peanut skin extract and its bioactive constituents.
Footnotes
Acknowledgments
We thank Dr. Takahiro Haruyama of Nagasaki University for useful discussions, Ms. Tomoko Yamaguchi of the Nagasaki Advanced Computing Center (Nagasaki University) for technical assistance, Dr. Todd Saunders of Nagasaki University for English editing, and Mr. Tamio Tajima of the Japan Peanuts Association for collecting peanut skin samples and useful discussions. This work was supported by a grant from The Tojuro Iijima Foundation for Food Science and Technology (K.W.) and a grant from JSPS KAKENHI (grant number 16K00853 to K.W.).
Author Disclosure Statement
No competing financial interests exist.