
Abstract
Adipose tissue is a key organ with substantial senescent cell accumulation under both obesity and aging conditions. Chia seed is an ancient seed and is the richest plant source of α-linolenic acid. We aimed to determine how cellular senescence markers will be altered in adipose tissue of senescence-accelerated mouse-prone 8 (SAMP8) mice fed with high-fat diets (HFDs); and how chia seed can affect the above markers. SAMP8 mice and their control senescence-accelerated mouse-resistant 1 (SAMR1) were divided into four groups, that is, SAMR1 low-fat diet group (R1LF), SAMP8LF group (P8LF), SAMP8 high-fat group (P8HF), and SAMP8HF group supplemented with 10% chia seed (P8HC). At the end of the intervention, body composition was measured through T1-weighted magnetic resonance imaging, and epididymal (EPI) and subcutaneous (SC) adipose tissues were dissected for further analysis. Compared with the R1LF group, the P8HF and P8HC groups had significantly increased body fat mass. In EPI fat, p16, CD68 and PAI-1 mRNA expression from P8HF group were significantly increased; chia seed partially reduced p16 and CD68 mRNA expression. The P8LF group has increased p16 and CD68, and the P8HF group has increased p16, p21, and CD68; and P8HC group has increased p16 mRNA expression. The protein expression of p-AMPK in EPI and SC fat from the P8HF group was reduced. In conclusion, reductions in AMPK activity might be partially responsible for elevation in HFD-induced senescence markers in both EPI and SC fat, and chia seed supplementation is able to reduce senescence-associated markers at least in EPI adipose tissue.
Introduction
S
Chia seed (Salvia hispanica L.) was the major dietary component in the Mayan and Aztec populations. 6 Chia seeds contain the richest plant source of [α-linolenic acid (ALA) 18:3, n-3], and are also an important source of protein, fiber, minerals, and polyphenolic compounds, mainly including quercetin, myricetin and cholorogenic acid. 7 Evidence from both human 8 –11 and animal studies 12,13 have suggested that Chia seed is capable of improving glucose tolerance and insulin sensitivity, 8 –10,12 adiposity, 13 and dyslipidemia. 11,13 Recently, Ait-Ghezala et al. 14 reported that Chia seed extract activated telomerase activity in an established cell model of telomere shortening (i.e., IMR90 cells). This suggests that chia seed may help maintain telomere length and, thus, may lead to improved health during aging. Collectively, we hypothesized that chia seeds might improve adipose tissue function through its effects on senescence markers in the settings of both obesity and aging.
The senescence-accelerated mouse-prone 8 (SAMP8) mice is a well-established model for researching aging-associated neurodegenerative diseases, including Alzheimer's disease (AD). 15 Findings from our laboratory suggest that SAMP8 mice fed normal diets also exhibit multiple metabolic characteristics associated with age in adipose tissue, such as elevation in endoplasmic reticulum stress markers and inflammation, as well as reduction in AMPK activity. 16 Of interest, Mehla et al. 17 reported that when fed high-fat diets (HFDs), SAMP8 mice would demonstrate worsened AD-related pathology compared with low-fat diet-fed SAMP8 mice, which indicates that high fat (HF)-induced SAMP8 mice may also demonstrate worsened metabolic dysfunction in adipose tissue.
With the above points in mind, the aim of this study was (1) by utilizing HF fed SAMP8 mice as a model, to determine how senescence markers will be altered in adipose tissue with aging and HFD; and (2) to determine how chia seed supplementation will affect the above markers in adipose tissue from HF-fed SAMP8 mice.
Materials and Methods
Materials
Reagents for sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) were from Beyotime Institute of Technology (Jiangsu, China). Molecular weight marker and nitrocellulose membranes for SDS-PAGE were from Bio-Rad (CA, USA). Immobilon western chemiluminescent horseradish peroxidase (HRP) substrate (cat# WBKLS0100) was purchased from Millipore (MA, USA). Antibodies against p-AMPK (cat# 2535) and SIRT1 (cat# 8469) were purchased from Cell Signaling (MA, USA). HRP-conjugated donkey anti-rabbit and goat anti-mouse IgG secondary antibodies were purchased from Jackson ImmunoResearch Laboratories (PA, USA). The Universal RNA Extraction Kit, PrimeScript RT Master Mix Kit, and Premix Ex TaqTM (Probe qPCR) were from TaKaRa Bio (Shiga, Japan). TaqMan Gene Expression Assays for Eukaryotic 18S rRNA (cat# 4333760T), p16 (cat# Mm00494449_m1), p21 (cat# Mm04205640_g1), p53 (cat# Mm01731290_g1), CD68 (cat# Mm03047343_m1), and PAI-1 (cat# Mm00435858_m1) were from Applied Biosystems (Foster City, CA, USA). Chia seed was obtained from NOW FOODS with a purity of ≥98% (IL, USA).
Treatment of animals
Ten-week-old male senescence-accelerated mouse-resistant 1 (SAMR1) (n = 8) and SAMP8 mice (n = 24) were purchased from Animal Service of Health Science Center, Peking University (Beijing, China). All animals were treated in accordance with the Guidelines in the Care and Use of Animals and with the approval of the Animal Studies Committee of Soochow University, Suzhou, China. Mice were housed together in groups of two animals in standard plastic mouse cages (30 × 20 × 13 cm3) and were acclimated to the animal housing facility for 1 week before the start of intervention. At 12 weeks of age, mice were randomly divided into four groups of eight animals each, that is, SAMR1 low-fat diet group (R1LF), SAMP8 low-fat diet group (P8LF), SAMP8 HFD group (P8HF), and SAMP8 HFD combined with chia seed intervention group (P8HC). For the R1LF and P8LF group, mice were allowed a low-fat diet ad libitum during the entire experimental period (i.e., 18 weeks). The low-fat diet was purchased from Research Diets Inc. (cat# D12450J, NJ, USA) and contained fat 10 Kcal%, carbohydrate 70 Kcal%, and protein 20 Kcal%. For the P8HF group, mice were allowed a HFD ad libitum during the entire experiment (i.e., 18 weeks). The HFD contained fat 60 Kcal%, carbohydrate 20 Kcal%, and protein 20% Kcal, and was purchased from Research Diets Inc. (cat# D12492). For the P8HC group, mice were allowed a HFD ad libitum supplemented with 10% chia seed (i.e., 10 g chia seed powder dissolved in 100 g HFD during the entire experiment. The HFD was the same as above and chia seed powder was added into the food following the recipe prepared by the researchers. We chose these dosages of chia seed mainly based on published findings from other research groups. 12,18 Body weight and food intake were determined every other week for the duration of the study. Epididymal (EPI) and subcutaneous (SC) adipose tissue were dissected at the end of the study for further analysis.
T1-weighted magnetic resonance imaging
At the end of the intervention, T1-weighted magnetic resonance imaging (MRI) was performed using NM-G1 imaging instrument (Shanghai Niumag Corp., Shanghai, China). Mice were anesthetized with ethyl ether. The instrumental parameters were set as follows: a 1.0T magnet, TR = 300 ms, TE = 18.2 ms, Averages = 12, Slice width = 2 mm. The fat highlight image was analyzed by imageSystem (Shanghai Niumag Corp.).
Histology
Approximately 50 mg of EPI and SC fat were fixed in 10% neutral-buffered formalin, dehydrated in 70% ethanol, and embedded in paraffin. Five micrometer sections were mounted on 1.2 mm Superfrost slides, stained with Hematoxylin and Eosin (H&E) stock and imaged (Olympus BX-51 light microscope; Olympus, Tokyo, Japan). Cells were sampled in each image to determine cross-sectional area (>200 cells/sample) through Image-Pro Plus6.0 software (Media Cybernetics, MD, USA).
Western blot analysis
Protein was extracted from the EPI and SC fat of the mice, and the content of p-AMPK/AMPK and SIRT1 were determined by western blotting as described previously by our laboratory. 19 –21 Signals were visualized using Immobilon Western Chemiluminescent Hrp Substrate and captured using a Syngene chemi-imaging system (MD, USA). Subsequently, bands were quantified by densitometry through Gene Tool according to the manufacturer's instructions (Syngene, ChemiGenius2; PerkinElmer). Protein contents of phosphorylated protein were quantified and normalized to the total levels of these proteins. Beta actin was used as internal controls.
Real-time PCR
RNA was isolated from adipose tissue using the RNeasy Kit according to the manufacturer's instructions and 1 μg of RNA was used for the synthesis of complementary DNA using the PrimeScript RT Master Mix Kit. Real-time PCR was performed using a 7500 Real-Time PCR system (Applied Biosystems). Results were normalized to the mRNA expression of 18S. Relative differences in gene expression between groups were determined using the 2−ΔΔCT method. 22 The amplification efficiencies of the gene of interest and the housekeeping gene were equivalent.
Statistical analysis
Data are expressed as mean ± standard error of the mean. Comparisons between groups were made using a one-way ANOVA followed by Tukey's post hoc test. Statistical significance was established at a P < .05.
Results
Final body weight, liver weight, and fat pad mass
As shown in Table 1, there were no differences in final body weight and liver weight among the groups. Compared with both R1LF and P8LF groups, the P8HF group had elevated EPI fat weight.
R1LF, SAMR1 low-fat diet group; P8LF, SAMP8 low-fat diet group; P8HF, SAMP8 HFD group; P8HC, SAMP8 HFD combined with chia seed intervention group.
P < .05 versus R1LF; # P < .05 versus P8LF.
EPI, epididymal; HFD, high-fat diet; SAMR, senescence-accelerated mouse-resistant.
Body composition and H&E staining
T1-weighted MRI was used to evaluate body fat distribution in mice. As shown in Figure 1A, compared with R1LF group, P8HF and P8HC groups showed significant signal strength (lightness), which indicates that these groups have significantly higher body fat mass. As shown in Figure 1B and C, H&E staining of EPI and SC fat demonstrated that compared with both R1LF and P8LF groups, the P8HF group had significantly enlarged adipocytes in both EPI and SC fat; furthermore, the adipocytes in SC fat from P8HC group were also significantly enlarged.
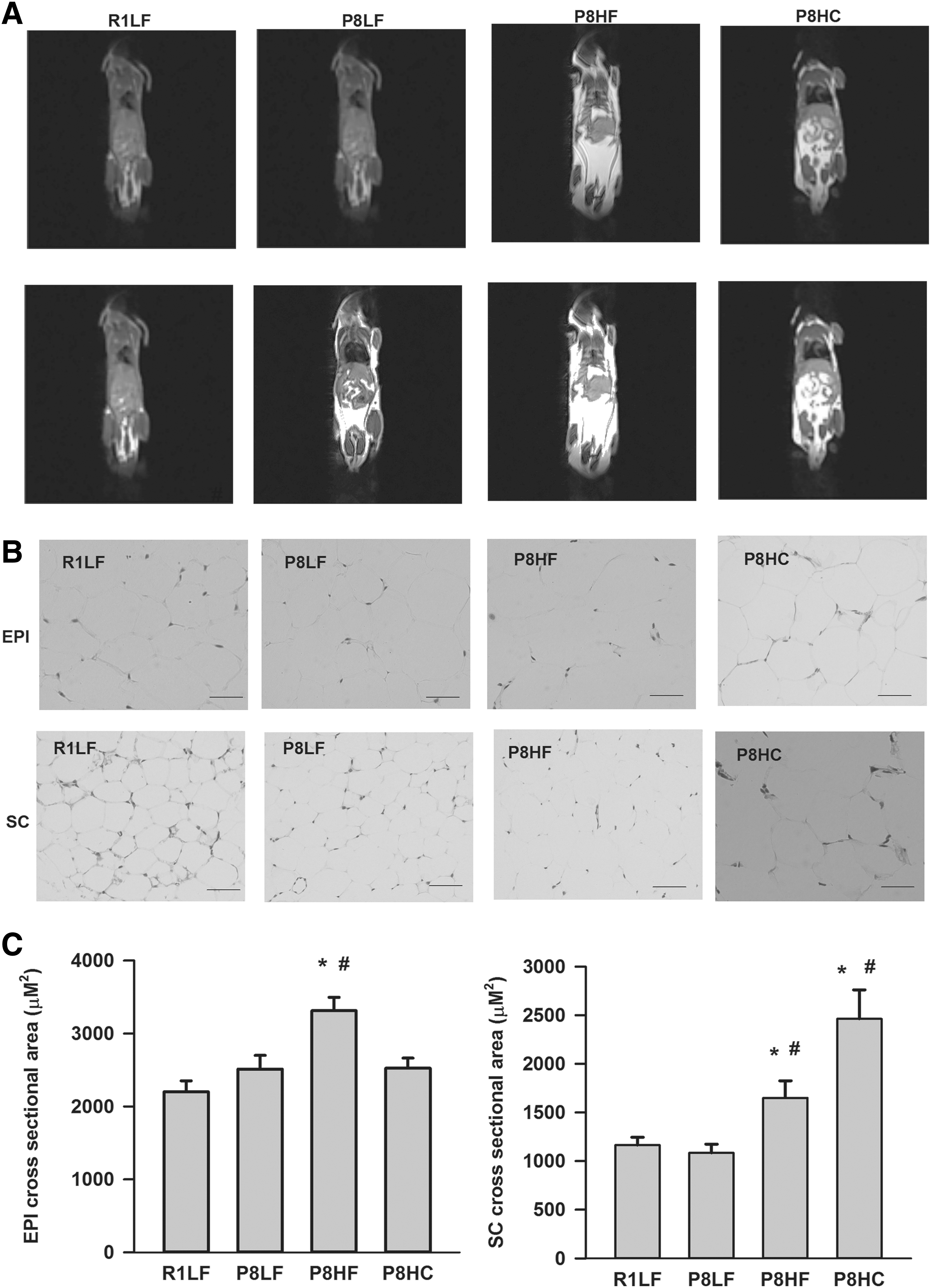
Body composition and H&E staining for EPI and SC fat.
Fasting plasma glucose, insulin and homeostasis model assessment of insulin resistance levels
As shown in Table 2, there were no differences in fasting plasma glucose levels among the groups. Compared with R1LF group, P8LF group had lower insulin levels; the P8HC group had lower insulin and homeostasis model assessment of insulin resistance (HOMA-IR) levels; and compared to P8LF group, P8HC group also had lower insulin levels.
R1LF, SAMR1 low-fat diet group; P8LF, SAMP8 low-fat diet group; P8HF, SAMP8 HFD group; P8HC, SAMP8 HF combined with chia seed intervention group.
P < .05 versus R1LF; # P < .05 versus P8LF.
HOMA-IR, homeostasis model assessment of insulin resistance.
Markers of cellular senescence mRNA expression in EPI and SC fat
Compared with R1LF group, the mRNA expression of p16, CD68, and PAI-1 in EPI fat from P8HF group were significantly increased (∼2–2.5 fold); compared with P8HF group, the mRNA expression of p16 and CD68 in EPI fat from P8HC group were significantly reduced to 60% and 80%, respectively (Fig. 2A). As for SC fat, in comparison with the R1LF group, the P8LF group had significantly higher p16 (∼10-fold) and CD68 (∼3-fold) mRNA expressions, P8HF group had significantly increased p16, p21, and CD68 mRNA expression (∼14, 3, and 3-fold, respectively); and the P8HC group also had significantly increased p16 mRNA expression (∼18 fold) (Fig. 2B).
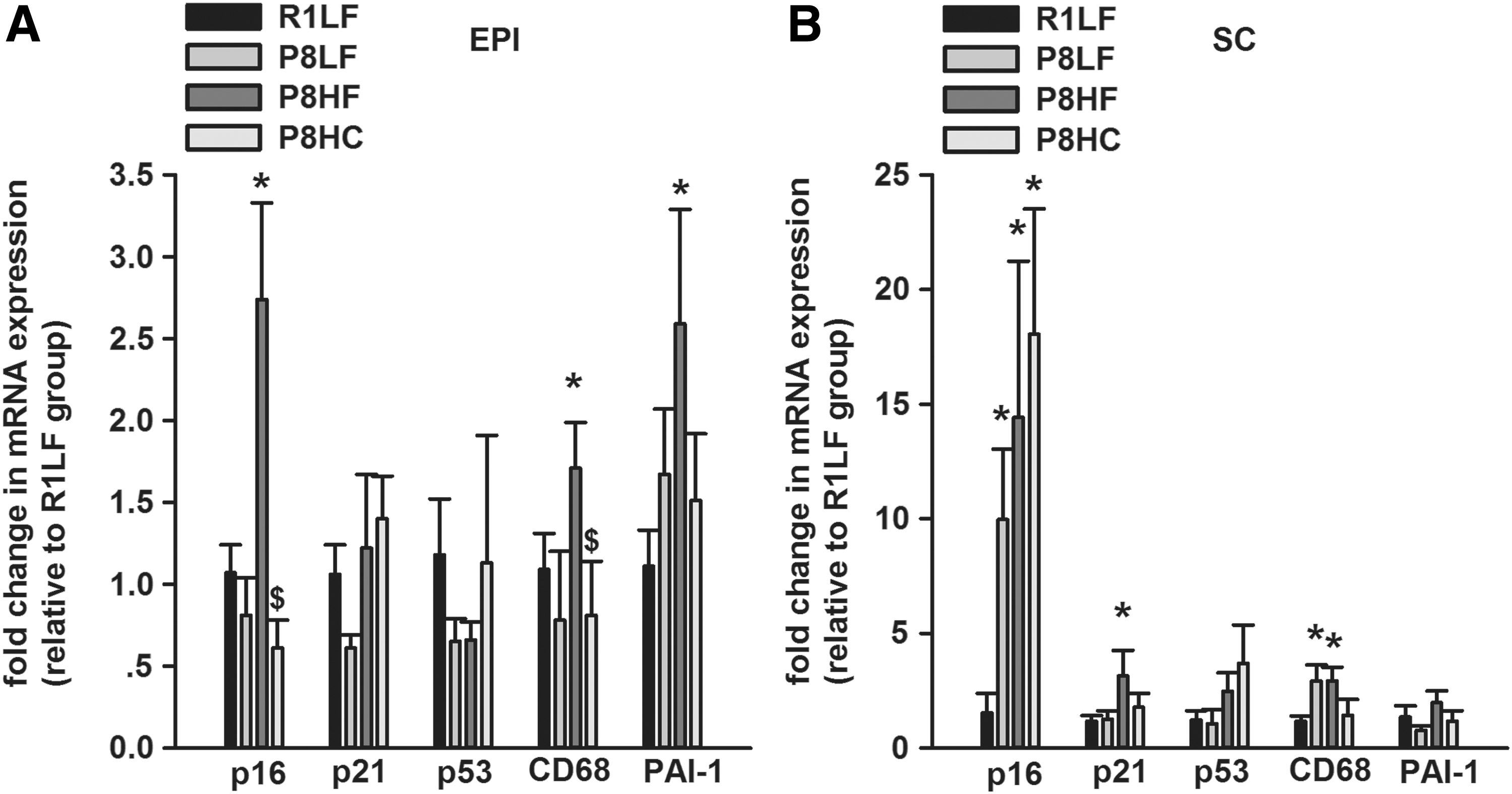
Cellular senescence-associated markers mRNA expression in both EPI and SC fat. The mRNA expression of p16, p21, p53, CD68, and PAI-1 from EPI
AMPK/SIRT1 protein expressions in EPI and SC fat
Compared with R1LF group, the protein expression of p-AMPK in EPI and SC fat from P8HF group were significantly reduced; there was no difference in SIRT1 protein expression among groups (Fig. 3A, B).

The phosphorylation of AMPK and SIRT1 protein expression in both EPI and SC fat. The protein expression of p-AMPK and SIRT1 from EPI
Discussion
The primary findings of the present study are that HFD elevated key senescence-related markers and reduced p-AMPK in both EPI and SC fat from SAMP8 mice, whereas chia seed supplementation only reduced p16 and CD68 mRNA expression in EPI fat; however, chia seed was able to improve insulin sensitivity in HFD SAMP8 mice with no alterations in final body weight.
Although there is no difference for final body weight among groups, MRI demonstrated that both P8HF and P8HC groups have increased total body fat mass; H&E staining of EPI and SC fat further confirmed that enlarged adipocytes existed in both EPI and SC fat from P8HF group, as well as in SC fat from P8HC group. A significant reduction in plasma insulin and HOMA-IR from the P8HC group suggested that chia seed is able to improve insulin resistance in HFD-fed SAMP8 mice. Taken collectively, it is suggested that HF induced enlarged adipocytes in both EPI and SC fat from SAMP8 mice, with no alteration in body weight, and that chia seed is unable to reverse this effect; but chia seed was efficacious preventing or reversing insulin resistance. Previously, Marineli et al. 12 reported that both short (3 weeks) and long-term (12 weeks) chia seed supplementation in HF and high-fructose diet-fed rats had no effects on body weight gain, which is consistent with our present study. However, our findings are inconsistent with the findings by Chicco et al., 13 where it was demonstrated that chia seed supplementation (2 months) in sucrose-rich diet (SRD) induced obese rats, led to a reduction in body weight and glucose levels, with no alterations in insulin level. This might be owing to species difference (SAMP8 mice vs. rats), diet intervention (HFD vs. SRD), chia seed supplementation duration (18 weeks vs. 2 months) etc. Nevertheless, our study suggests that chia seed supplementation might improve insulin sensitivity in HFD-fed SAMP8 mice, although with no effects on body weight.
It is inevitable that senescent cell accumulation occurs with advancing age; however, prematurely elevated senescent cell burden may be both a cause and consequence of metabolic dysfunction. 23,24 We found that compared with R1LF control, the P8LF group had increased p16 and CD68 mRNA expression only in SC, but not in EPI fat, which indicates that under low-fat diet condition, in adipose tissue of SAMP8 mice, senescence-associated markers occur in SC fat before EPI fat. This is in contrast with the findings by Schafer et al., 25 who reported that in 8-month-old mice harboring an EGFP transgene driven by the p16 INK4α promoter, 16 weeks of western diet (fat 40 Kcal% with 0.2% cholesterol) induced senescent markers in visceral fat, but not in SC fat. This might be due to diet composition differences or species differences. However, HFD resulted in elevated senescence-associated markers in both EPI and SC fat, suggesting that HFD consumption per se would induce adipocyte senescence. This is in agreement with the findings by Schafer et al. 25 and List et al. 4 We further demonstrated that chia seed supplementation is able to reduce p16 and CD68 mRNA expression only in EPI fat, which indicates that chia seed is efficacious for reversing aging and HFD-induced senescence markers at least in EPI fat. It is likely that chia seed supplementation might also improve senescence-associated markers in other fat depots, such as SC, omental, and retroperitoneal adipose tissue with longer intervention duration. Nevertheless, our study is the very first to demonstrate that chia seed supplementation might be a promising strategy for reversing aging and diet-induced cellular senescence, at least in adipose tissue.
SIRT1 is involved in regulating a wide variety of cellular processes, including metabolism, mitochondrial biogenesis, antioxidative stress responses, and anti-inflammatory pathways. 26 A decrease in SIRT1 has been described in neurons of SAMP8 mice, 27 as well as aging endothelial cells and vascular tissue undergoing senescence. 28 AMPK is a metabolic sensor that maintains energy balance within a cell. 29 Evidence suggests that AMPK is also essential for antisenescence. 30 It has been well established that AMPK and SIRT1 have a long-standing partnership, 31 and it is likely there are interconnections between AMPK and SIRT1 that are involved in regulating aging. We observed reductions in AMPK activity in both EPI and SC fat only in P8HF group, indicating that reduction in AMPK activity might occur before SIRT1 reduction, and that reduced AMPK from P8HF group is at least partially responsible for the elevation in senescent markers in both EPI and SC fat.
In conclusion, we demonstrated that reduction in AMPK activity might be partially responsible for the elevation in HF-induced senescence markers in both EPI and SC fat, but chia seed supplementation is able to reduce senescence-associated markers at least in EPI adipose tissue.
Footnotes
Acknowledgments
This work is supported by grants 2017YFC1310700, and 2017YFC1310701 from National Key R&D Program of China, the Natural Science Foundation of China (grant no. 81472975), China Postdoctoral Science Foundation (grant no. 2016 M591918), Postdoctoral Science Foundation of Jiangsu Province (grant no. 1601046A), and scientific and technological projects of Suzhou City (grant no. SYS201707).
Author Disclosure Statement
No competing financial interests exist.