
Abstract
The stem of Opuntia ficus-indica var. saboten is edible and has been used as a medicinal herb on Jeju Island in Korea. We previously reported that the butanolic extract of O. ficus-indica var. saboten exerts the enhancement of long-term memory in mice. However, the antiamnesic effects of O. ficus-indica var. saboten and its mode of action has not been clearly elucidated. In the present study, we explored the effects of the ethanolic extract of stems of O. ficus-indica var. saboten (EOFS) on cognitive performance in mouse and attempted to delineate its mechanism of action. We used the passive avoidance, Y-maze, and novel object recognition tests to assess its effects on cognitive functions in scopolamine-induced memory-impaired mice. We observed that EOFS (100, 200, and 400 mg/kg) ameliorated scopolamine-induced cognitive dysfunction. We also explored its mechanism of action by conducting an acetylcholinesterase (AChE) activity assay using the mouse whole brain and Western blot using the mouse hippocampal tissue. Western blot analysis and the ex vivo study revealed that EOFS increased the levels of phosphorylated extracellular signal-regulated kinase and cAMP response element-binding protein (CREB) and the levels of brain-derived neurotrophic factor (BDNF) expression in the hippocampus. It also inhibited AChE activity in the brain. Our findings suggest that EOFS would be useful for the treatment of cholinergic blockade-induced cognitive dysfunction.
Introduction
A
Donepezil, rivastigmine, and galantamine have been used in clinics, and these agents enhance the activity of the cholinergic neurotransmitter system by the inhibition of acetylcholinesterase (AChE) activity. 5 These drugs exhibit therapeutic potential in patients with moderate AD, but some adverse effects were reported, such as nausea, vomiting, or diarrhea. 6 These results underscore the need for better AChE inhibitors with fewer adverse effects. 7 Hence, we investigated some antiamnesic or antidementia agents from traditional and edible herbal materials, because these materials exhibit fewer safety issues. 8 These agents may have the potential to be effective alternatives while overcoming the adverse effects of AD medications.
Opuntia ficus-indica var. saboten, Makino (Cactaceae), also known as prickly pear cactus and nopal, is a tropical or subtropical herb and is broadly cultivated on Jeju Island in Korea. 9 It is used as folk medicine on Jeju Island as a treatment for burns, dyspepsia, and asthma. Indeed, a previous study showed that O. ficus-indica var. saboten has analgesic or anti-inflammatory effects. 10
This plant contains many potentially active ingredients, such as, minerals or vitamins, and it has received attention as a health-promoting food ingredient. 11 –13 Notably, O. ficus-indica, the origin plant of O. ficus-indica var. saboten, has shown to be a potential treatment for classical neurodegenerative diseases, such as AD or Parkinson's disease. 14 In our previous study, we reported that the butanolic extract of O. ficus-indica var. saboten exhibits long-term memory enhancement in mice. 15 Furthermore, our recent pilot study demonstrated that an ethanolic extract of O. ficus-indica var. saboten (EOFS) exerts inhibitory activities against AChE. These findings prompted us to examine whether O. ficus-indica var. saboten has anti-amnesic activities and elucidate its mechanisms of action.
In this present study, we investigated whether the EOFS can ameliorate scopolamine-induced cognitive impairments in mice using the passive avoidance, Y-maze, and novel object recognition paradigms. AChE activity was examined in whole brain tissue, and western blot was performed using hippocampal tissue to elucidate its mechanism of action.
Materials and Methods
Animals
Six-week-old CD-1® male ICR mice (25–30 g) were purchased from the Orient Co. Ltd., a division of Charles River Laboratories (Gyeonggi-do, Korea). Animals were housed five per cage (27 × 22 × 14 cm), provided with food and water ad libitum, and kept on a 12-h light/12-h dark cycle (light on at 7:30 and off at 19:30) in a room with constant humidity (60% ± 10%) and temperature (23°C ± 1°C). All animal experiments were carried out in accordance with the Animal Care and Use Guidelines issued by Kyung Hee University, Republic of Korea. The experimental protocol using animals was approved by the Institutional Animal Care and Use Committee (No. KHUASP(SE)-16-084).
Materials
Scopolamine hydrobromide and donepezil hydrochloride monohydrate were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Scopolamine and donepezil were used after dissolving in a 0.9% saline solution. Rabbit polyclonal anti-phosphorylated extracellular signal-regulated kinase (pERK) antibody was purchased from Cell Signaling Technology (Danvers, MA, USA). Rabbit polyclonal anti-ERK or anti-β-actin antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Rabbit polyclonal anti-cAMP response element-binding protein (CREB), anti-phosphorylated CREB (pCREB) or anti-brain-derived neurotrophic factor (BDNF) antibodies were purchased from Millipore (Temecula, CA, USA). All other agents were acquired from common commercial sources and were of the highest grade available.
Preparation of herbal extracts
Stems of O. ficus-indica var. saboten were collected in March 2016 on Jeju Island, and ground into a powder after cutting and hot air drying. Powder (160 kg) was placed in a fermenter and extracted with 2400 L of 70% ethanol at 80°C for 2 h under stirring at 100 g. The extracts were filtered, and the filtrate was concentrated to 15–20 Brix. The same amount of dextrin compared with solid content was added into the concentrate. The concentrate was autoclaved at 95°C for 1 h and spray dried. The spray-dried powder was passed through a 60-mesh sieve. The final product of EOFS was prepared and packaged (yield; 31.6% ± 1.1%, n = 3).
The final product was standardized using dihydrokaempferol, which is a marker compound of 70% EOFS, for quality consistency. The content of dihydrokaempferol was calculated using a calibration curve (Fig. 1). The content of dihydrokaempferol in the final product was 0.506 mg/g.

Dihydrokaempferol
Passive avoidance test
The passive avoidance test was used to measure the effects of EOFS on long-term memory in mice.
The test was conducted on two consecutive days as an acquisition and retention trial. The passive avoidance apparatus consists of two cuboid chambers (20 × 20 × 20 cm), a light chamber (50 W bulb) and a dark chamber without light source. The two chambers were separated by a guillotine door (5 × 5 cm), as described elsewhere. 16 Both chambers were equipped with stainless steel bars for electric shock. Mice received EOFS (50, 100, 200, or 400 mg/kg, p.o.) or donepezil (5 mg/kg, p.o) 1 h before the test for the acquisition. The doses of EOFS employed in the present study were determined by a pilot study and our previous work. 15 Donepezil was used as a positive control. The control group received vehicle solution (0.9% saline). Scopolamine (1 mg/kg, i.p.) was administered 30 min before the test.
Mice in the acquisition trials were gently placed in the light chamber, and the guillotine door between the two chambers was opened 10 sec later. The guillotine door automatically closed when the mouse entered the dark chamber, and electrical shock (0.5 mA, 3 sec) was applied through the stainless steel bars. The mouse was forced to enter the dark chamber if it did not enter the dark chamber within 60 sec, and a latency of 60 sec was recorded. The retention trial was performed 24 h after the acquisition trial. The mouse was placed into the light chamber again on the testing day, and the time taken to enter the dark one was recorded for each mouse.
If a mouse did not enter the dark chamber within 300 sec, it was assumed that the mouse remembered the foot stimuli from the acquisition trial and recorded the latency as 300 sec. The latency was defined as the time to enter the dark chamber for each trial.
Y-maze test
The Y-maze test was employed to examine the effects of EOFS on working memory. The Y-maze was composed of a three-arm horizontal maze (40-cm long and 3-cm wide with 12-cm high walls) in which the arms were placed at 120 degree angles to each other. The floor and walls of the maze were composed of dark opaque polyvinyl plastic as previously described. 17 Mice were treated with various doses of EOFS (50, 100, 200, or 400 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.) as a positive control 1 h before testing, and memory impairment was caused by scopolamine administration (1 mg/kg, i.p.) 30 min later. The control group animals were treated with vehicle solution (0.9% saline) rather than EOFS or donepezil.
Mice were placed in one arm of the Y-maze 30 min after scopolamine administration and were allowed to investigate all three arms of maze freely for 8 min. All behaviors were recorded using a video tracking system, and a researcher who was blinded to treatment manually analyzed the sequence (i.e., ABCCAB) and number of arm entries. An actual alternation was considered as entries into all three arms with consecutive choices (i.e., ABC, CAB, or BCA, but not ABA). Each arm was thoroughly cleaned between each task to remove residual odors. The percentage of alternation score (%) for each mouse was calculated using the following equation: % alternation = [(number of alterations)/(total arm entries −2)] × 100. The number of entries into each arm was used as an indicator of locomotor activity. 18
Novel object recognition test
The novel object recognition test was conducted, as described elsewhere. 19 The novel object recognition task was performed in a black polyvinyl plastic square open field (25 × 25 × 25 cm). The experiment was performed for 4 days and included the habituation trial, the training trial, and the testing trial. The habituation trial was performed for 2 days by exposing the animal to the experimental equipment for 10 min per day in the absence of objects. The animal was introduced into a training trial 24 h after the last habituation training. EOFS (50, 100, 200, or 400 mg/kg, p.o.) or donepezil (5 mg/kg, p.o.) was administered 1 h before the training trial. The control group received vehicle solution (0.9% saline) rather than EOFS or donepezil. Memory impairment was produced through the administration of scopolamine (1 mg/kg, i.p.) 30 min before the training trial.
Two identical objects were placed in the experimental arena during the training trial, and the mice were left to explore each object for 5 min. Mice were again placed in the apparatus for the testing trial after a 24-h retention interval. One of the objects was exchanged by a novel object, and the mice were left to explore each object for 5 min in the apparatus.
All behaviors were recorded using a video tracking system. The objects used in this experiment were a metal cylinder and a rectangular plastic block of approximately the same height. The apparatus and each object were cleaned using a paper towel and a 70% ethanol spray between each test. A researcher who was blinded to treatment history analyzed the amount of time spent investigating each object (familiar object, Tfamiliar; novel object, Tnovel) and the total investigation time of novel and familiar objects, preference ratio for novel versus familiar objects, and discrimination ratio [(Tnovel − Tfamiliar)/(Tnovel + Tfamiliar) × 100] were calculated.
AChE activity assay
The AChE activity was assayed using the acetylthiocholine iodide-based colorimetric method according to the method of Ellman et al. (1961). Mice were treated with EOFS (50, 100, or 200 mg/kg, p.o.), donepezil (5 mg/kg) as a positive control, or the same volume of vehicle (0.9% saline, p.o.) and sacrificed by decapitation 1 h after administration. The EOFS-treated groups and donepezil-treated group received scopolamine (1 mg/kg, i.p.) 30 min after the above treatments.
Whole mouse brain, except cerebellum, was homogenized in the homogenizing buffer (10 × volume of each tissue weight, 100 mM phosphate buffer, pH 8.0) using a glass Teflon homogenizer (Eyela, Japan) and centrifuged at 14,000 g for 10 min at 4°C. The supernatant was collected and used as a crude enzyme source for the assay. The supernatant solution in a 96-well plate was mixed with 144 μL of phosphate buffer (100 mM, pH 8.0), 22 μL of Ellman's solution [10 mM of 5,5-dithiobis-(2-nitrobenzoic acid) and 15 mM of sodium bicarbonate], and 1.1 μL of acetylthiocholine iodide solution (75 mM), and the reaction mixtures were incubated at room temperature for 10 min.
Thereafter, the reaction was stopped by adding 4.4 μL of a neostigmine solution (100 mM). By using a UV spectrophotometer (OPTIZEN 2120UV, Mecasys Co., Ltd., Korea), absorbance was taken at 412 nm. The percentage of inhibition was calculated through comparison with the AChE activity of the vehicle-treated control group.
Western blot
Mice were treated with EOFS (50, 100, or 200 mg/kg, p.o.) or donepezil (5 mg/kg), and scopolamine (1 mg/kg, i.p.) was administered to the same animal 30 min after the above treatments. Hippocampi from each mouse were isolated 30 min after scopolamine treatment.
Tissues were then homogenized in ice-cold Tris–HCl buffer (20 mM, pH 7.4) solution which was prepared in advance. The homogenizing buffer solution contained 0.32 M sucrose, 1 mM EDTA, 1 mM EGTA, 1 mM PMSF, 1 mM sodium orthovanadate, and protease inhibitor tablet (1 tablet per 50 mL buffer).
The lysate was centrifuged at high speed (14,000 g) for 20 min at 4°C. Homogenates (15 μg of total protein) were subjected to SDS-PAGE (8% or 12% (only for BDNF) gel), and gels were run under reducing conditions. Proteins separated by SDS-PAGE were transferred to polyvinylidene difluoride (PVDF) membranes in Tris–HCl transfer buffer (25 mM, pH 7.4) containing 192 mM glycine and 20% methanol (v/v) at 4°C and 400 mA for 2 h. The membranes were blocked with 5% skim milk in Tris-buffered saline/Tween 20 (TBST) for 2 h at room temperature.
Thereafter, the membranes were incubated with primary antibodies [anti-ERK or anti-pERK (1:5000), anti-CREB or anti-pCREB (1:3000), anti-β-actin (1:5000), and anti-BDNF (1:1000)] overnight at 4°C and washed with TBST (six times). Each primary antibody was diluted in 2% skim milk. The washed membranes were treated with a 1:5000 dilution of a horseradish peroxidase-conjugated secondary antibody and incubated for 2 h at room temperature. Finally, the immunoblots were developed using enhanced chemiluminescence (Amersham Life Science, Arlington Heights, IL, USA). The images of immunoblots were captured using the LAS-4000 mini bioimaging program (Fujifilm Lifescience USA, Stamford, CT) and analyzed by ImageJ analysis program.
Statistical analysis
Data were presented as the mean ± SEM. Latencies in the passive avoidance test, spontaneous alternations in the Y-maze test, the object preference ratio discrimination ratio in the novel object recognition test, and data from western blotting were analyzed using one-way analysis of variance (ANOVA). For multiple comparison tests, the Student–Newman–Keuls test was used. The level of statistical significance was set at P < .05.
Results
The effect of EOFS on cognitive dysfunction induced by cholinergic blockade in the passive avoidance test
We used the passive avoidance test to examine whether EOFS ameliorated scopolamine-induced memory impairment. In the retention trial, there were significant group effects on the step-through latencies [F(6, 62) = 42.38, P < .05, Fig. 2]. EOFS (100, 200, or 400 mg/kg) and donepezil (5 mg/kg) administration significantly reversed the reduced step-through latency of scopolamine administration (P < .05). In the acquisition trial, however, no significant differences were observed in the step-through latency across groups.
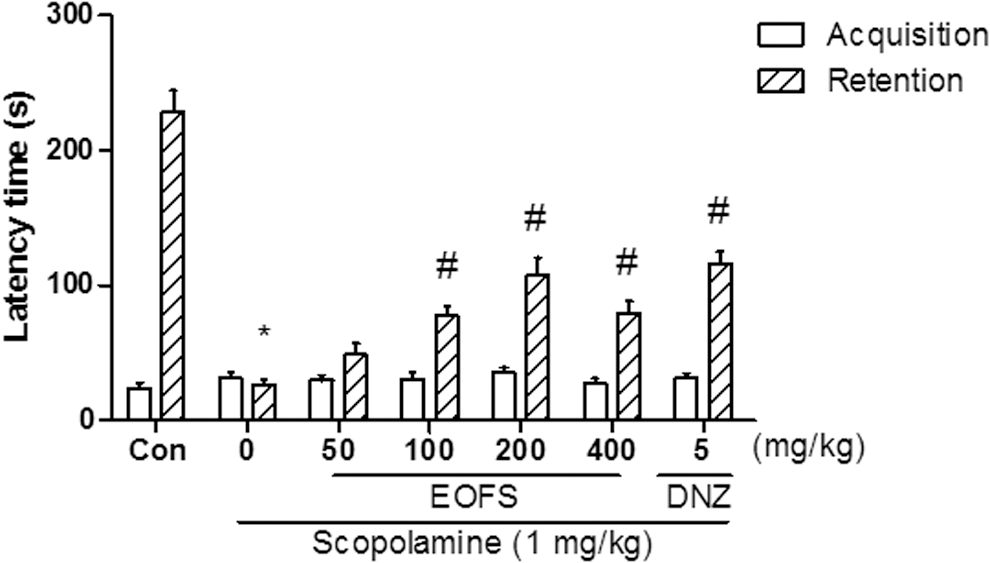
Effects of the EOFS (50, 100, 200, and 400 mg/kg, p.o.) on scopolamine-induced memory deficits in the passive avoidance test in mice. Results from the acquisition trial and retention trial are presented. Control group (CON) received vehicle solution (0.9% saline). Donepezil (DNZ) was used as a positive control. Data represent the mean ± SEM. (n = 8–10/group) (*P < .05, vs. the vehicle-treated control; # P < .05, vs. the scopolamine-treated group).
The effect of EOFS on the cognitive impairment caused by cholinergic blockade in the Y-maze test
We conducted the Y-maze test to investigate the effect of EOFS on spontaneous alternation behavior. Spontaneous alternation behavior was significantly different between EOFS administration groups [F(6, 59) = 5.560, P < .05, Fig. 3A]. EOFS (200 mg/kg) and donepezil (5 mg/kg) administration significantly reversed the scopolamine-induced decrease in the percentage of spontaneous alternations (P < .05). In contrast to the data from the passive avoidance task, the percentage of spontaneous alternations was not significant at the low or high dose of EOFS (100 or 400 mg/kg). The number of entries into each arm was no significant differences among the experimental groups (Fig. 3B), which indicate that EOFS treatment did not produce any change of general locomotor activity.

Effects of the EOFS (50, 100, 200, and 400 mg/kg, p.o.) on scopolamine-induced memory deficits in the Y-maze test in mice. The results from the spontaneous alternation
The effect of EOFS on cholinergic blockade-induced memory impairment in the novel object recognition test
We used the novel object recognition test to evaluate the effect of EOFS on object recognition memory in mice. There were significant group effects in the object preference ratio [F(6, 62) = 2.274, P < .05, Fig. 4A] and the discrimination ratio [F(6, 62) = 5.585, P < .05, Fig. 4B]. The scopolamine-induced cognitive impairments, as denoted by decreases in the preference for the novel object and the discrimination ratio between the novel object and familiar one, were significantly attenuated by EOFS (100 or 200 mg/kg, P < .05) and donepezil (5 mg/kg, P < .05). However, total exploration times were not significantly different across treatment groups for novel and familiar objects [F(6, 62) = 1.668, P > .05, Fig. 4C].

Effects of the EOFS (50, 100, 200, and 400 mg/kg, p.o.) on scopolamine-induced memory deficits in the novel object recognition test in mice. The results from the percentage of preference on the novel or the familiar objects
The effect of EOFS on AChE activity
We investigated whether EOFS inhibited AChE activity under ex vivo conditions and observed significant group effects [F(5, 24) = 5.573, P < .05, Fig. 5]. AChE activity was markedly inhibited after the administration of EOFS in a dose-dependent manner, as observed in the donepezil-treated group (P < .05). Scopolamine treatment significantly increased AChE activity compared with the vehicle-treated group (P < .05).
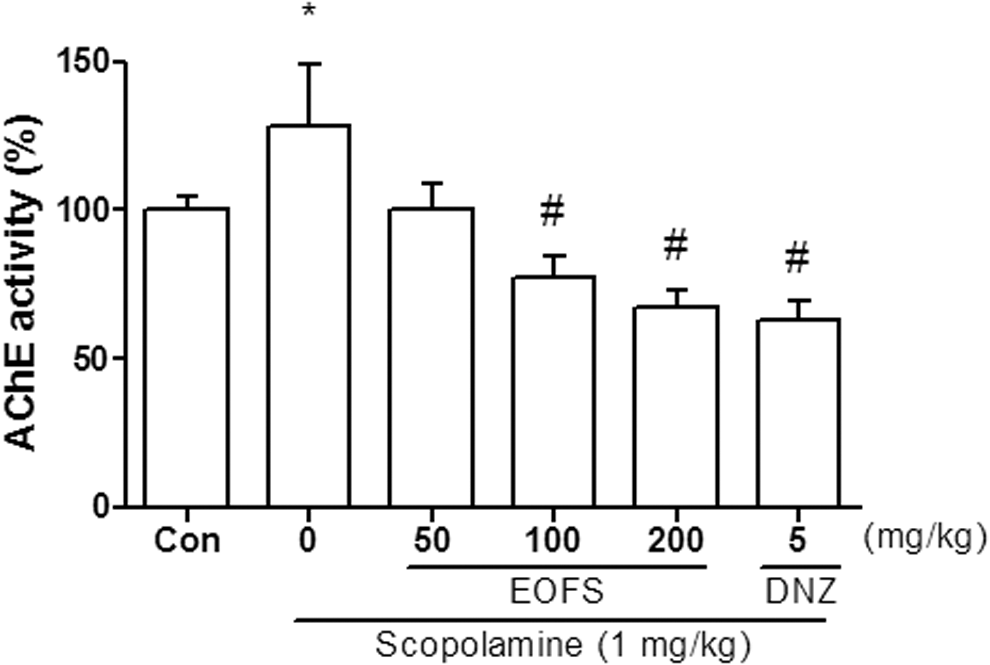
Effects of the EOFS (50, 100, and 200 mg/kg, p.o.) on AChE activity. Ex vivo AChE activity assay was performed as described in the Materials and Methods section. Control group (CON) received vehicle solution (0.9% saline). Donepezil (DNZ) was used as a positive control. Data represent the mean ± SEM (n = 4–5/group) (*P < .05, vs. the vehicle-treated control; # P < .05, vs. the scopolamine-treated group). AChE, acetylcholinesterase.
The effect of EOFS on the expression of memory-related signaling molecules in the hippocampus
We conducted western blotting to examine whether EOFS administration affected memory-related signaling molecules in the hippocampus. EOFS administration (100 or 200 mg/kg) significantly increased the levels of pERK [F(5, 23) = 5.064, P < .05], or pCREB [F(5, 18) = 9.963, P < .05], and the expression levels of BDNF [F(5, 23) = 3.734, P < .05] compared with the scopolamine-treated group (Fig. 6). However, the total unphosphorylated forms of each signaling molecule and β-actin did not change.
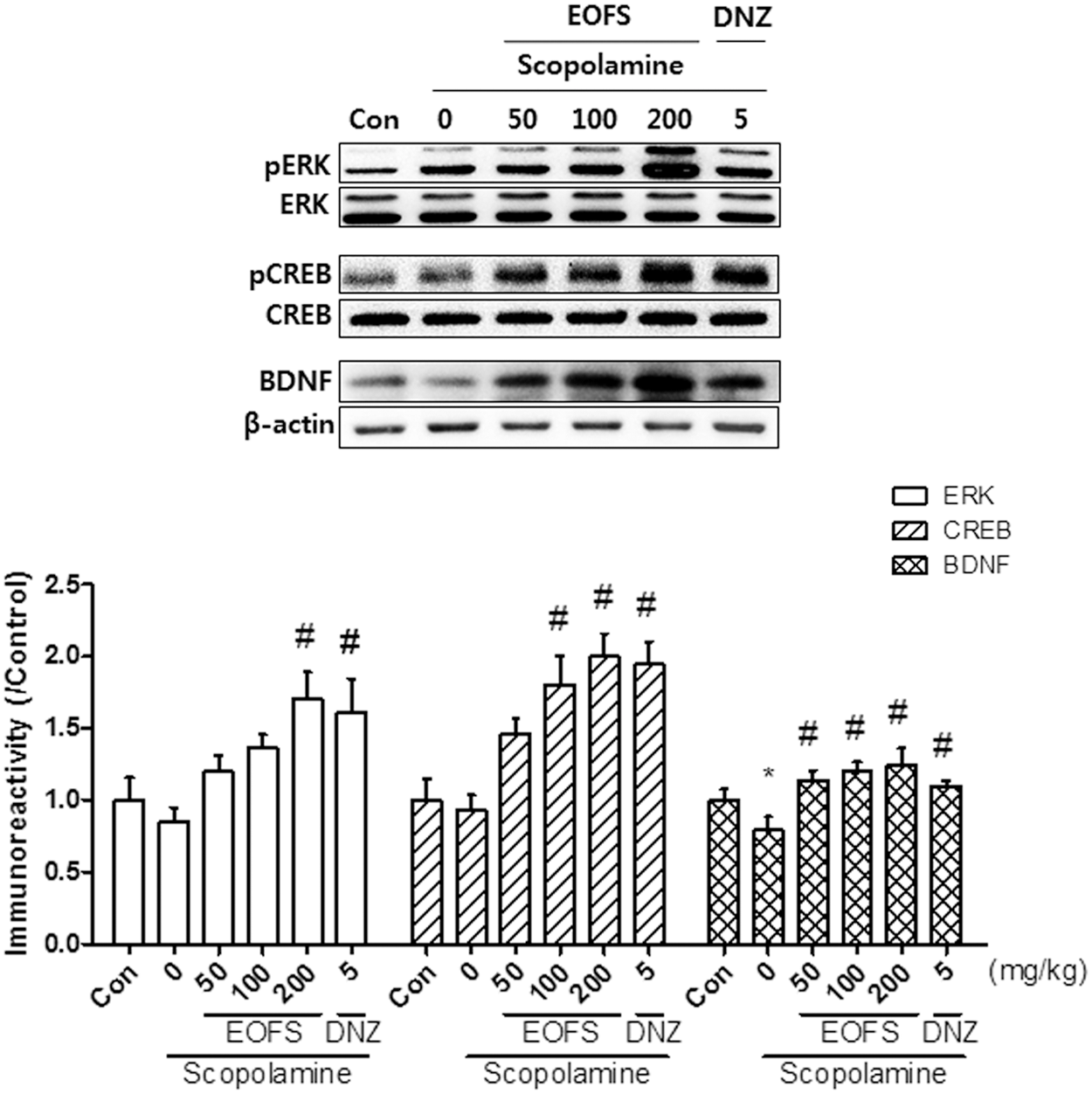
Effects of EOFS (50, 100, and 200 mg/kg, p.o.) on the expression levels of ERK, CREB, or BDNF in hippocampal tissue. Immunoreactivity and quantitative analysis of the phosphorylation levels of ERK, CREB, and BDNF expression levels are presented. Control group (CON) received vehicle solution (0.9% saline). Donepezil (DNZ) was used as a positive control. Densitometric analysis of ratios of pERK/ERK, pCREB/CREB, and BDNF/β-actin were normalized to the control group. Data represent the mean ± SEM (n = 4–5/group) (*P < .05, vs. the vehicle-treated control; # P < .05, vs. the scopolamine-treated group). BDNF, brain-derived neurotrophic factor; CREB, cAMP response element-binding protein; ERK, extracellular signal-regulated kinase.
Discussion
This study explored the effect of EOFS on scopolamine-induced cognitive impairment in mice. EOFS ameliorated scopolamine-induced cognitive dysfunction in mice as observed by the passive avoidance task, Y-maze, and novel objective recognition tasks. EOFS also markedly inhibited AChE activity in an ex vivo study and increased the levels of pERK and CREB and the expression levels of BDNF in the hippocampus.
In our previous studies, we examined some antiamnesic or antidementia agents exhibiting AChE inhibition activity from edible or herbal materials. Memory deficits in AD and mild cognitive impairment are associated with abnormal cholinergic neurotransmitter system, that is, decreased acetylcholine levels and markedly increased AChE activity. 20
In this study, we examined the effects of EOFS on the activity of AChE in mouse brain after scopolamine treatment. We found that AChE activity in scopolamine-treated group was significantly higher than that in the control group. Treatment of EOFS markedly inhibited AChE activity, and the inhibition rate was similar to that produced by donepezil. Thereafter, we investigated whether EOFS, similar to donepezil, can reverse the cognitive dysfunction associated with scopolamine-induced cholinergic blockade.
Our results show that EOFS-treated mice displayed significant increase in the step-through latency in the passive avoidance task compared with the scopolamine-treated mice. EOFS also reversed the decrease in spontaneous alternations caused by cholinergic blockade in the Y-maze task. These findings suggest that EOFS can reverse cognitive impairments by mitigating cholinergic system dysfunction. Furthermore, we examined whether EOFS can also attenuate scopolamine-induced impairment in object recognition memory in mice. Recognition memory, which is related to the integrity of the hippocampus and perirhinal cortex, 21 is commonly damaged in patients who suffer from AD. 22 Our results demonstrate that EOFS ameliorated the scopolamine-induced decreased discrimination ratio or preference for the novel object.
Taken together, our results in the ex vivo studies on AChE activity and behavioral experiments suggest that EOFS reverses the cognitive dysfunction during hypocholinergic states through AChE inhibition. On the other hand, scopolamine increased AChE activity in the brain. The mechanism behind this result is rather unclear; however, it could be speculated that scopolamine inhibited the binding of ACh to muscarinic receptors competitively and subsequently increased the accumulation of the synaptic ACh content. 23 As a result, AChE activity would be increased to catalyze the hydrolysis of ACh. 24 Further investigation is necessary to clarify this issue.
We next examined the molecular mechanisms of EOFS to further elucidate how EOFS reversed the cognitive impairments induced by scopolamine.
The cholinergic neurotransmitter system is related to the mitogen-activated protein kinase (MAPK) pathway, which is associated with cognitive performance. 25 Increased acetylcholine through EOFS-induced AChE inhibition activates muscarinic or nicotinic receptors. The muscarinic M1 receptor is the major mACh receptor in the cortex and hippocampus, and it activates MAPK signaling in mouse forebrain. 26 Nicotinic receptor is also involved in MAPK activity in the brain. 27 For example, nicotine increases the levels of pERK and CREB in the hippocampus. 20 ERK activity plays an important role in synaptic plasticity, especially during learning, and activated ERK translocates from cytoplasm into the nucleus and activates transcription factors, such as CREB. 28
The present study demonstrates that EOFS markedly increased the levels of pERK and its downstream signal molecule, CREB. The inhibition of AChE by EOFS suggests that the effect of EOFS on cognitive dysfunction involves the activation of the cholinergic neurotransmitter system, resulting in the activation of ERK–CREB signaling. Activated CREB enhances the transcription of several CREB target genes, including immediately early genes and BDNF. 29 BDNF is associated with memory integration 30,31 and LTP formation. 32 Our result also shows that EOFS administration enhanced BDNF expression level. Collectively, these findings suggest that the MAPK signaling pathway and enhanced BDNF expression in the hippocampus are responsible for the reversing effects of EOFS against the cholinergic blockade-induced cognitive impairments in our study.
O. ficus-indica var. saboten, which has a history of use as a folk medicine, exhibits neuroprotective and memory-enhancing activities, 15,33 which supports its use as a potential agent for the treatment of cognitive impairment associated with neurodegenerative diseases. EOFS ameliorated scopolamine-induced cholinergic dysfunction in the present study. These effects were derived from AChE inhibition and activation of the ERK–CREB–BDNF signaling pathway. Collectively, the present study suggests the use of EOFS as a promising therapeutic candidate for the treatment of cognitive dysfunctions observed in AD patients.
Footnotes
Acknowledgments
This work was supported by the Seoul Research and Business Development Program (CI152006). The authors thank Chrislean Jun Botanas (College of Pharmacy, Shamyook University, Republic of Korea) for his editorial input in the article.
Author Disclosure Statement
No competing financial interests exist.