
Abstract
We have previously reported the lipid-lowering effects of a Korean rice cookie called dasik (RCD) in comparison with a western style cookie. In this study, Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine) fruit-supplemented RCD (SRCD) was added to a diet, and the hypolipidemic and antidiabetic effects of different diets were examined by using the ICR and db/db mouse models, respectively. ICR mice were fed the AIN-76 diet, or high-fat diet (HFD), or the RCD- or SRCD-supplemented HFD (10%, w/w) for 9 weeks (n = 7 per group). Compared with the RCD group, plasma and hepatic triglyceride and cholesterol concentrations were decreased in the SRCD group. Hepatic expressions for fatty acid and cholesterol synthesis were downregulated, whereas those for beta-oxidation and cholesterol export were upregulated (P < .05). The antidiabetic effects of SRCD were tested in db/db mice for 10 weeks (n = 7 per group). Glucose tolerance was improved in the SRCD group through the regulation of gluconeogenic enzymes and biomarkers related to the insulin signaling pathway (P < .05). In addition, SRCD increased the expression levels of antioxidative enzymes, and decreased those of inflammatory cytokines (P < .05). Moreover, oxidative stress, leptin, and insulin levels were lower in the SRCD group than in the other groups (P < .05). In conclusion, the lipid-lowering and antidiabetic effects of SRCD were greater than those of RCD with respect to the suppression of lipid synthesis, oxidative stress, and inflammation and the improvement of glucose metabolism.
Introduction
T
The control of hyperlipidemia is essential because it is a primary cause of T2D development. Plasma lipid levels are controlled by hepatic lipid homeostasis, which is regulated by several transcription factors, including sterol regulatory element-binding protein-1 and -2 (SREBP-1 and -2), peroxisome proliferator-activated receptor-alpha (PPAR-α), and cytochrome P450 7A1 (CYP7A1). SREBP-1 upregulates lipogenic enzymes, such as fatty acid synthase (FAS) and acetyl-CoA carboxylase (ACC). PPAR-α transcribes lipolytic enzymes, such as carnitine palmitoyltransferase 1 (CPT1) and acetyl-CoA oxidase 1 (ACOX1). SREBP-2 initiates cholesterol synthesis through upregulation of 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR), whereas CYP7A1 controls the conversion of cholesterol into bile acids.
Insulin signaling plays an important role in glucose oxidation, which is mediated through the insulin receptor substrate (IRS)/phosphoinositide 3-kinase (PI3K)/Akt pathway. 5 In addition, AMP-activated protein kinase (AMPK) also participates in the regulation of glucose metabolism and insulin resistance in T2D 6 by promoting glucose utilization and inhibiting lipogenesis through ACC and HMGCR downregulation. 7 Thus, natural products that activate the IRS/PI3K/AKT and/or AMPK pathways to cause an elevation in glucose uptake and a reduction in insulin resistance and lipogenesis would have beneficial effects on T2D. 4
Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine) fruit (SF) (five-flavor fruit, omija in Korean) has been used for the treatment of cancer in Chinese traditional medicine. The pharmacological components of SF are dibenzocyclooctadiene lignans 8 and polyphenolic compounds, such as anthocyanins and flavonols. 9 SF and its active compounds have revealed antidiabetic, 10 antioxidative, 9 antibacterial, 11 anti-inflammatory, 12 and lipid-lowering effects. 13 The food industry has focused on the use of sugar alternatives to reduce the adverse effects of sugar. Fructooligosaccharides (FOSs) have fewer calories than sugar and have demonstrated prebiotic effects. Numerous studies have shown health benefits of FOSs, including their anti-inflammatory, 14 antidiabetic, 15 and lipid-lowering effects. 16
Daily snacking can result in a sharp increase in energy intake, which can reach up to 25% of the total caloric intake in modern society. The Korean rice cookie called dasik (RCD) is considered a prudent snack because its ingredients are of plant origin. In our previous study, the plasma and hepatic lipid-lowering effects of RCD made with rice flour, mungbean, and honey were higher than those of a traditional western style cookie. 17 Moreover, the plasma triglyceride level was shown to be significantly reduced in middle-aged women who consumed a different kind of RCD made with brown rice, red ginseng, and FOS compared with a traditional western style cookie. 18
Agents that possess hypolipidemic, antioxidative, and/or anti-inflammatory activities might improve the impaired glucose metabolism and insulin intolerance of diabetes. 19 In this study, basic RCD ingredients supplemented with SF and FOS were added to a diet and their mechanisms of action in lowering lipid levels and on the insulin signaling pathway were examined in both high-fat diet (HFD)-fed ICR and db/db mice.
Materials and Methods
Ingredients of the rice cookies
The RCD ingredients were rice cake flour (55.6%, Imgeumnimpyo; Seulsung Nonghyup, Icheon, Gyeonggi, Korea), mung bean starch (22.2%; Emart Co., Seoul, Korea), and honey (22.2%; Dongsuh Food, Seoul, Korea). The SF-supplemented RCD (SRCD) ingredients were rice cake flour (47.6%), mung bean starch (19.0%), FOS (19.0%; CJ Cheiljedang Co., Seoul, Korea), SF extract (omija juice, 14.3%; Moonkyungfarm Co., Moonkyung, Gyeongbuk, Korea), and propolis (0.1%; Withealth Co., Geochang Gun, Gyeongnam, Korea).
Experimental diets and animal care for ICR mice
The lipid-lowering effects and mechanism of action of SRCD were elucidated by using the ICR mouse. First, to prepare the HFD, lard (21.7%, w/w) and cholesterol (0.4%, w/w) were added to the AIN-76 synthetic diet (normal diet, ND). Subsequently, each ingredient of RCD or SRCD was added directly to the HFD to make up an isocaloric experimental diet of 5.0 kcal/g diet. The ND contained 3.9 kcal/g diet (Table 1). The calories provided from the rice cookie were 7% of the total calories of the experimental diet.
NOR, ICR mice fed the AIN-76 diet; HFD, ICR mice fed the HFD only; HFD-RCD, ICR mice fed the HFD supplemented with the Korean RCD ingredients; HFD-SRCD, ICR mice fed the HFD supplemented with the RCD ingredients with Schisandra chinensis (Turcz.) Baill. fruit (SF) added. All diets were given for 9 weeks.
RCD, db/db mice fed the AIN-76 diet supplemented with the RCD ingredients; SRCD, db/db mice fed the AIN-76 diet supplemented with the SRCD ingredients. All diets were given for 10 weeks.
FOS, fructooligosaccharide; HFD, high-fat diet; RCD, rice cookie called dasik; SRCD, SF-supplemented rice cookie called dasik.
Male ICR mice (4 weeks old) were purchased from Orient Bio, Inc. (Seongnam, Korea). After 1 week of adaptation, the mice were assigned to four groups (n = 7 per group) based on body weight, and they were raised for 9 weeks. The four groups of mice were fed the ND (NOR group), HFD-only (HFD group), RCD-supplemented HFD (RCD group), and SRCD-supplemented HFD (SRCD group). The mice were kept in individual cages during the entire experimental period under a controlled atmosphere (room temperature, 23°C ± 1°C; humidity, 50% ± 5%; 12 h light-dark cycle). The dietary intake was checked daily, and the body weight was measured every week. After 9 weeks of feeding, all mice were fasted for 12 h and anesthetized by intraperitoneal administration of 30 mg/kg of Zoletil 50 (Virbac Laboratories, Carros, France) and 10 mg/kg of xylazine (Rompun; Bayer Korea, Seoul, Korea). Blood and liver was excised and stored at −80°C. The animal study was approved by the Pusan National University Institutional Animal Care and Use Committee (PNU-IACUC, Approval No. PNU-2012-0118).
Experimental diets and animal care for db/db mice
The antidiabetic effects of SRCD in comparison with those of RCD were examined in the db/db mouse model. For preparation of the experimental diet, each ingredient of RCD or SRCD was directly added to the AIN-93 diet (10%, w/w) to make up isocaloric diets of 3.9 kcal/g diet (Table 1).
Male C57BLKS/J lar- +Leprdb/+Leprdb mice (db/db mice, 6 weeks old) were purchased from SLC, Inc. (Hamamatsu, Japan). After 1 week of acclimatization, the mice were assigned to either the RCD group or the SRCD group based on body weight and raised for 10 weeks. The care of the db/db mice and the procedures for plasma and liver sample collection were carried out in the same manner as for the ICR mice. The animal study was approved by PNU-IACUC (Approval No. PNU-2012-0124).
Analysis of biochemical parameters
The plasma aminotransferase (AM101-K) and lipid levels such as triacylglycerol (TG, AM157S-K) and total cholesterol (TC, AM202-K) were measured by using commercially available kits (Asan Pharmaceuticals, Seoul, Korea). The plasma insulin (80-INSMS-E01; ALPCO Diagnostics, Salem, NH, USA) and leptin (ADI-900-019A; Enzo Life Sciences AG, Lausen, Switzerland) levels were measured by using ELISA kits. The hepatic lipids were extracted from the liver tissue by using a modified method of Folch. 20 The hepatic TG and TC concentrations were measured with the same kits used for the plasma lipid analyses.
Oxidative stress-related parameters
Thiobarbituric acid reactive substances (TBARS) 21 and glutathione (GSH) 22 were determined by using malondialdehyde and GSH standards, respectively. For assay of reactive oxygen species (ROS) and peroxynitrite, a post-mitochondrial fraction was prepared by two separate centrifugations of the liver homogenate (3,012 × g, followed by 18,627 × g at 4°C for 20 min) and determined by using 2′,7′-dichlorofluorescein-diacetate 23 and dihydrorhodamine 123 buffer, 24 respectively.
Oral glucose tolerance tests
On the last day of experimental feeding, an oral glucose tolerance test (OGTT) was performed on the db/db mice after an overnight fasting period of 16 h. After an oral administration of glucose (0.1 g/100 g body weight), blood was drawn from the tail at 0, 30, 60, and 120 min and the glucose concentration was determined by using a glucometer (Roche Diagnostics GmbH, Mannheim, Germany).
Western blot analysis
The western blot assay was performed as previously described. 25 Protein expression was first visualized by enhanced chemiluminescence, detected by using a CAS-400 system (Core Bio, Seoul, Korea), and finally evaluated by ImageJ software (National Institutes of Health, Bethesda, MD, USA). The protein expression level was standardized to that of α-tubulin or β-actin.
The primary antibodies used for detecting proteins of lipid metabolism were as follows: β-actin (ab8226) and FAS (ab22759), purchased from Abcam, Inc. (Cambridge, UK); and SREBP-1 (sc-8984), ACCα (sc-26817), PPAR-α (sc-9000), CPT1 (sc-139482), ACOX1 (sc-98499), SREBP-2 (sc-5603), HMGCR (sc-33827), and CYP7A1 (sc-25536), all from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
The primary antibodies used for the diabetes study were as follows: pyruvate carboxykinase (PCK, ab70358), purchased from Abcam, Inc.; phospho-AMPKα (pAMPK, #2535), phospho-IRS-1 (pIRS, #2385), phospho-PI3K (pPI3K, #4228), and phospho-Akt (pAkt, #9271), purchased from Cell Signaling Technology (Beverly, MA, USA); and glucose-6-phosphatase (G6pase, sc-25840), nuclear factor-like 2 (Nrf2, sc-13032), kelch-like ECH-associated protein 1 (Keap1, sc-15246), heme oxygenase-1 (HO-1, sc-10789), GSH S-transferase (GST, sc-138), nuclear factor kappaB (NF-κB, sc-109), cyclooxygenase-2 (COX-2, sc-1747), inhibitor of NF-κB (IκB, sc-371), and inducible nitric oxide synthase (iNOS, sc-651), all from Santa Cruz Biotechnology.
Statistical analysis
Data are presented as the mean ± standard deviation. Statistical analysis was performed by using SPSS Version 23 (SPSS, Inc., Chicago, IL, USA). For the study of lipid metabolism, the statistical significance of differences among the four groups was assessed by one-way analysis of variance (ANOVA), followed by Duncan's multiple-range test. For the db/db mouse model, data differences between the RCD and SRCD groups were analyzed by Student's t-test. Significance was considered at P < .05.
Results
Food intake, body weight gain, and aminotransferase activity in HFD-fed ICR mice
Food intakes among the experimental groups were not different (Table 2). Compared with the NOR group, the HFD-induced body weight gain and epididymal fat mass increases were significantly decreased in both rice cookie groups (P < .05). The plasma alanine transaminase and aspartate transaminase levels of the RCD and SRCD groups were lower than those of the HFD group (P < .05).
Data are the mean ± SD (n = 7 per group). a–cData with different letters in the row are significantly different according to one-way ANOVA followed by Duncan's multiple-range test at P < .05 in comparison to ICR mice. NSNot significantly different at P < .05.
See the footnote text of Table 1 for a detailed description of the experimental groups.
ALT, alanine transaminase; ANOVA, analysis of variance; AST, aspartate transaminase; SD, standard deviation.
Plasma and hepatic lipid concentrations in HFD-fed ICR mice
The HFD significantly elevated the plasma TG and TC levels, but those were reduced in both rice cookie groups (Table 3, P < .05). The plasma TG and TC levels in the SRCD group were significantly lower (by 35.18% and 2.48%, respectively) than the RCD group. Similarly, hepatic TG and TC levels in the SRCD group were significantly lower (by 30.24% and 13.51%, respectively) than those in the RCD group.
Data are the mean ± SD (n = 7 per group).
Data with different letters in the row are significantly different according to one-way ANOVA followed by Duncan's multiple-range test at P < .05 in comparison to ICR mice. *Significant differences are expressed as P-values calculated by Student's t-test in the comparisons for db/db mice (* P < .05, *** P < .001). NSNot significantly different at P < .05.
See the footnote text of Table 1 for a detailed description of the experimental groups.
TC, total cholesterol; TG, triacylglycerol.
Protein expression of lipid metabolism-related markers in the HFD-fed ICR mice
Rice cookie supplementation suppressed lipogenesis but augmented fatty acid oxidation in the HFD-fed ICR mice. The expression levels of SREBP-1 (mature form), FAS, and ACCα in the SRCD group were significantly decreased by 38.67%, 31.56%, and 25.32%, respectively, relative to the levels in the RCD group (Fig. 1, P < .05). In contrast, the expression levels of PPAR-α, CPT1, and ACOX1 in the SRCD group were significantly higher (by 137.41%, 131.88%, and 131.85%, respectively) than the levels in the RCD group.
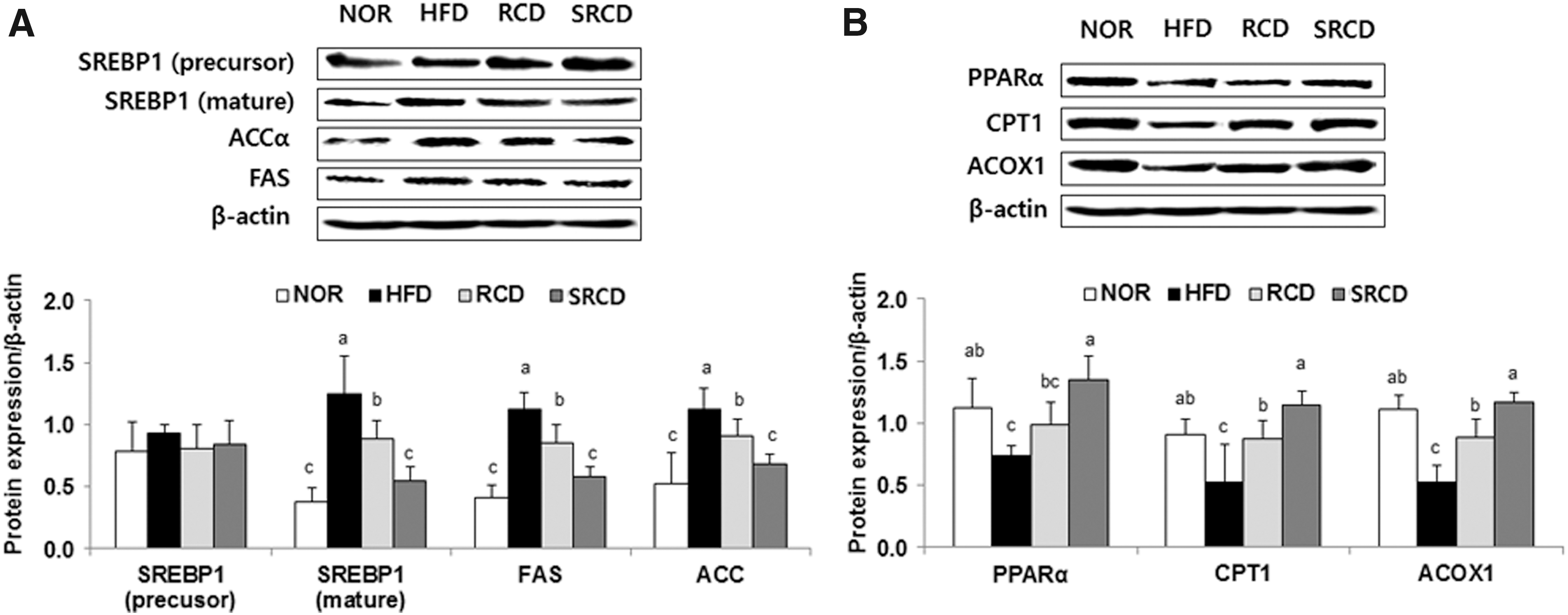
Hepatic protein expression involved in fatty acid synthesis
The expression levels of SREBP-2 (mature form) and HMGCR in the SRCD group were significantly lower (by 101.86% and 98.25%, respectively) than the levels in the RCD group (Fig. 2, P < .05). On the other hand, the expression of CYP7A1 in the SRCD group was significantly higher (by 146.40%) than that in the HFD group.
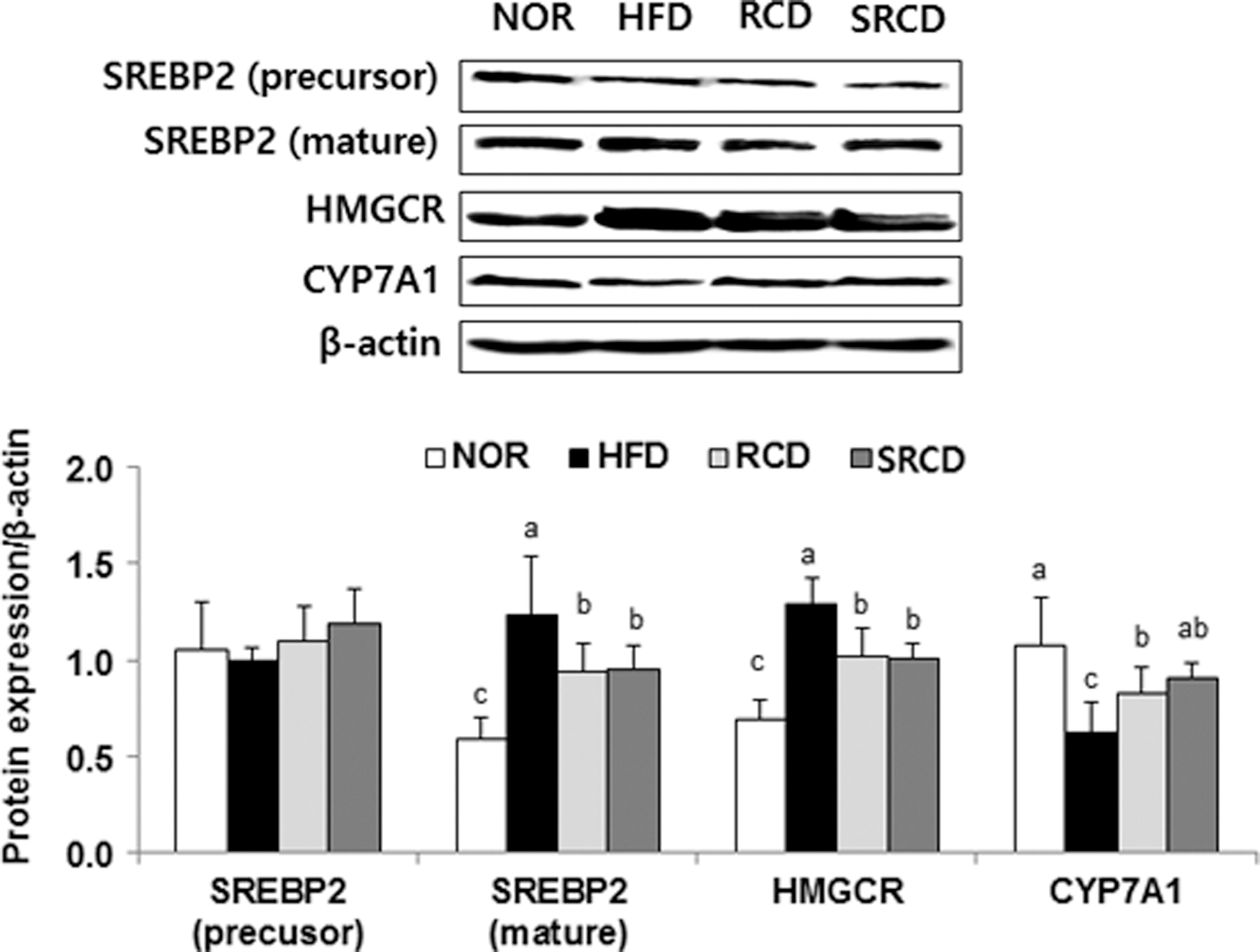
Hepatic protein expression involved in cholesterol synthesis and catabolism in HFD-fed ICR mice. See the footnote text of Table 1 for a description of the experimental groups. a–cData with different letters in the column are significantly different according to one-way ANOVA followed by Duncan's multiple-range test at P < .05.
Plasma and hepatic lipid concentrations in db/db mice
Plasma and hepatic TG concentrations in the SRCD group were lower (by 12.95% and 40.11%, respectively) than the levels in the RCD group (Table 3, P < .05). However, the plasma and hepatic TC concentrations were not significantly different between the two groups.
Glucose tolerance and leptin and insulin levels in db/db mice
The area under the curve (AUC) value from the OGTT of the SRCD group was significantly lower than that of the RCD group by 16.04% (Fig. 3, P < .05), indicating that glucose tolerance was improved in the SRCD group. The leptin and insulin levels in the SRCD group were lower (by 48.36% and 50.64%, respectively).
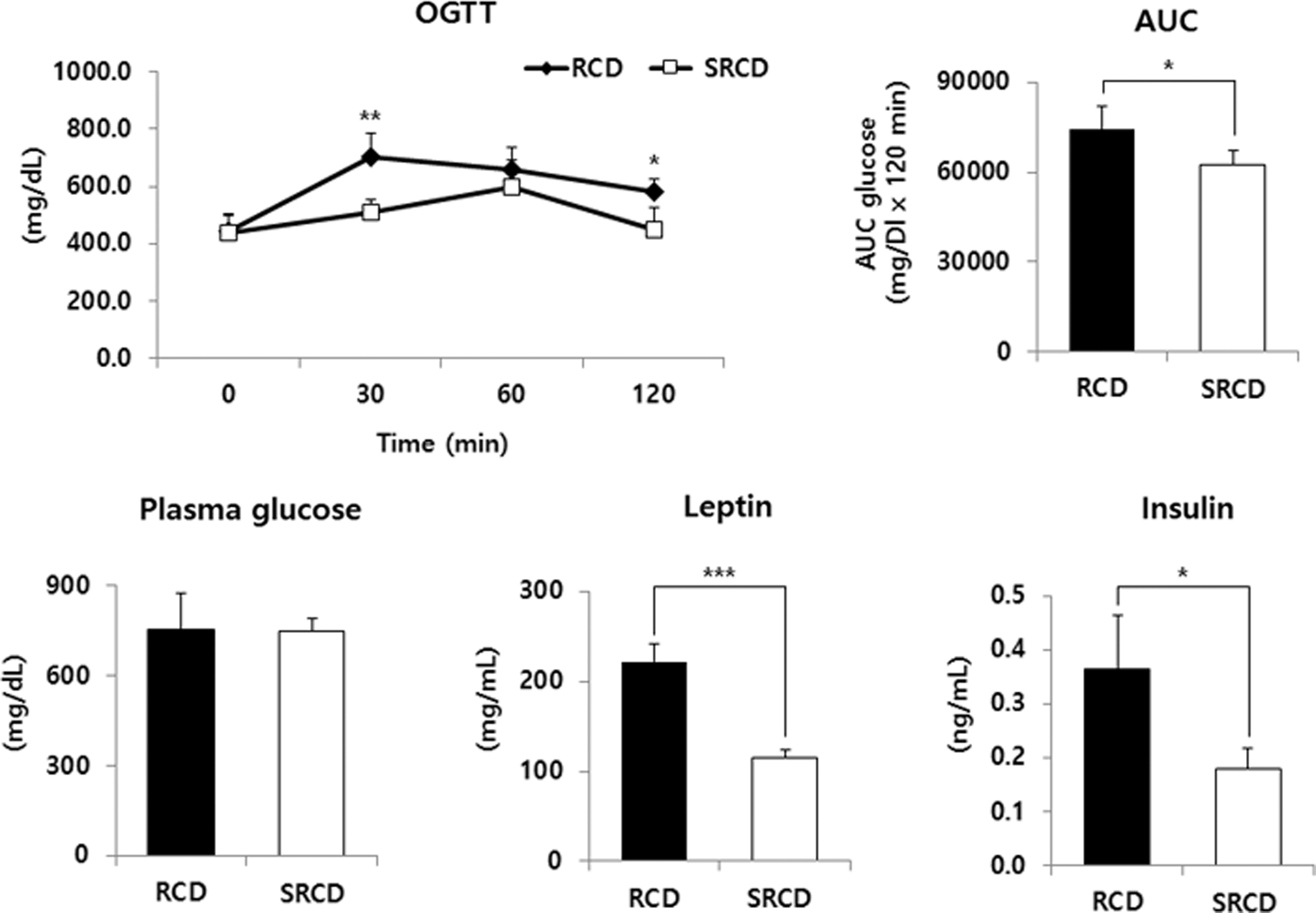
OGTT, glucose AUC, plasma glucose, leptin, and insulin levels in db/db mice. See the footnote text of Table 1 for a description of the experimental groups. *Significant differences between two experimental groups are expressed as P-values calculated by Student's t-test (*P < .05, **P < .01, and ***P < .001). AUC, area under the curve; OGTT, oral glucose tolerance test.
Lipid peroxidation and oxidative stress levels in db/db mice
Compared with the plasma ROS and TBARS levels in the RCD group, the respective levels in the SRCD group were significantly decreased by 77.59% and 15.70%, respectively (Table 4, P < .05). The hepatic peroxynitrite and TBARS levels in the SRCD group were lower (by 48.86% and 33.42%, respectively). In contrast, the hepatic GSH level of the SRCD group was increased by 256.91% (P < .05).
Data are the mean ± SD (n = 7 per group). *Significant differences are expressed as P-values calculated by Student's t-test in the comparisons for db/db mice (* P < .05, ** P < .01). NSNot significantly different at P < .05.
See the footnote text of Table 1 for a detailed description of the experimental groups.
GSH, glutathione; MDA, malondialdehyde; ONOO−, peroxynitrite; ROS, reactive oxygen species; TBARS, thiobarbituric acid reactive substances.
Protein expression of glucose metabolism-related biomarkers in db/db mice
Compared with the protein expression levels of pAMPK, pIRS, pPI3K, and pAkt in the RCD group, the levels in the SRCD group were significantly increased by 119.85%, 142.79%, 144.30%, and 161.71%, respectively, and that of G6pase was decreased by 26.44% (Fig. 4, P < .05). The PCK expression levels were not significantly different between the two groups.
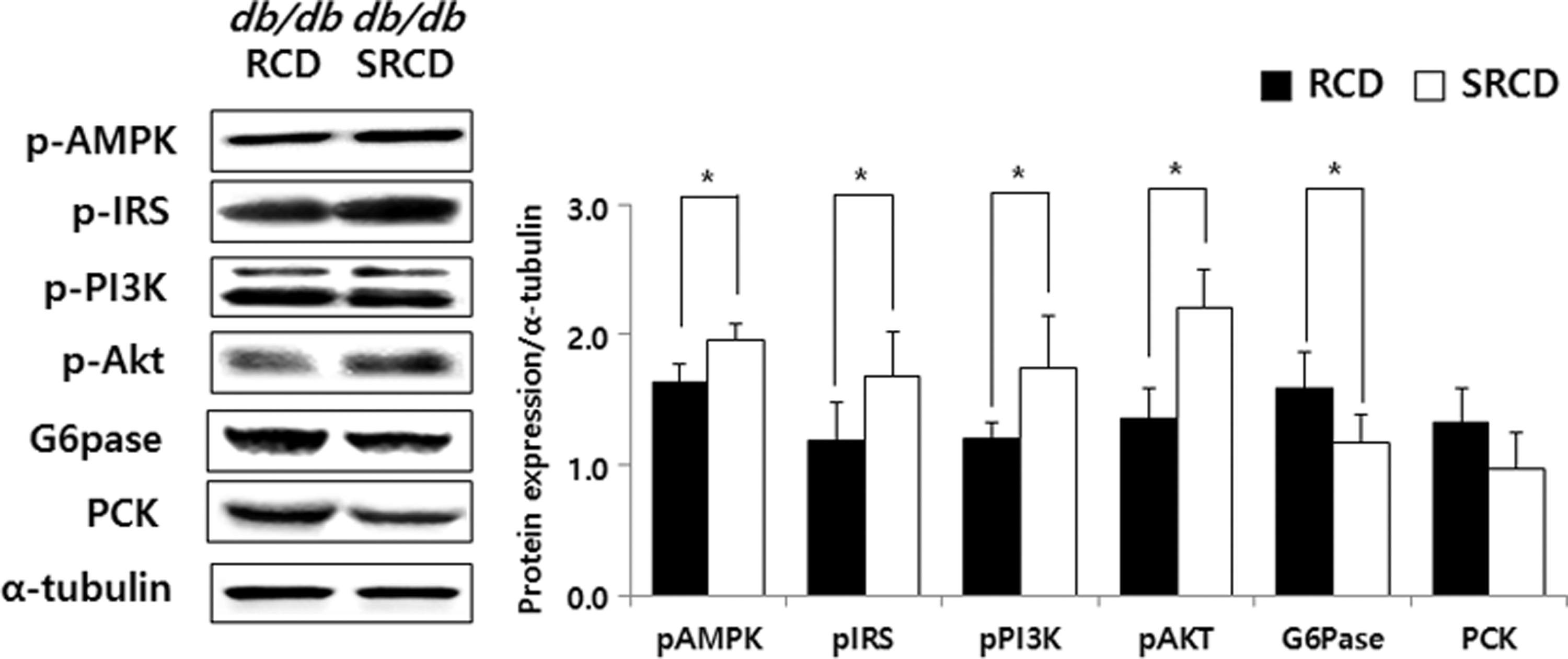
Hepatic protein expression involved in gluconeogenesis and insulin signaling pathway in db/db mice. See the footnote text of Table 1 for a description of the experimental groups. *Significant differences between two experimental groups are expressed as P-values calculated by Student's t-test (*P < .05).
Protein expression of antioxidative and anti-inflammatory biomarkers in db/db mice
The Nrf2, HO-1, and GST expression levels in the SRCD group were 129.83%, 135.62%, and 144.97%, respectively, higher than those in the RCD group (Fig. 5, P < .05). The expression levels of NF-кB, COX-2, and iNOS in the SRCD group were lower (by 52.27%, 29.71%, and 44.22%, respectively) (Fig. 6, P < .05). However, the Keap1 and IкB expression levels were not significantly different between the two groups.
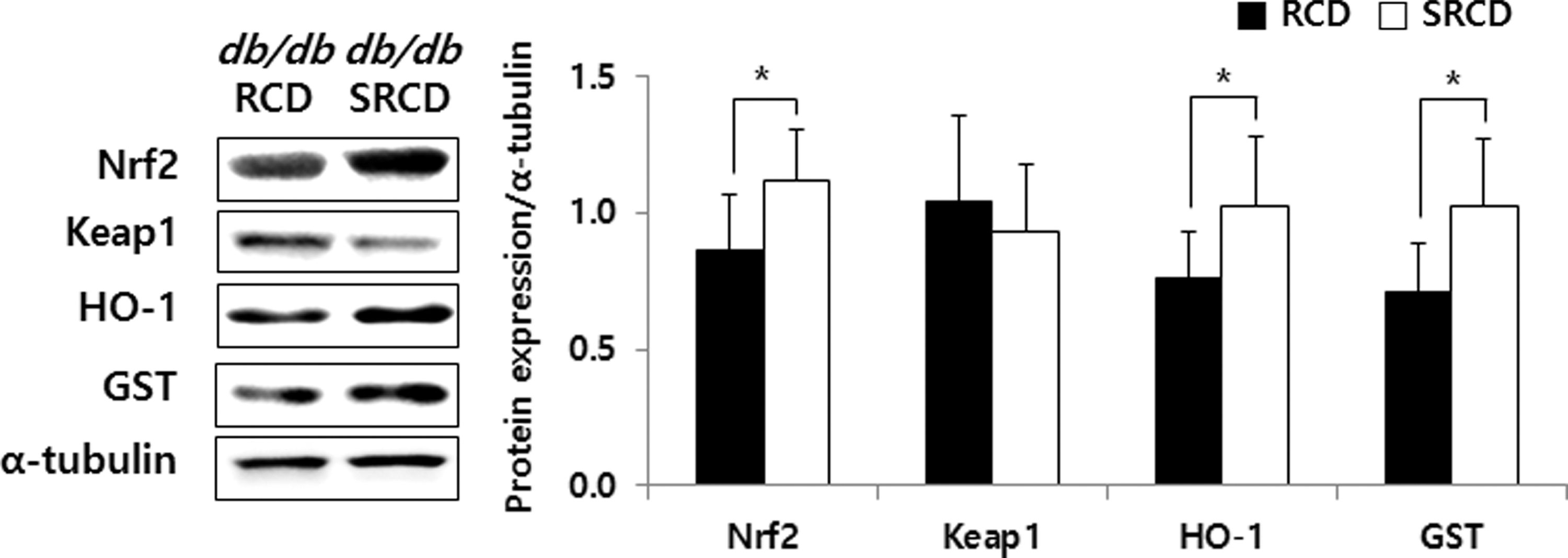
Hepatic protein expression of antioxidants in db/db mice. See the footnote text of Table 1 for a description of the experimental groups. *Significant differences between two experimental groups are expressed as P-values calculated by Student's t-test (*P < .05).
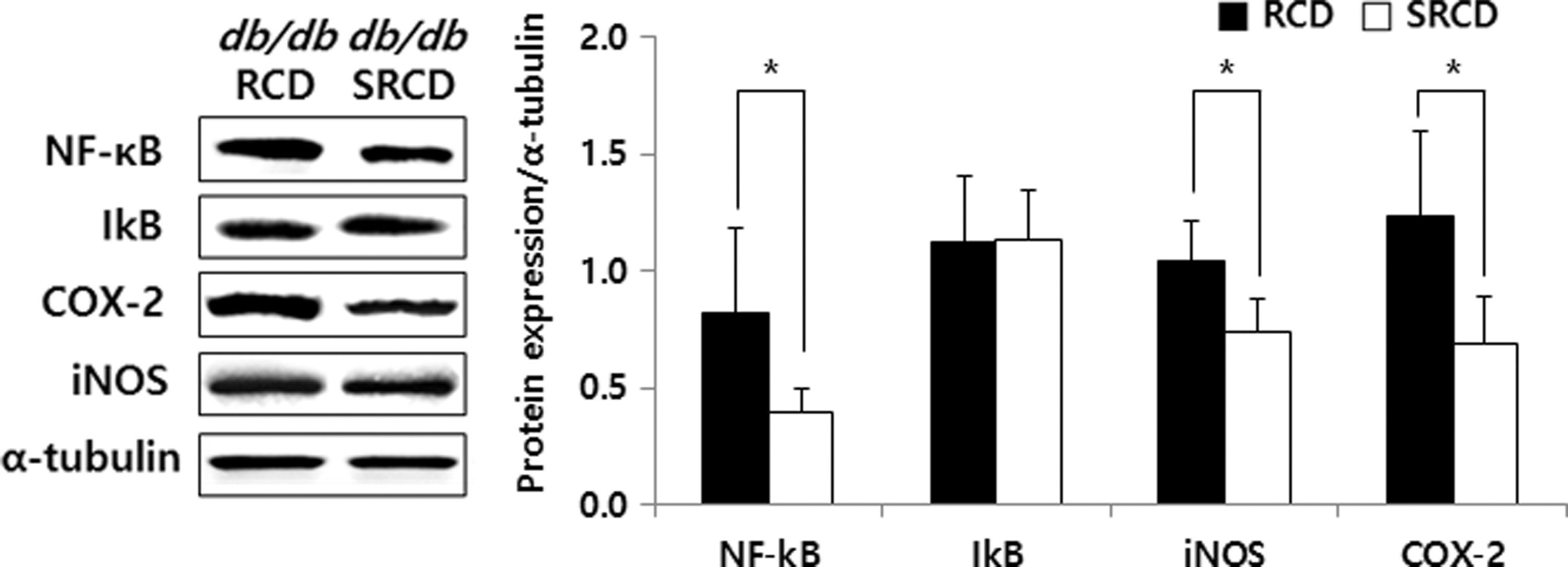
Hepatic protein expression of inflammatory cytokines in db/db mice. See the footnote text of Table 1 for a description of the experimental groups. *Significant differences between two experimental groups are expressed as P-values calculated by Student's t-test (*P < .05).
Discussion
Aside from good control of blood glucose levels, regulation of the plasma lipid concentration in T2D is an important issue because obesity is strongly associated with the development of this disease. Snacking on sweet foods is irresistible to people because of the comforting sensation elicited by the sweetness taste. However, table sugar and high-sugar foods are the least recommended food items to the diabetic patient. Stevia and FOS, which have a sweet taste, but few no or less calories than sugar, have drawn attention for efforts to cut down the calories in snacks. SF (omija) has demonstrated antidiabetic, 10 antioxidative, 9 antibacterial, 11 anti-inflammatory, 12 and lipid-lowering effects. 13 SF extracts reduced the serum and hepatic TG and TC levels in HFD-fed rats by upregulating lipolytic genes, including PPAR-α and CPT1, and downregulating the ACC, FAS, and HMGCR genes regulated by SREBP-1 and -2. 10 In addition, SF extracts improved hepatic steatosis and dyslipidemia in HFD-fed mice by decreasing plasma and hepatic TG, TC, and free fatty acid levels and increasing fecal lipid excretion. 13 These lipid-lowering effects of SF cited by other researchers are consistent with our current study, wherein the mechanism of action of SRCD in reducing the plasma and hepatic TG concentrations was found to be mediated through downregulation of the SREBP-1-regulated FAS with concomitant upregulation of the PPAR-α-regulated ACOX1 and CPT1. Similarly, the cholesterol levels in the SRCD group were decreased through downregulation of SREBP-2 and HMGCR, and upregulation of CYP7A1. These results indicate that lipogenesis was suppressed but fatty acid oxidation or excretion was increased. Moreover, the lipid-lowering effects observed in the SRCD group might be attributed, in part, to the replacement of honey with FOS. In another study, 2.5% (w/w) FOS supplementation decreased the plasma and hepatic lipid levels in HFD-fed mice, leading to a reduction in the epididymal fat mass. 15 In addition, 5.0% (w/w) FOS supplementation to an ND decreased the hepatic lipid levels in obese rats. 26 In our study, the FOS content in the SRCD recipe was ∼19%.
Glucose uptake is dependent on the insulin signaling initiated by IRS phosphorylation, which accordingly activates PI3K and Akt. Under this condition, gluconeogenesis is suppressed via the suppression of PCK and G6pase. 1 In addition, AMPK, another regulator of lipid and glucose metabolism, stimulates insulin secretion with the concomitant deactivation of gluconeogenic enzymes such as PCK and G6pase. Moreover, AMPK downregulates ACC and HMGCR, 6,7 thereby decreasing lipogenesis as well. It is apparent that AMPK plays an important role in the diabetes-related metabolic syndrome, which is attributed to its regulatory activities on insulin signaling and lipid and glucose metabolism. In this study, the expression levels of pAMPK, pIRS, pPI3K, and pAkt in the SRCD group were increased and that of G6pase was decreased, suggesting that the insulin sensitivity and glucose utilization for energy production were increased in the SRCD group. As a result, the leptin and insulin levels in the SRCD group were decreased. Previous studies have reported that SF extract alleviated insulin resistance in HFD-fed hamsters by improving oral glucose tolerance and decreasing the glucose AUC and insulin levels. 9 SF also exhibited beneficial effects on the plasma glucose and insulin levels and on hepatic PCK and G6pase activities in HFD-fed mice. 13 The lignan-rich fractions in SF were recognized as the active principles for upregulating pAMPK and pAkt in the liver of diabetic rats. 27 In addition, FOSs exerted a serum insulin-lowering effect in obese rats. 26 Therefore, taken together, the antidiabetic effects in the SRCD group observed in our current study might be because of SF and FOS.
Hyperlipidemia and hyperglycemia cause cellular damage by elevating oxidative stress and increased production of inflammatory cytokines, which accelerate lipid peroxidation. 28 In contrast, antioxidants have been shown to alleviate obesity by decreasing body fats through the suppression of inflammatory responses. 29 In this study, SRCD decreased the plasma and hepatic oxidative stress levels in both the HFD-fed mice and db/db mice. Moreover, in db/db mice, SRCD increased the hepatic Nrf2-induced antioxidants, and decreased the NF-κB-induced inflammatory cytokines. Schisandrin, the active component of SF, increased the expression of Nrf2 and HO-1 in RAW 264.7 cells 30 and decreased the expression of NF-κB, COX-2, and iNOS in LPS-treated macrophage cells. 12 These results are in good agreement with our current findings on the effects of SRCD.
The mechanism that we have examined in this study is shown in Figure 7. In summary, SRCD significantly attenuated the obesity-associated diabetic state by inhibiting lipid synthesis and promoting fatty acid oxidation in HFD-fed mice. In addition, SRCD improved insulin sensitivity and glucose metabolism in db/db mice, mediated through elevation of the antioxidant status and alleviation of the inflammatory reaction. In conclusion, a functional rice cookie supplemented with SF and FOS would ameliorate the pathophysiological conditions of obesity-associated diabetes in individuals who cannot restrain themselves from snacking.
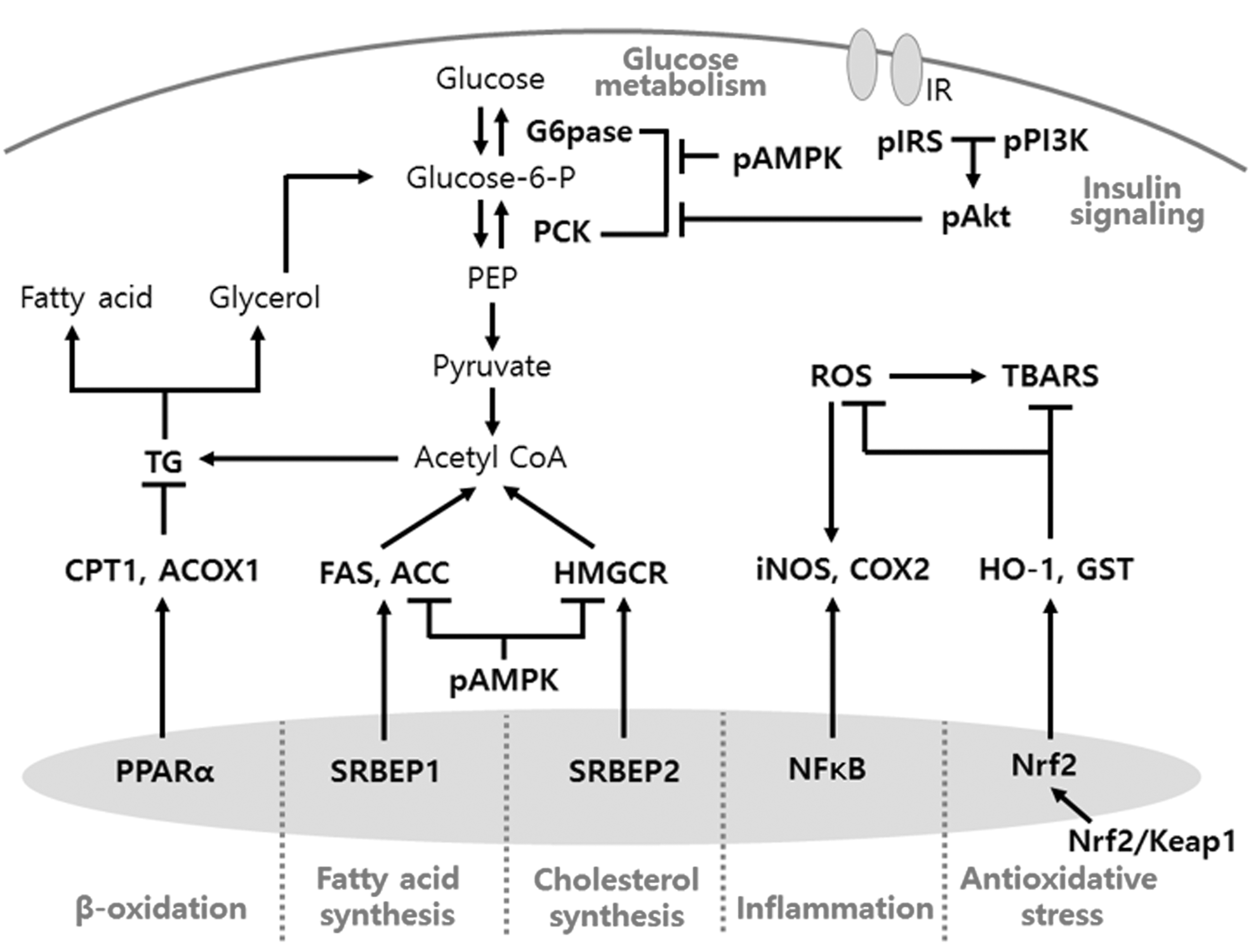
Schematic of the mechanism elucidated in this study.
Footnotes
Acknowledgments
This research was supported by the Superiority and Functionality of Hansik (Korean Food) Research Program (No. 911040-1), iPET (Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries), and Ministry of Agriculture, Food and Rural Affairs.
Author Disclosure Statement
No competing financial interests exist.