
Abstract
Obesity is the most common metabolic disease in developed countries and has become a global epidemic in recent years. Obesity is associated with various metabolic abnormalities, including glucose intolerance, insulin resistance, type 2 diabetes, dyslipidemia, and hypertension. Leaves from the plant Dendropanax morbiferus are beneficial to health as they contain high levels of vitamin C and tannin. There have been seminal studies on the anticancer, antimicrobial, antidiabetes, and antihyperglycemic effects of treatments with D. morbiferus trees. Herein, we investigated the toxicity of D. morbiferus water (DLW) extracts in vitro, and demonstrated no toxicity at 5–500 μg/mL in 24–72-h experiments with 3T3-L1 cells. The DLW increased cell viability at 48 h and inhibited adipogenesis in 3T3-L1 cells by reducing intracellular triglyceride levels and glucose uptake. In addition, mRNA and protein expression levels of adipogenesis-related genes were lowered by DLW, suggesting antiobesity effects in mouse 3T3-L1 cells. Because few studies have demonstrated cholesterol-lowering effects of D. morbiferus, we investigated the activities of adipogenic transcriptional factors following treatments of 3T3-L1 cells with D. morbiferus and observed increased CEBPα, CEBPβ, PPARγ, and SREBP1 activities in the cells, indicating that DLW extracts inhibit adipogenesis.
Introduction
D
D. morbiferus leaf extracts also have excellent cytoprotective and skin whitening effects, suggesting the presence of inhibitors of tyrosinase and melanin formation. In addition, recent studies show anticancer effect, antimicrobial activities, and benefits for patients with diabetes and hyperglycemia. 3,4 D. morbiferus contains various physiologically active substances and exhibits antioxidant effects against oxidative stress. A few studies have reported improvement in lipid metabolism following treatments with D. morbiferus. 5 –8
Obesity is an increasingly common cause of metabolic disease in developed countries, and its prevalence has warranted consideration as a global epidemic in recent years. Obesity is characterized by excessive accumulation of body fat due to an imbalance between energy intake and consumption, and is associated with chronic diseases, including glucose intolerance, insulin resistance, type 2 diabetes, dyslipidemia, and hypertension. These metabolic disorders have become a major health issue for adults and children in modern society. 9 –12 Recent research efforts have been designed to test the hypothesis that excessive weight gain and obesity can be ameliorated by inhibiting adipogenesis and lipogenesis. 13 –15
Accumulation of body fat has been correlated with increased adipocyte numbers and sizes, and various dietary factors have been shown to affect numbers of adipocytes (hyperplasia) and adipocyte sizes (hypertrophy). Hence, we examined antiobesity and cholesterol-lowering effects of D. morbiferus leaf extracts and correlated these with lipid peroxidation levels.
Materials and Methods
Cell culture
293T, HepG2, and 3T3-L1 cells were cultured in Dulbecco's modified Eagle's medium (DMEM, Hyclone, USA) containing 10% fetal bovine serum (FBS, Hyclone) and 1% antibiotics (Hyclone) or 10% newborn calf serum (BCS; Gibco), and 1% antibiotics (Hyclone) at 37°C in 5% CO2.
Preparation of Dendropanax morbifera leaf extracts
Dendropanax morbifera leaf (DL) extracts were prepared by adding leaves to 600-L of water or 20% or 50% ethanol (DLE20, DLE50) and incubating at 80°C for 3 h. Subsequently, solvents were filtered and concentrated to 60 L before freeze drying; dried extracts were then mixed and dissolved in water for 10 min. Solutions were then centrifuged at 17,000 g for 10 min, and supernatants were collected for experiments.
Preparations of dexamethasone, isobutyl methylxanthine, and insulin
Dexamethasone (D4902; Sigma-Aldrich), isobutyl methylxanthine (IBMX, I7018; Sigma-Aldrich), and insulin (I9278, Sigma-Aldrich) solutions were filtered using 0.2-μm syringe filters, and dexamethasone and IBMX were diluted to 1 mM and 0.5 M, respectively.
Differentiation of 3T3-L1 cells
Two days after reaching confluence (day 0), cells were cultured in DMEM differentiation medium (DM) containing the following: (BCS; Gibco), 0.5-mM IBMX, 1 μM dexamethasone (Dex; Sigma), and 167-nM insulin (Sigma). After differentiation, the medium was replaced with DMEM post-DM containing 167-nM insulin for another 2 days. Thereafter, cells were cultured in normal DMEM, and the medium was changed every 2 days. 3T3-L1 preadipocytes were treated with the indicated concentrations of extracts during days 2–7.
Cell viability assays
Cell viability was determined using WST-1 (EZ-CYTOX, Daeillab) assays according to the manufacturer's instructions. Briefly, cells were plated in 96-well plates at a density of 1–3 × 103 cells/well and were incubated at 37°C in a CO2 incubator. Cells were incubated with various concentrations of D. morbiferus water (DLW), DLE20, or DLE50 extracts for 24, 48, or 72 h, and 10-μL of WST-1 reagent was then added to each well and incubated for 90 min at 37°C in a CO2 incubator. Absorption was then measured at 450 nm (reference wavelength 600–650 nm) using a microplate reader (Model 550; Bio-Rad Laboratories, Hercules, CA, USA). Data are presented as means ± standard deviations (SDs) of three independent experiments.
Oil red O staining
On day 9 of differentiation, mature 3T3-L1 adipocytes were washed three times with PBS and were then fixed with 4% (v/v) formaldehyde at room temperature for at least 30 min. Fixed adipocytes were then washed three times using tap water, and mature adipocytes were stained with 3-mg/mL Oil red O (ORO) solution in isopropanol at room temperature for 15 min. Subsequently, ORO-stained 3T3-L1 cells were washed three times with tap water and dried, and stained lipid droplets were then dissolved in 300 μL of DMSO and were transferred to 96-well microplates. The absorbance of dissolved ORO was determined at 510 nm using a microplate reader.
AdipoRed assays of intracellular triglycerides
Glycerol release was measured using a commercially available Adipolysis Assay Kit according to the manufacturer's instructions. Briefly, differentiated adipocytes in 96-well plates were stimulated with S1P or a positive control solution containing isoproterenol for 24 h. After stimulation, cell culture supernatants were collected from each well and were stored at −20°C until use. Subsequently, 100-μL aliquots of free glycerol assay reagent were incubated with 25-μL supernatant samples for 15 min at room temperature, and absorbance was measured at 540 nm.
2-NBDG assays; glucose uptake measurements
At day 9 of differentiation, adipocytes were incubated for 24 h with the respective test solutions. In these experiments, ethanol was used as a negative control and 10-μM rosiglitazone was used as a positive control. On the following day, cells were rinsed with 1 × PBS and were incubated for 60 min at 37°C in the dark in DMEM containing 80-μM fluorescent glucose. As a second positive control, cells were treated with 100-nM insulin during incubation with 2-NBDG, and insulin-stimulated glucose uptake was determined. The reaction of 2-NBDG uptake was terminated by washing cells with precooled 1 × PBS, and remaining fluorescence in the cells was measured using a fluorescence microplate reader at an excitation wavelength of 485 nm and an emission wavelength of 535 nm. Fluorescence in the absence of 2-NBDG was subtracted from all values.
RNA extraction and real-time quantitative PCR analyses
Total RNA was isolated using RNA EasySpin kits according to the manufacturer's instructions (Intron Biotechnology, Seongnam, Korea). Briefly, total RNA from each sample was reverse transcribed using random primers and StrataScript™ reverse transcriptase kits (Agilent technology, Santa Clara, CA, USA) according to the protocols suggested by the manufacturer. All samples were normalized to mouse actin and were expressed as fold induction, and all reactions were performed in triplicate. Relative expression levels were calculated with SD using the comparative quantification method with the following primers: 5′-CTGCGAGCACGAGACGTCTA-3′ and 5′-GCCAGGAACTCGTCGTTGAA-3′, CEBPα; 5′-GTTTCGGGAGTTGATGCAATC-3′ and 5′-AACAACCCCGCAGGAACAT-3′, CEBPβ; 5′-TGACTTGGCCATATTTATAGCTGTCA-3′ and 5′-CGATGGGCTTCACGTTCAG-3′, PPARγ; 5′-CCAGAGGGTGAGCCTGACAA-3′ and 5′-AGCCTCTGCAATTTCCAGATCT-3′, SREBP1; 5′-TCCCCAAGTTCTTCACGTTCA-3′ and 5′-CAGGCTCCAAGTGGCGATAA-3′, AMPK; 5′-AACCCCTGGCAGGAAAGG-3′ and 5′-TGAACGCTGAGCGATACACAT-3′, adiponectin; 5′-GACCCTGACTCCAAGTTATTCGA-3′ and 5′-CGTCAAGCGGGAGACAGACT-3′, FAS; 5′-GTGACTGGTGGGAGGAATAC-3′ and 5′-GAGCATCTCCATGGCGTAG-3′, CPT1; 5′-CTATGAGCTGCCTGACCGTC-3′ and 5′-AGTTTCATGGATGCCACAGG-3′, mACTIN.
Western blot analysis
Before Western blotting analysis, 3T3-L1 cells were washed with cold PBS and were harvested by scraping. Cell extracts were then prepared in lysis buffer containing 50-mM Tris-HCl (pH 7.5), 150-mM NaCl, 1% NP-40, 10-mM NaF, 10-mM sodium pyrophosphate, and protease inhibitors and were incubated for 30 min on ice. Lysates were then centrifuged at 20,000 g for 10 min at 4°C, and total proteins were separated using 8% and 12% sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) and were transferred to nitrocellulose membranes. Membranes were then blocked by incubating for 2 h in 5% (w/w) nonfat Difco™ skim milk (BD Biosciences, Franklin Lakes, NJ, USA) in 1 × PBST. Blocked membranes were then incubated with primary antibodies for 2 h or overnight at 4°C. We used the following antibodies: anti-fatty acid synthase (FAS; Cell Signaling Technology, #4233), anti-peroxisome proliferator-activated receptor gamma (PPARγ; Cell signaling Technology, #2443), anti-CCAAT-enhancer-binding proteins alpha (C/EBPα; Cell Signaling Technology, #8178), anti-sterol regulatory element-binding protein 1 (SREBP1; Santa Cruz Biotechnology, sc-365513), and anti-β-actin (Sigma Aldrich, A5441). After washing three times with 1 × PBST, membranes were incubated with secondary horseradish peroxidase-conjugated antibodies for 1 h. Protein signals were then visualized.
Statistical analyses
Statistical analyses were performed using SPSS (SPSS, Inc., Chicago, IL, USA). Differences between treatment groups were identified using ANOVA and were considered significant when P < .05.
Results
Cytotoxicity of DL extracts in 293T, HepG2, and 3T3-L1 cells
To evaluate cytotoxicity of DL extracts, 293T cells were treated with DLW, DLE20, and DLE50 extracts at 25, 50, and 100 μg/mL for 24 to 48 h. In these experiments, cell viability was unchanged in the presence of DL extracts (DLW, DLE20, DLE50) but was greater in the presence of DLW than in the presence of DLE20 and DLE50 (Fig. 1A). In subsequent experiments, cells were treated with DLW extracts only at 9 concentrations between 5 and 500 μg/mL, and the effects of extracts on viability of 293T and HepG2 cells were evaluated after 24 or 48 h by using WST-1 assays (Fig. 1B). We also confirmed the cytotoxicity of DLW extracts at 100–1000 μg/mL in HepG2 cells after 24–48-h treatment (data not shown). Time dependence of DLW effects on adipogenesis was also assessed in terms of cell viability in 3T3-L1 preadipocytes, and these experiments showed no significant changes between time points. Taken together, the present data show that DLW extracts are not toxic in preadipocytes or adipocytes (Fig. 1C.).

Effects of DL treatments on cell viability. DL extracts are not toxic in the various cells and concentrations.
DLW inhibits adipogenesis in 3T3-L1 adipocytes
To assess antiobesity and cholesterol-lowering effects of DLW extracts, we performed experiments in 3T3-L1 cells after differentiation with methyl isobutyl xanthine (MDI), dexamethasone, and insulin for 9 days. 14,15 Antiadipogenic effects of DLW in 3T3-L1 cells were determined on day 7 by incubating preadipocytes in six-well plates with MDI media with or without DLW during the adipocyte differentiation phase. After differentiation over 7 days in the presence of various concentrations of DLW in adipogenic medium, lipid accumulation was significantly reduced (Fig. 2A). Adipocytes had increased in size and spread, with greater numbers of lipid droplets than undifferentiated control cells. However, treatment with DLW extracts led to decreased cell sizes and numbers of lipid droplets. 16 –20 The differentiation of preadipocytes into adipocytes is associated with increased numbers of ORO-stained cells due to lipid accumulation. 21,22 Accordingly, microscope observations revealed gradual reductions in numbers of lipid droplets with increasing concentrations of DLW. In addition, cell densities were increased under all treatment conditions, indicating inhibitory effects of DLW extracts on adipogenesis in 3T3-L1 cells (Fig. 2B).

DLW inhibits 3T3-L1 adipogenesis. Lipid accumulation decreases are dependent on DLW concentration.
DLW reduces triglyceride accumulation and glucose uptake in adipocytes
To confirm that DLW inhibits triglyceride (TG) accumulation, we measured TG levels directly in 3T3-L1 cells after differentiation for 2 days. 23 Under these conditions, DLW treatments inhibited TG accumulation during the differentiation of 3T3-L1 preadipocytes (Fig. 3A). Analysis of accumulated TGs on day 7 revealed significantly lower lipid contents in cells that were treated with 300-μg/mL DLW. Moreover, the inhibitory effects of DLW on TG accumulation during adipogenesis were dose dependent, with a 37.7% decrease after 7 days of treatment with 300-μg/mL DLW, and did not affect cell viability in the WST-1 assays (Fig. 1C).
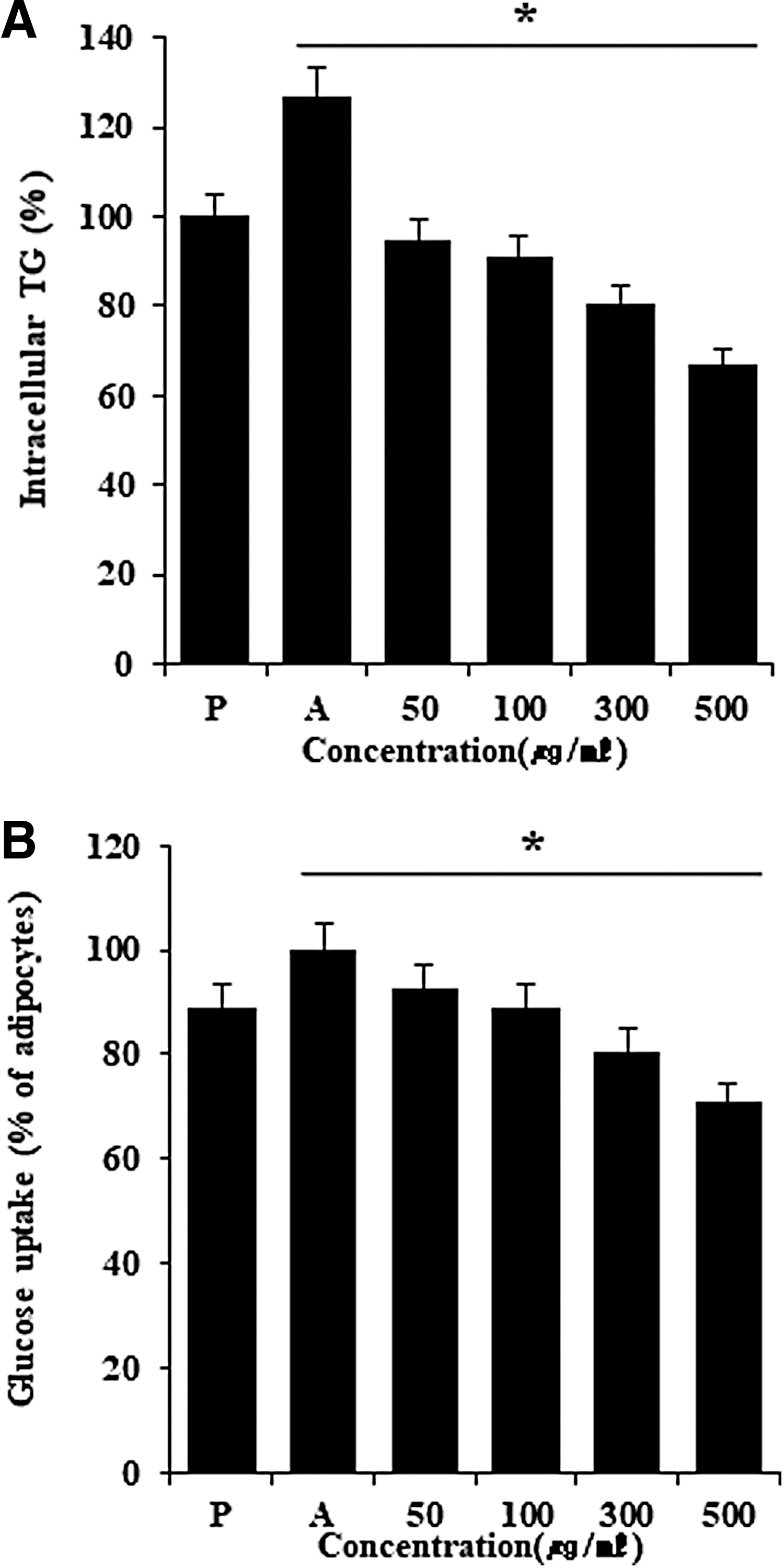
Inhibitory effect of DLW on intracellular triglycerides and glucose uptake in 3T3-L1 cells. DLW inhibits intracellular triglycerides and glucose uptake in adipocyte-differentiated 3T3-L1 cells.
To study the effects of DLW on lipolysis, differentiated adipocytes were incubated with various concentrations of DLW for 24 h and glycerol levels were determined in culture medium. DLW affected glucose uptake and inhibited lipid accumulation during adipogenic differentiation, but did not affect lipolysis in differentiated adipocytes (Fig. 3B). 24,25 Moreover, the effects of DLW extracts were apparent at concentrations above 300 μg/mL, and clear antiadipogenic effects, with restoration of preadipocyte morphology and lower lipid content than in control adipocytes, were observed at 500 μg/mL. These results indicate that DLW effectively blocks adipocyte differentiation in 3T3-L1 preadipocytes. In addition, maximal inhibitory effects of DLW were observed at 500 μg/mL (Fig. 3A, B). 26
DLW suppresses protein and mRNA expression of adipogenesis-related genes in 3T3-L1 cells
To confirm the inhibitory effects of DLW on adipocyte differentiation, protein and mRNA expression levels of the differentiation markers, peroxisome proliferator activated receptor gamma (PPARγ), 27 –29 CCAAT/enhancer binding protein alpha (C/EBPα), 29,30 and SREBP1, 31,32 were determined. These experiments showed greater decreases in protein expression following DLW treatment than in differentiated control adipocytes. Protein expression levels of FAS, PPARγ, SREBP1, and C/EBPα were also reduced (Fig. 4A), and when 3T3-L1 preadipocytes were differentiated in MDI, mRNA levels increased correspondingly with differentiation compared with the control. Under these conditions, DLW treatments led to significant dose-dependent reductions in C/EBPα, C/EBPβ, and PPARγ mRNA expression levels, and in PPARγ protein expression (Fig. 4A, B). Because DLW strongly induces apoptosis in the presence of ceramide, we determined 3T3-L1 cell numbers during differentiation in the presence of 500-μg/mL DLW. In a previous study, 24-h MDI induction led to mitosis and doubling of cell numbers. In agreement, cell numbers were doubled following 24-h MDI treatment in the presence or absence of DLW extract in our study. These results demonstrated that DLW has antiadipogenic activity and acts by downregulating transcription factors of adipocyte differentiation.
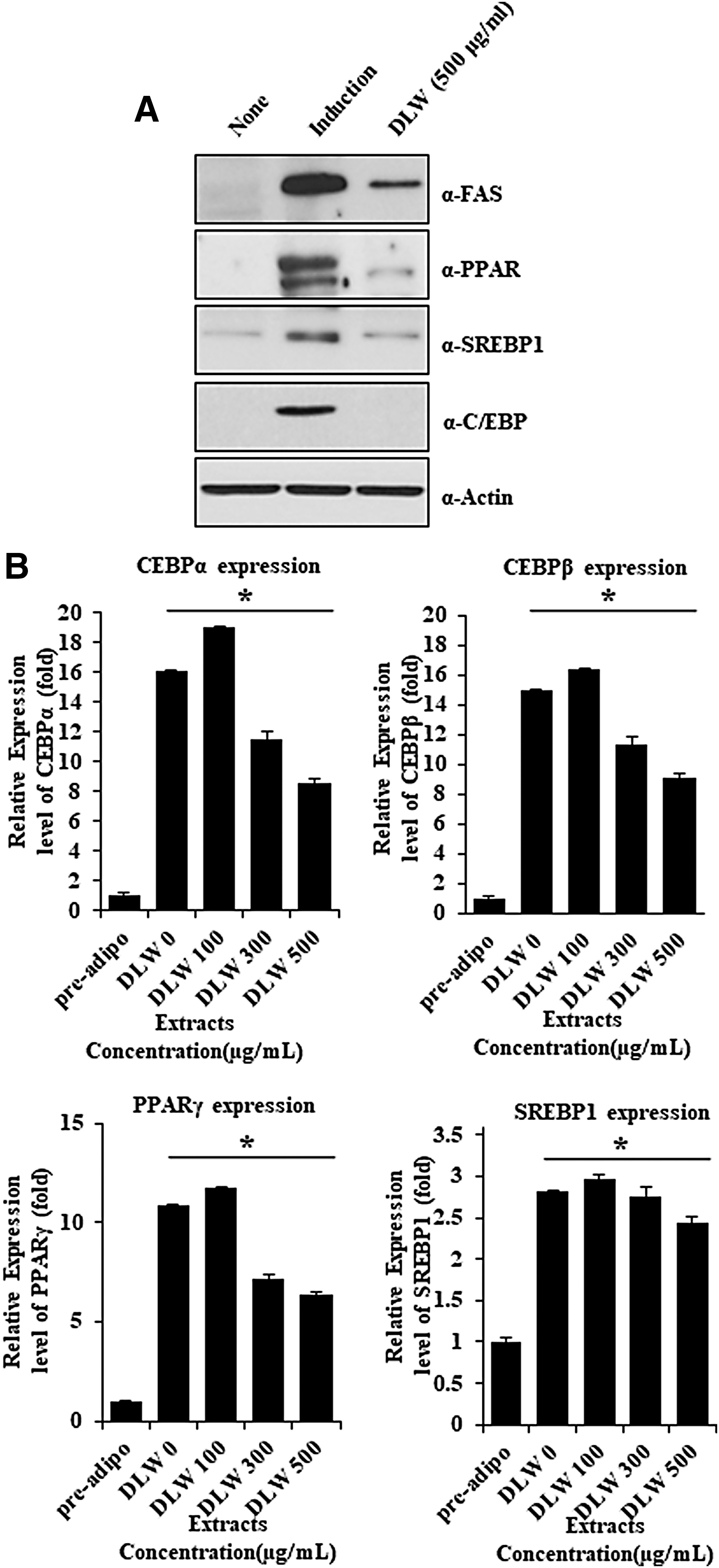
Effects of DLW on protein and mRNA expression levels of adipogenesis-related genes in 3T3-L1 cells. DLW inhibits the protein and RNA level of adipogenesis-related genes.
Discussion
Adipocytes are derived from mesenchymal stem cells that can differentiate into myoblasts, chondroblasts, osteoblasts, or adipocytes. Adipocytes are essential components of fat tissues and play important roles in TG storage and endocrine signaling. Adipogenesis from preadipocytes is closely associated with the development and stimulation of adipocytes according to systemic regulation of nutrients, hormones, and cytokines. Hence, adipocyte differentiation involves an elaborate network of transcription factors that regulate the expression of numerous mature adipocyte genes. 32 –38 Among transcription factors that promote preadipocyte differentiation and influence adipogenesis, PPARγ is considered the master regulator and other adipogenic transcription factors include the CCAAT/enhancer binding proteins C/EBPα, C/EBPβ, and C/EBPγ. These transcription factors are necessary for the expression of adipocyte-specific genes such as adiponectin, and PPARγ and C/EBPα in particular, and are regulated by the mitogen-activated protein kinase pathway during adipogenesis. 39
In the present experiments, as confirmed by microscopic observations, DLW treatments suppressed sizes and numbers of lipid droplets. The treatment also inhibited intracellular TG levels and glucose uptake in 3T3-L1 cells (Fig. 3). Hence, we investigated cytotoxic, antiobesity, and cholesterol-lowering effects of DLW in 3T3-L1 cells (Fig. 1C). Herein, we show unprecedented data demonstrating that D. morbiferus is nontoxic after 24–48-h treatments at 25–100 μg/mL in 293T cells (Fig. 1A) and report increased cell viability at 48 h. Similarly, in further 24–72-h experiments with DLW concentrations of 5–500 μg/mL, no cytotoxicity was observed in 293T, HepG2, and 3T3-L1 cell lines (Fig. 1B, C).
Following induction of 3T3-L1 differentiation, increases in fat mass are accomplished by increases in adipocyte numbers and sizes. Previous cellular and molecular studies of obesity show that changes in the numbers of adipocytes (hyperplasia) and adipocyte sizes (hypertrophy) reflect dietary factors. The present data show significant decreases in lipid accumulation, adipocyte sizes, numbers of lipid droplets, intracellular TG levels, and glucose uptake following treatment with DLW. In addition, the adipogenic transcriptional factors C/EBPs, PPARγ, and SREBP-1c and the lipogenic enzymes FAS and acetyl-CoA-carboxylase, considered key regulators of adipogenesis, were suppressed by treatments with DLW.
Taken together, the experiments shown in Figures 2, 3, and 4 demonstrate that DLW reduces lipogenesis and lipid accumulation in adipocytes, and these changes were reflected in intracellular TGs and glucose uptake rates. These data also suggest that DLW suppress lipogenesis by decreasing the numbers and sizes of adipocytes and by downregulating adipogenesis-related genes, warranting consideration of DLW as a natural source of agents that ameliorate obesity.
In conclusions, we demonstrated that DLW inhibits adipogenesis and lipogenesis in 3T3-L1 cells and suggest that DLW extracts contain natural antiadipose agents. The suppression of adipogenesis-related genes leading to reduced TG levels and glucose uptake rates further confirmed this conclusion. 40 –43 Hence, DLW extracts may provide a natural therapy for obesity and hypercholesterolemia.
Footnotes
Acknowledgments
This work was supported by the “Food Functionality Evaluation program” under the Ministry of Agriculture, Food and Rural Affairs, and partly by the Korea Food Research Institute.
Author Disclosure Statement
No competing financial interests exist.