
Abstract
Some lactic acid bacteria (LAB) and their cellular components have antiobesity effects. In this study, we evaluated the antiadipogenic effects of a mixture of two LAB—Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032—using 3T3-L1 preadipocytes and HepG2 hepatocarcinoma cells. 3T3-L1 cells treated with a 1:1 ratio of HY7601 and KY1032 during differentiation showed reduced lipid accumulation by Oil Red O staining, as well as decreased leptin secretion and mRNA expression of peroxisome proliferator-activated receptor-γ and CCAAT/enhancer binding protein-α. HY7601 and KY1032 treatment also suppressed mitochondrial biogenesis and inhibited the expression of genes encoding mitochondrial transcription factors, as well as those related to fatty acid synthesis in HepG2 cells. The antiadipogenic effects of LAB were associated with the cell membrane fraction. These results demonstrate that a mixture of two LAB (HY7601 and KY1032) inhibits adipogenesis in preadipocytes and liver cells and is a potential therapeutic strategy for the treatment of obesity.
Introduction
O
Intestinal microbiota play a critical role in regulating host metabolic functions. 5 Metagenomic studies have identified gut microbiota as an environmental factor that can influence whole-body metabolism by modulating not only energy balance but also immune and gut barrier functions, 6,7 thereby altering nutrient absorption; moreover, the intestinal microbiome composition has been shown to differ between obese and nonobese individuals, 8 suggesting that it contributes to the development of obesity. Many probiotic strains have been shown to reduce obesity in experimental models. 9 Members of the Lactobacillus and Bifidobacterium genera have shown beneficial effects, including weight loss, a decrease in visceral fat, and alleviation of glucose intolerance in many animal and some human studies of metabolic syndromes. 10,11
We recently reported that oral intake of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 (hereafter referred to as HY7601 and KY1032, respectively) isolated from kimchi, a Korean traditional fermented cabbage dish, reduced body fat in mice and humans. 12 –15 Health benefits such as antioxidative, lipid-lowering, and antiatherogenic effects of kimchi are well established. 16 Clinical studies prove that consumption of kimchi reduces bodyweight in obese patients, but the main mechanism is yet to be revealed. 17,18 Kimchi contains numerous types of microorganisms, as novel strains are still being discovered, 19,20 and the composition of microflora in kimchi shows diversity depending on factors, including fermentation stage, temperature, and major ingredients. 21
For example, Leuconostoc mesenteroides are reported to predominate in the early stages of fermentation, whereas Lactobacillus plantarum becomes predominant as the pH gradually falls during fermentation. 22,23 In addition, in another study, Lactobacillus curvatus, Lactobacillus sakei, and Lactobacillus mesenteroides were reported to be the most dominant fermenting bacteria in kimchi. 24 In the present study, we selected HY7601 and KY1032, isolated from various types of kimchi, by measuring the lipid contents in preadipocytes treated with lactic acid bacteria (LAB). Fat accumulation in adipose tissue and liver was reduced in mice fed with high-fat diets (HFD) containing HY7601 and KY1032. 15 When mice were fed a HFD to induce obesity and then given a HFD supplemented with probiotics for an additional 10 weeks, body weight gain and fat accumulation along with the levels of obesity-related factors such as plasma insulin, leptin, total cholesterol, and liver toxicity biomarkers were reduced. 14 These results were corroborated in a human trial in which consumption of HY7601 and KY1032 resulted in weight loss and reduced body fat mass in overweight subjects. 13 Based on these findings, the present study investigated the mechanistic basis for the antiadipogenic effects of HY7601 and KY1032 using 3T3-L1 preadipocytes and HepG2 hepatocarcinoma cells.
Materials and Methods
Screening and isolation of LAB strains and preparation of the cell extract
LAB of 25 strains were isolated from various types of kimchi, and screening was conducted by measuring lipid contents in LAB-treated 3T3-L1 mouse preadipocytes using AdipoRed assay reagent (Lonza, Switzerland). HY7601 and KY1032 were selected, and cell extracts were prepared as previously described. 25 Briefly, HY7601 and KY1032 were grown anaerobically in deMan–Rogosa–Sharpe medium at 37°C for 18 h; the cells were collected by centrifugation and washed with phosphate-buffered saline (PBS) and were counted and resuspended in PBS at 10 10 colony-forming U/mL followed by sonication. The cells were centrifuged at 1500 × g for 10 min, yielding a supernatant containing whole cell extract. To separate the cell cytosol and membrane, the whole cell extract was centrifuged at 6500 × g for 30 min, yielding the supernatant containing cell cytosol and pellet containing the cell membrane. 26 Samples were stored at −20°C until use.
Cell culture
Cell lines were purchased from the American Type Culture Collection (Manassas, VA, USA), and cell viability was evaluated using a Cell proliferation Kit I (Roche, Mannheim, Germany). 3T3-L1 mouse preadipocytes were cultured as described elsewhere. 27 Briefly, the cells were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) in an atmosphere of 5% CO2 at 37°C until they reached confluence (day 0); 2 days later (day 2), cells were stimulated with 3-isobutyl-1-methylxanthine, dexamethasone, and insulin (MDI) induction medium composed of DMEM, 10% FBS, 1 μg/mL insulin, 0.5 mM 3-isobutyl-1-methylxanthine, and 1 μM dexamethasone. After 2 days (day 4), the medium was replaced with DMEM containing 10% FBS and 1 μg/mL insulin followed by incubation for 2 days (until day 6) in maintenance medium containing 10% FBS. After 4 days (day 10), over 90% of cells were mature adipocytes containing fat droplets. To evaluate the effects of LAB, HY7601, KY1032, or the two combined at different ratios were added to the medium at total concentration of 0.1%, starting from day 0.
HepG2 cells were cultured as described elsewhere. 28 Briefly, cells were cultured in DMEM containing 10% FBS in an atmosphere of 5% CO2 at 37°C until they reached 70% confluence, then starved overnight in FBS-free medium before treatment with free fatty acid (FFA) containing bovine serum albumin (BSA). The FFA/BSA solution was prepared by adding 100 mM palmitate to 5% BSA solution at a final concentration of 5 mM. 29 Palmitate and BSA were dissolved in 0.1 M NaOH and distilled water, respectively. The FFA solution was diluted 1:5 in serum-free DMEM at final concentrations of 1 mM FFA/1% BSA for experiments. Cells were treated with 0.1% HY7601, KY1032, or both in FFA/BSA solution for 24 h.
Oil Red O staining
Neutral triglycerides and lipids were stained with Oil Red O as previously described, 30 with slight modification. Cells were washed with PBS and fixed with 10% formaldehyde for 10 min; they were then washed and fixed with 10% formaldehyde for 30 min before staining with 60% Oil Red O diluted in distilled water. After 30 min, cells were washed twice and photographed under a microscope. The dye retained in cells was eluted by adding 100% isopropanol and quantified with a microplate reader at 500 nm.
Leptin expression
3T3-L1 and HepG2 cells were collected on day 9 and after 24 h of treatment, respectively. Leptin expression was measured using a Mouse Leptin Immunoassay Kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions.
RNA extraction
RNA was extracted from 3T3-L1 cells and HepG2 cells on day 9 and after 24 h of treatment, respectively. The RNA was collected using an Easy-spin Total RNA Extraction Kit (Intron Biotechnology, Seoul, Korea) according to the manufacturer's instructions.
Reverse transcription and real-time polymerase chain reaction
Total RNA (2 μg) was reverse transcribed to cDNA using the Omniscript RT Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. The cDNA served as a template for polymerase chain reaction (PCR) amplification using the TaqMan Gene Expression Master Mix (Applied Biosystems, Foster City, CA, USA) and specific TaqMan primers (Applied Biosystems; Table 1) under the following conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Data are representative of three independent experiments.
ACC, acetyl-CoA carboxylase; C/EBPα, CCAAT/enhancer-binding protein alpha; FAS, fatty acid synthase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; LPL, lipoprotein lipase; mtTFA, mitochondrial transcription factor A; Nrf1, nuclear respiratory factor 1; PPAR-γ, peroxisome proliferator-activated receptor gamma; PGC1α, peroxisome proliferator-activated receptor gamma coactivator 1-alpha; PPAR-α, peroxisome proliferator-activated receptor alpha; SREBP-1c, sterol regulatory element-binding transcription factor 1.
Detection of mitochondria biogenesis
On day 9, 3T3-L1 cells were stained with 20 nM MitoTracker Green FM (Molecular Probes, Eugene, OR, USA) for 30 min at 37°C according to the manufacturer's instructions. After 30 min, the medium was refreshed, and fluorescence was measured at excitation/emission wavelengths of 485/516 nm.
Statistical analysis
Experiments were performed at least thrice, and data are presented as the mean ± SD of triplicate preparations. Differences between groups were evaluated with the unpaired Student's t-test and were deemed statistically significant at P < .05.
Results
LAB have antiadipogenic effects in preadipocytes
3T3-L1 cells were treated with various LAB during adipocyte differentiation, and results showed that cells treated with HY7601 and KY1032 had the lowest lipid contents (Supplementary Fig. S1; Supplementary Data are available online at
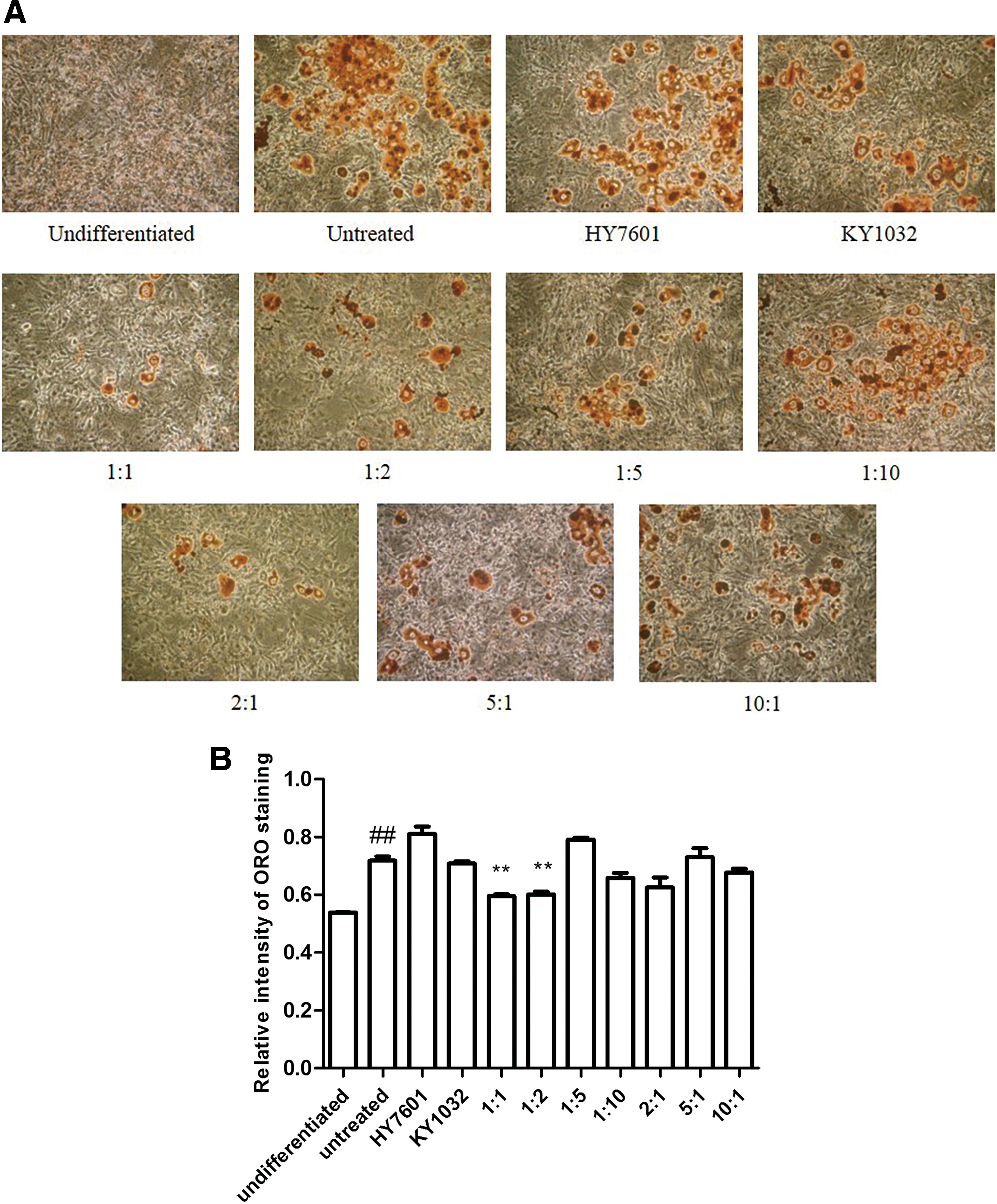
Effect of LAB on preadipocytes during differentiation. 3T3-L1 cells were treated with 0.1% HY7601 and KY1032 at various ratios during differentiation.
LAB suppress leptin production and transcription of adipogenic genes in preadipocytes
Leptin is released by adipocytes, and the expression and secretion of leptin are correlated with adipose cell mass and size. 31,32 To measure leptin secretion in 3T3-L1 cells, the supernatant of 3T3-L1 cells was collected on day 9. Treatment with equivalent amounts of HY7601 and KY1032 decreased leptin secretion, indicating that preadipocyte differentiation was inhibited (Fig. 2A).
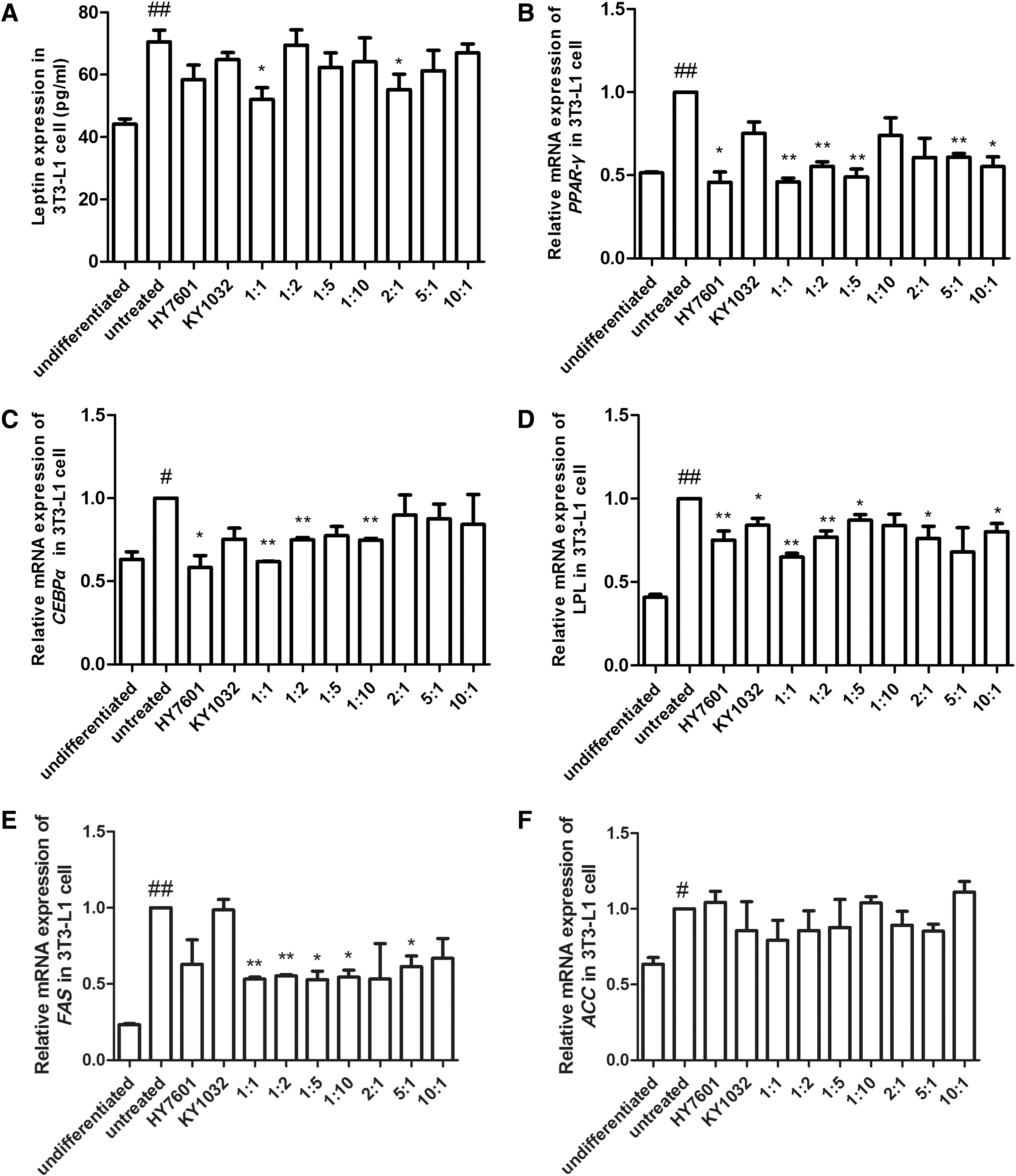
Leptin and adipogenic transcription factor expressions in preadipocytes treated with LAB during differentiation. 3T3-L1 cells were treated with 0.1% HY7601 and KY1032 at various ratios during differentiation. The levels of
To examine the molecular changes associated with HY7601 and KY1032 treatment, expression levels of the adipogenic genes, peroxisome proliferator-activated receptor (PPAR)-γ, CCAAT/enhancer-binding protein (C/EBP)α, lipoprotein lipase (LPL), fatty acid synthase (FAS), and acetyl-CoA carboxylase (ACC), were examined. Treatment with HY7601 alone and with various ratios of HY7601 and KY1032 reduced PPAR-γ expression, with a 1:1 ratio of HY7601 and KY1032 having the greatest effect (53% decrease relative to untreated differentiated cells) (Fig. 2B). C/EBPα, LPL, and FAS levels were also reduced (by 38%, 35%, and 47%, respectively) upon treatment with the same ratio of HY7601 and KY1032 (Fig. 2C–E). The level of ACC was also reduced by 21%, but with no significance (Fig. 2F).
Antiadipogenic effects of LAB are associated with the membrane fraction
To identify the origin of the antiadipogenic effects of LAB, whole cell extracts of HY7601 and KY1032 were lysed, and the cytosolic and membrane fractions were separated by centrifugation as the supernatant and pellet, respectively; these were used to treat 3T3-L1 cells with a 1:1 ratio during differentiation. Oil Red O staining revealed that lipid accumulation was reduced by treatment with whole cell extracts or pellet (membrane fraction), which was confirmed by the quantitative analysis (Fig. 3).
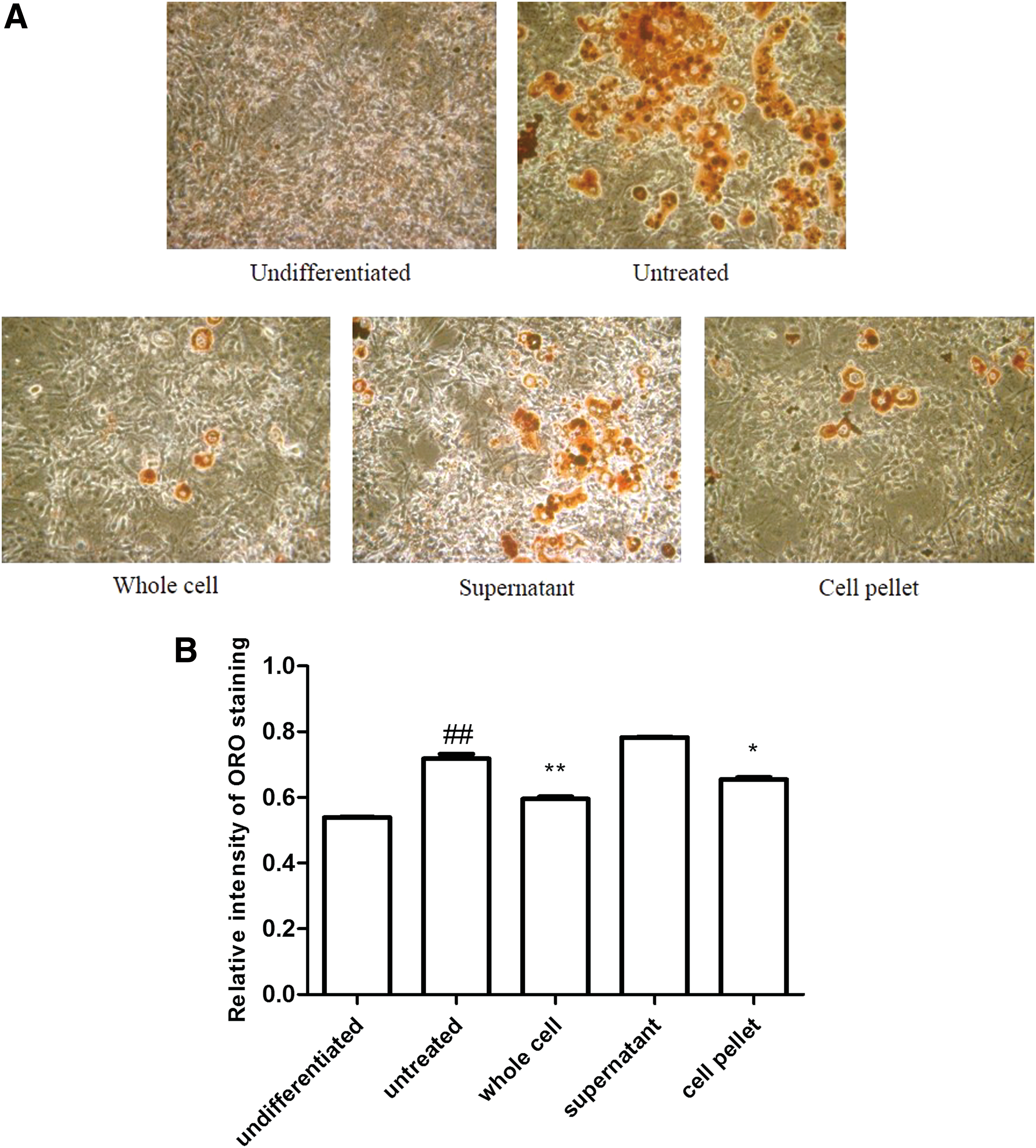
Effect of LAB whole cell extracts, supernatants, and pellets on preadipocytes during differentiation. 3T3-L1 cells were treated with 0.1% whole cell extracts, supernatants, or cell pellets of HY7601 and KY1032 during differentiation.
We also measured the expression levels of the adipogenic genes PPAR-γ and C/EBPα by reverse transcription PCR. Treatment of 3T3-L1 cells with the whole cell extract and cell pellet reduced PPAR-γ level by 40% and 37%, respectively, compared to untreated 3T3-L1 cells, whereas the supernatant reduced the level by 18%, which was a nonsignificant difference. In contrast, C/EBPα expression was reduced by 52%, 36%, and 52% in the presence of whole cell extract, supernatant, and cell pellet, respectively (Fig. 4).
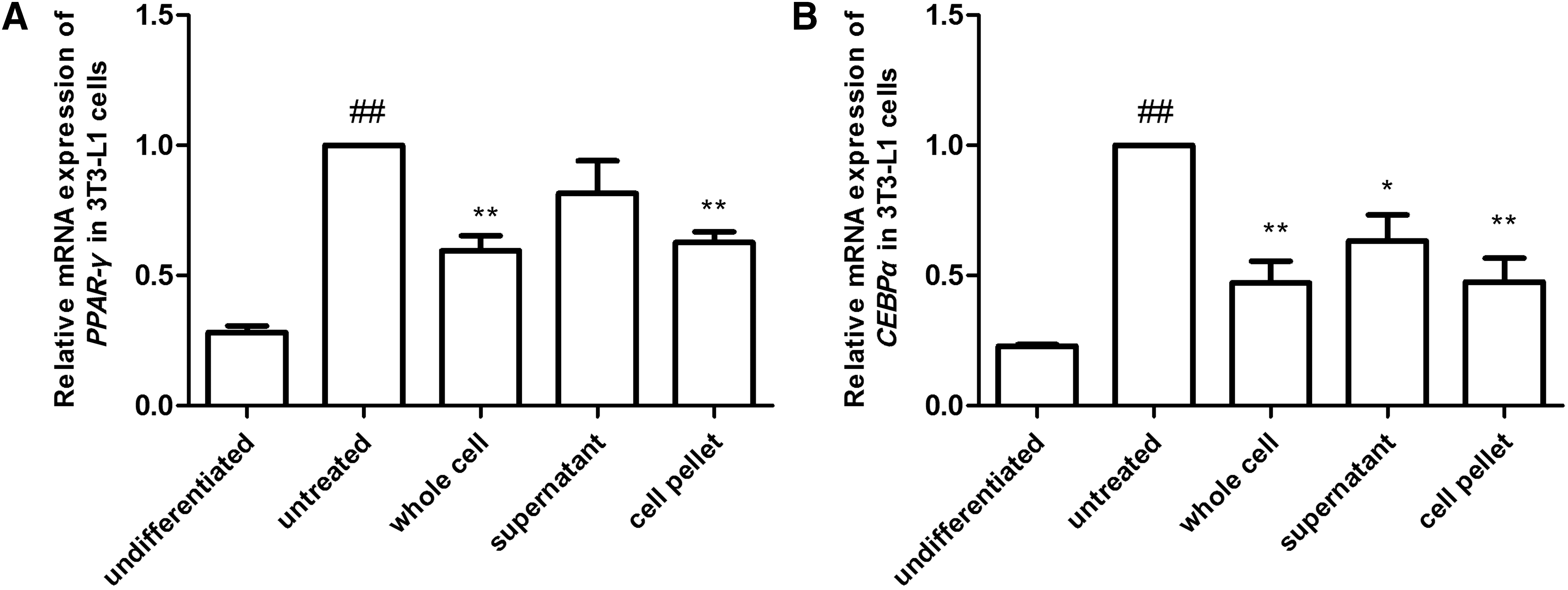
mRNA expressions of adipogenic genes in preadipocytes treated with LAB whole cell extracts, supernatants, or cell pellets during differentiation. 3T3-L1 cells were treated with 0.1% HY7601 and KY1032 whole cell extracts, supernatants, or cell pellets during differentiation. The expression levels of
LAB inhibit mitochondrial biogenesis during preadipocyte differentiation
Mitochondrial biogenesis and activity increase dramatically during adipocyte differentiation. 33 To investigate the effect of LAB on mitochondrial biogenesis, 3T3-L1 cells were treated with HY7601 and KY1032 during differentiation, and mitochondrial biogenesis was measured by staining 3T3-L1 cells with MitoTracker dye after full differentiation. HY7601 and KY1032 suppressed mitochondrial biogenesis relative to untreated 3T3-L1 cells (Fig. 5A). Furthermore, there was no change in nuclear respiratory factor 1 expression; however, the levels of other mitochondrial transcription factors (mtTF) such as PPAR-γ coactivator (PGC)-1α and mtTFA were decreased by 32% and 35%, respectively, relative to untreated 3T3-L1 cells, although only the latter was statistically significant (Fig. 5B–D).
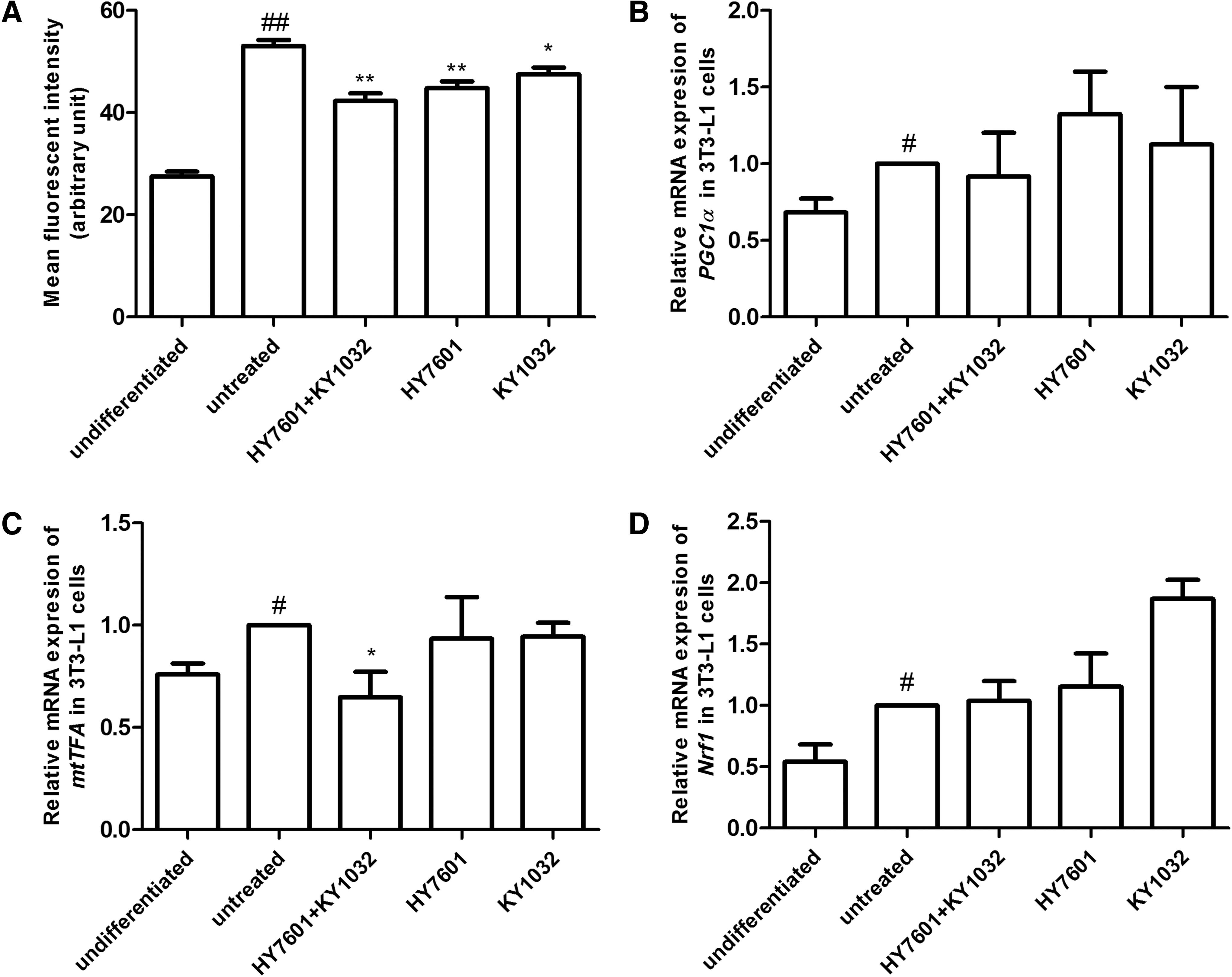
Effects of LAB on mitochondrial biogenesis during preadipocyte differentiation. 3T3-L1 cells were treated with 0.1% HY7601, KY1032, or both during differentiation. Mitochondrial biogenesis was evaluated by
LAB prevent lipid accumulation in hepatocytes
To assess lipid accumulation in HepG2 hepatocytes, cells were treated with HY7601, KY1032, or both in FFA/BSA solution for 24 h. Oil Red O staining revealed that lipid accumulation was reduced by HY7601 or KY1032 single treatment; however, the combination of both LAB reduced the number of lipid droplets in HepG2 cells (Fig. 6A). In addition, HY7601 and KY1032 exhibited no cytotoxity in HepG2 cells, as cell viability showed 1.31%, 5.25%, and 9.48% increases in cells treated with HY7601+KY1032, HY7601, and KY1032, respectively, compared to untreated cells (Supplementary Fig. S3).
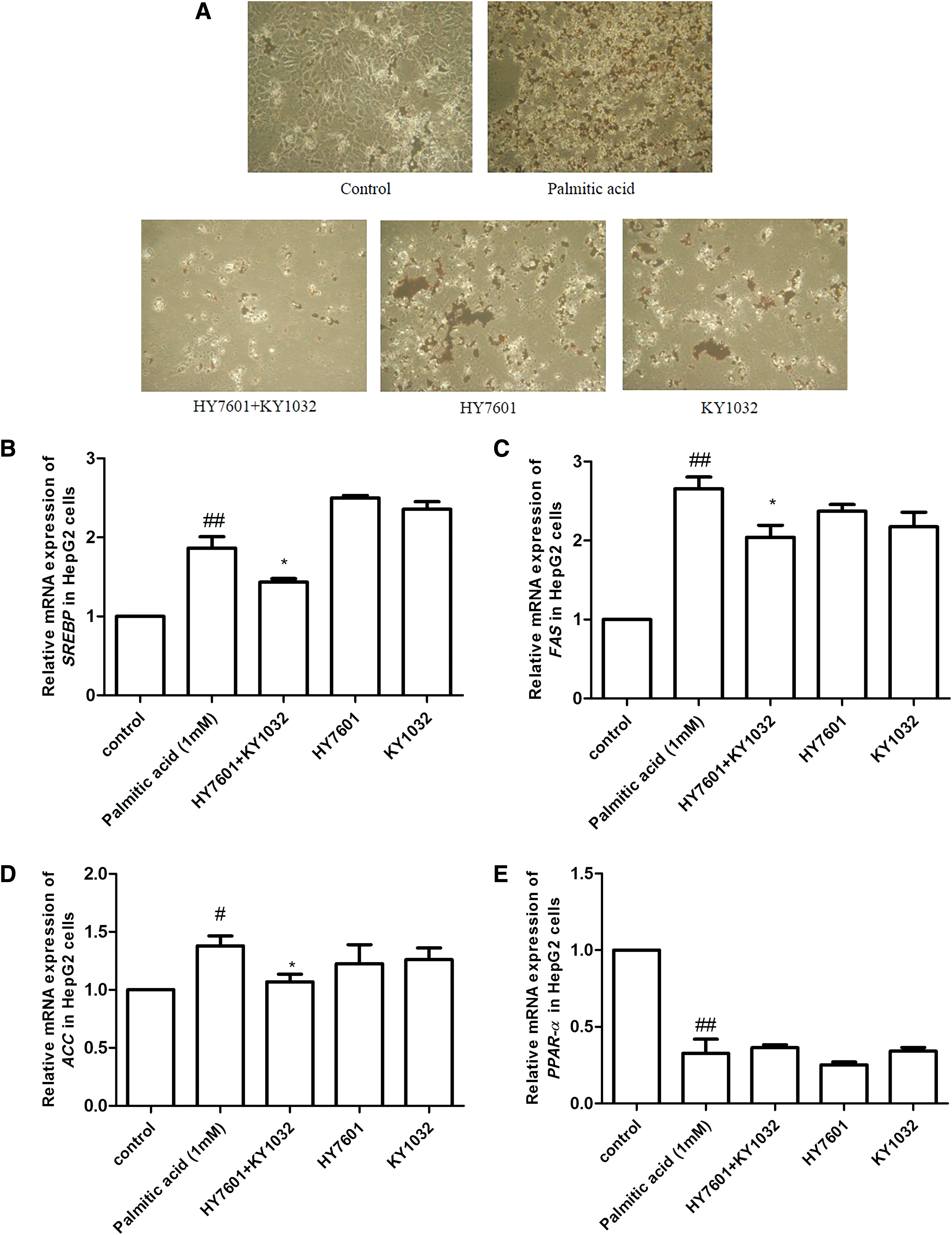
Effects of LAB on liver cells. HepG2 cells were treated overnight with 0.1% HY7601, KY1032, or both in the presence of 1% FFA. Lipid accumulation was evaluated by
We also examined the expression of several genes that are upregulated in fatty liver, including sterol regulatory element-binding transcription factor (SREBP)-1c and FAS. 34 SREBP-1c and FAS levels were both reduced by 23% in cells treated with HY7601 and KY1032 compared to those treated with FFA/BSA only. Expression of ACC, another transcription factor related to fatty acid synthesis, was also decreased by 22%. However, there was no change in the level of PPAR-α, a transcription factor related to fatty acid catabolism (Fig. 6B–E). Thus, LAB prevent fatty liver by suppressing FAS in liver cells.
Discussion
Obesity is a chronic metabolic disease that has become a global healthcare burden. It is also related to the development of various metabolic diseases, including type 2 diabetes, hypertension, coronary heart disease, respiratory complications, cancer, and osteoarthritis. 1,35 Although various medications have been developed to induce weight loss, many have negative secondary effects. Therefore, safe and effective treatment strategies for treating obesity are needed. We recently demonstrated that the probiotic stains HY7601 and KY1032 reduced body fat in mice and humans. 12 –15 In the present study, we investigated the molecular basis for the antiadipogenic effects of HY7601 and KY1032 using cultured 3T3-L1 preadipocytes and HepG2 liver cells. We found that treatment with both HY7601 and KY1032 reduced lipid accumulation in both cell types, especially when administered at a 1:1 ratio.
Leptin is a hormone that regulates the balance between food intake and energy expenditure through its action on central nervous system targets. 36 It is exclusively secreted by adipocytes depending on the proportion of stored triglyceride; thus, circulating leptin concentration is correlated with the extent of obesity—with higher levels in obese compared to nonobese subjects 37,38 —and can be used as an in vitro and in vivo indicator of obesity. In this study, treatment of 3T3-L1 cells with a 1:1 ratio of HY7601 and KY1032 decreased leptin expression, reflecting reduced lipid accumulation.
Adipogenesis is a tightly controlled differentiation process, regulated by sequential activations of various transcriptional factors; PPAR-γ and C/EBPα are the two main factors controlling adipogenesis during the early stages of differentiation. 39 PPAR-γ is predominantly expressed in adipose tissue and mediates the transcription of genes related to fatty acid synthesis, oxidation, transport, storage, and energy expenditure, 40,41 whereas C/EBPα directly regulates adipocyte differentiation and is expressed at high levels in obese subjects. 42,43 PPAR-γ and C/EBPα engage in cross-regulation through a positive feedback loop that induces the expression of downstream target genes such as LPL, FAS, and ACC, leading to lipid accumulation. 44 LPL is a transcription factor expressed in early stages of adipocyte differentiation; its overexpression induces lipid accumulation during adipogenesis. 45 FAS and ACC are involved in the late stage of adipogenesis. 46,47
HY7601 and KY1032 treatment reduced PPAR-γ, C/EBPα, LPL, and FAS expression in 3T3-L1 cells, suggesting that the antiadipogenic effects of HY7601 and KY1032 are mediated through inhibition of adipogenesis. A cell fractionation experiment revealed that these effects were mainly associated with the cell membrane fraction of HY7601 and KY1032. A study showed that the exopolysaccharides from Lactobacillus rhamnosus GG alleviated adipogenesis in mice 48 ; however, as there are many other effective molecules in the membrane of Lactobacillus, such as lipoteichoic acid, peptidoglycan, and S-layer protein, 49 the precise factors involved remain to be determined. Furthermore, as the cell supernatants also showed slight decreases in adipogenic gene expression, the possibility of the synergic effect of certain components in the cell membrane and cytosol may also be considered.
Mitochondria are the main organelles regulating cellular metabolism, including lipid metabolism. They are involved not only in lipolysis through induction of lipid β-oxidation, but also in anabolic processes such as lipogenesis and lipid storage in adipocytes and other tissues. 50 There is a clear correlation between mitochondrial DNA copy number (mitochondrial mass) and lipogenesis. 51,52 It is thought that mitochondria contribute to lipid storage by supplying ATP to lipid anabolic reactions 53 or through ACC-mediated transformation of acetyl-CoA exported from mitochondria into malonyl-CoA, which is required for triglyceride production. 54 We found in this study that cells treated with HY7601 and KY1032 showed a decreased mitochondrial mass and lower expression levels of PGC-1α and mtTFA, the principal regulators of mitochondrial biogenesis.
Liver cells participate in many aspects of lipid metabolism since the liver is the major site of lipid synthesis, storage, and release. Hepatic de novo lipogenesis is a fundamental biosynthetic pathway within the liver, controlled by various transcriptional factors. 55 FAS is a multifunctional protein involved in lipogenesis that synthesizes the saturated fatty acid palmitate from acetyl CoA, malonyl CoA, and reduced nicotinamide adenine dinucleotide phosphate 56 ; it plays critical roles in de novo fatty acid synthesis and long-term regulation of lipogenesis. 57 FAS expression is directly regulated by SREBP-1c, 58,59 which also regulates the expression of other genes related to fatty acid synthesis such as ACC. 60 Malonyl CoA generated from acetyl-CoA by ACC is a substrate for fatty acid biosynthesis and suppresses fatty acid β-oxidation. 61 In our study, treatment with HY7601 and KY1032 decreased FAS, SREBP-1c, and ACC expression in HepG2 cells, indicating that these LAB inhibit adipogenesis.
Using 3T3-L1 preadipocytes and HepG2 liver cells, we confirmed the antiobesity activity of HY7601 and KY1032. The mixture of two LAB decreased adipogenesis in 3T3-L1 cells by regulating the main transcription factors related to adipogenesis and by reducing mitochondrial biogenesis which inhibits ATP supply required for adipocyte differentiation. In HepG2 cells, it decreased the expression of SREBP-1c, inhibiting the central enzyme in the de novo lipogenesis pathway such as FAS and ACC. Taken together, our findings suggest that HY7601 and KY1032 may be effective for the treatment of obesity and related metabolic diseases.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.