
Abstract
Post-traumatic stress disorder (PTSD) is a traumatic stress-related psychiatric disorder stimulated by experience. Green tea has potent antioxidative properties, due, in part, to the catechin (−) epigallocatechin-3-gallate (EGCG). EGCG is an important polyphenol with advantageous effects on anxiety and depression. Nevertheless, the mechanism about the inhibition of PTSD-like symptoms of EGCG is still unidentified. We examined whether EGCG improved learning and memory deficit stimulated in rats after single prolonged stress (SPS). Rats were administrated intraperitoneally (i.p.) with EGCG for 14 successive days after the SPS process. The SPS procedure stimulated cognitive deficit in the Morris water maze test and the object recognition task, and this impairment was improved by EGCG (25 mg/kg, i.p.). Daily EGCG administration significantly decreased the freezing response to contextual fear conditioning. The administration of EGCG also significantly moderated memory-related decreases in the alternation of cAMP-response element-binding protein and brain-derived neurotrophic factor in the hippocampus. Our results suggest that EGCG alleviated SPS-stimulated learning and memory deficit by inhibiting the increase of neuroinflammation in the rat brain. In addition, EGCG reversed the alternation of allopregnanolone and progesterone in the brain, and diminished simultaneously the hypothalamic–pituitary–adrenal axis dysfunction. Thus, EGCG reversed learning and memory-related behavioral dysfunction and molecular alternation accelerated by traumatic stress and may be a useful therapeutic material for PTSD.
Introducton
P
Re-experiencing symptoms consist of aversive recollections of the intrinsic trauma in the form of recurrent memories during the day, nightmares, and flashbacks. 6 Thus, memory dysfunction and cognitive deficits also happen often in the development of PTSD. 4,7
Green tea is one of the most popular teas, and catechins such as (−)-epigallocatechin-3-gallate (EGCG), (−)-epicatechin gallate, and (−)epigallocatechin are its major active compounds. 8,9 EGCG is the most prevalent biologically specific component in green tea, 10 and it exerts neuroprotective effects against toxins, depression, ischemia, and stress-stimulated behavioral impairment. 9,11 –13 Some studies have shown that EGCG treatment reduces the radical increment in blood pressure in spontaneously hypertensive rats, and prevents most of the learning and memory dysfunction in addition to decreasing increased locomotor activity. 9 EGCG treatment decreased the lipid peroxidation and increased antioxidative effects stimulated by reserpine in the hippocampi of rats. 14 Chronic treatment with EGCG ameliorated cognition impairment in streptozotocin-diabetic rats through attenuation of oxidative stress and nitric oxide alteration. 15,16 These results have led to the hypothesis that EGCG may be effective in modulating stress-stimulated behavioral alternations such as PTSD. Therefore, EGCG may be an effective natural supplement for the protection of trauma-stimulated psychiatric conditions, including PTSD. Nevertheless, it is not known whether EGCG treatment can ameliorate single prolonged stress (SPS)-stimulated deficits in recognition of novelty and spatial learning abilities in rats. Our study estimated the effects of EGCG on PTSD-like symptoms by adjusting the SPS model in rats. After the SPS procedure, the rats were subjected to several behavioral tests such as Morris water maze (MWM), open field test (OFT), and object recognition task (ORT). Furthermore, we also investigated how these effects were associated with the molecular modulation of neuroinflammation in the neural mechanisms underlying the improvement of EGCG in the learning and memory impairment. We also studied the biosynthesis of neurosteroids and the hypothalamic–pituitary–adrenal (HPA) axis activation after EGCG treatment.
Materials and Methods
Animals
Seven-week-old male SD rats weighing 200–220 g were obtained from Samtaco Animal Co. (Osan, South Korea). The vivarium room was kept on a 12-h light/dark cycle (lights on at 9:00 am, lights off at 21:00 pm) under relative humidity of 55% ± 10% and a controlled temperature at 22°C ± 2°C. All rats were caged for 7 days to acclimatize before beginning the experimental protocol. Ethics approval was obtained from Kyung Hee University's Institutional Animal Care and Use Committee (KHUASP(SE)-15-115). All experimental procedures were performed according to the Guide for the Care and Use of Laboratory Animals.
Epigallocatechin gallate administration
EGCG (5, 10, and 25 mg/kg, body weight; Sigma-Aldrich Chemical Co. St. Louise, MO, USA) and the fluoxetine for positive drug (10 mg/kg, FLX, fluoxetine hydrochloride; Sigma) were applied intraperitoneally after exposure to SPS for 14 days. Rats were given EGCG and FLX once daily for 14 successive days after the SPS process (6–7 rats per group). The standard doses and period of EGCG used in this study was as applied in another study. 17 EGCG and FLX were liquefied in 0.9% saline before use. The entire experimental schedules are shown in Figure 1.
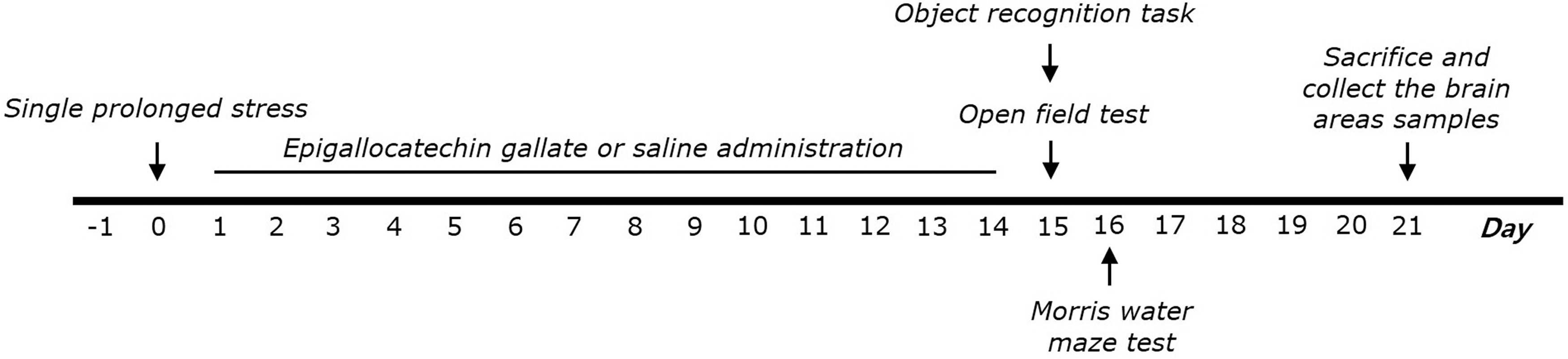
Experimental protocol for development of SPS-induced memory impairment and EGCG treatment in rats. Different groups of rats (SAL group, n = 7; PTSD group, n = 7; PTSD+EGCG5 group, n = 6; PTSD+EGCG10 group, n = 6; PTSD+EGCG25 group, n = 7; PTSD+FLX group, n = 7) were used for each experimental condition. OFT, open field test; ORT, object recognition task; MWM, Morris water maze test. EGCG, epigallocatechin-3-gallate; PTSD, post-traumatic stress disorder; SPS, single prolonged stress.
Single prolonged stress
In brief, rats were controlled for 120 min in a holder and then immediately placed in a forced swimming situation for 20 min. The rats were enabled to recuperate for 15 min and successively exposed to ether vapor until loss of consciousness. After SPS stressors, rats were housed one per cage and left undisturbed for 14 days in their home cage to enable PTSD-like symptoms to become apparent.
Object recognition task
The novel ORT is used to estimate the cognitive capability of rats. In brief, the equipment consisted of a square wooden box painted black (45 × 45 × 45 cm 3 ). The objects in the equipment were divided into familiar objects and a novel object. The familiar objects (A1 and A2) to be discriminated were two similar toys that were heavy enough, so that the rats could not move them. 18 The novel object (B) was of different shape and a different colored toy. To habituate, the rats were left in the object recognition box for 10 min. The test phase started 24 h after the habituation. Rats were exposed to the equipment with two familiar objects for 5 min. During the test phase, rats housed in the testing chamber were exposed to one novel object (B) and one of the familiar objects for 5 min. The exploration (sniffing) time for the novel and familiar objects is measured. The discrimination index is calculated as discrimination between the familiar and the novel objects accurate for exploration. It is expressed as: (time spent on novel object − time spent on familiar object)/(time spent on novel object + time spent on familiar object). 18
MWM test
MWM test was made of a spatial probe test and a place navigation test. The MWM consisted of a circular pool (200 cm diameter and 50 cm deep). The pool contained water maintained at a temperature of 22°C ± 2°C. The escape platform (15 cm diameter) was located 1.5 cm below the water in one of the four sections of the pool. The hidden platform trial for acquisition test and probe trials for retention test were monitored by a video camera mounted on the ceiling, and data were analyzed using a tracking program (S-MART: PanLab Co., Barcelona, Spain). The rats performed three training trials per day for 5 successive days. Each trial was terminated when the rat found the platform or after 180 sec. On day 6, the platform was removed. In the probe trial, each trial was 1 min in duration. The swimming path length, swimming speed, and time spent in the target quadrant were analyzed.
Open field test
In the dimly lit room, rats were exposed singly in a square black plexiglass apparatus (60 × 60 × 30 cm) and tracked by a video tracking system for 5 min. Locomotion was analyzed by the distance and speed of movements and observed by a computerized video-tracking analysis program S-MART (PanLab Co.). The number of rearing was also manually scored by examining the records in the OFT.
Contextual fear conditioning and extinction
In segregated experiment, rats that did not undergo MWM testing (N = 4–5 for each group) were exposed for contextual fear conditioning and extinction, beginning 14 days after SPS procedure. The apparatus was made of a brightly lit acrylic chamber (30 × 26 × 22 cm) with grid floor made of stainless steel rods. Briefly, on day 1, rats were habituated to the chamber for 5 min and received a single pairing of a tone (30 sec, 5 kHz, 75 dB). After then, rats received five electric foot shocks (2 sec, 2 mA) that were randomly divided over a 10-minute period. All the animals were exposed to situational reminders (fear memory was reactivated by placing rats in the same chamber and presenting the tone without the shock) for 5 min on days 7 and 14, respectively. The percentage of freezing = (freezing time)/(total time). 19
Corticosterone, corticotropin-releasing hormone, adrenocorticotropic hormone, cAMP-response element-binding protein, brain-derived neurotrophic factor, allopregnanolone, progesterone, and proinflammatory markers measurement
Twenty-one days after the SPS procedure, corticosterone (CORT), adrenocorticotropic hormone (ACTH), and corticotropin-releasing hormone (CRH) in the plasma, cAMP-response element-binding protein (CREB), brain-derived neurotrophic factor (BDNF), progesterone (P4), allopregnanolone (ALLO), interleukin-1β (IL-1β), and tumor necrosis factor-α (TNF-α) concentrations in the brain for assay were applied as in a previous study. 20 Rats were deeply anesthetized through isoflurane inhalation (1.2%) and killed after behavioral test (4 rats per group). The plasma was rapidly collected through the abdominal aorta. The prefrontal cortex and hippocampus were quickly removed from the rat brains. The CORT, CRH, ACTH, CREB, BDNF, P4, ALLO, IL-1β, and TNF-α concentrations were measured by a competitive enzyme-linked immunosorbent assay (ELISA) using a CORT antibody (Novus Biologicals, LLC., Littleton, CO, USA), a CRH antibody (Novus Biologicals), a ACTH antibody (Novus Biologicals), a CREB antibody (Thermo Fisher Scientific, Waltham, MA, USA), a BDNF antibody (R&D Systems, Minneapolis, MN, USA), a P4 antibody (Abcam, Cambridge, MA, USA), a ALLO antibody (Thermo Fisher Scientific), an IL-1β antibody (Abcam), and a TNF-α antibody (Abcam) according to the manufacturer's protocol.
Total RNA isolation and reverse transcription–polymerase chain reaction analysis
The expression levels of IL-1β, TNF-α, CREB, and BDNF mRNA were evaluated by reverse transcription–polymerase chain reaction (RT
Statistical analysis
All data are expressed as mean ± SEM. The data were analyzed using SPSS 13.0 (Chicago, IL, USA) and by multiway of analysis of variance (ANOVA) and Tukey's post hoc tests. Statistical significance was set at P < .05 and is indicated using symbols in all figures.
Results
Effects of EGCG on SPS-stimulated memory deficits
The novel object recognition for learning and memory function was indicated by means of the exploration (sniffing) times of familiar and novel objects and by computation of the discrimination index by the ORT (Fig. 2A, B). No significant differences in the exploration (sniffing) times on the training day were detected between experimental groups, by two-way ANOVA, which offers no external stimuli or reinforcement [F(5,39) = 1.857, P = .128]. There was no differential preference for a specific object. Analyses of exploration times for novel objects by two-way ANOVA revealed significant differences between groups [F(5,39) = 37.059, P < .001]. The exploration (sniffing) times of novel object in the PTSD group were significantly decreased after the SPS procedure, compared with control (P < .001), during testing period. The PTSD+EGCG25 group showed increased exploration (sniffing) times compared with the PTSD group (P < .001). Post hoc examination with Tukey test revealed that the discrimination index of PTSD group was significantly decreased compared with the saline-treated (SAL) group (P < .05). In addition, the discrimination index of the PTSD group that received EGCG (25 mg/kg) was significantly increased compared with the PTSD-only group (P < .05). The PTSD+EGCG25 group in recovery of object recognition memory was approximated to those in the FLX-treated rats. In MWM test, SPS-stimulated rats were weak to learn during acquisition trial and retention trial. However, the memory abilities of SPS-stimulated rats were significantly impaired compared with the SAL group (P < .01 on days 3 and 5). A significant reduction in escape latency was shown in the PTSD+EGCG25 group compared with that in the PTSD group (P < .05). In the probe trial of the MWM test, treatment of PTSD significantly decreased the number of platform-crosses compared with that of the SAL group (P < .05). Compared with PTSD group, EGCG-treated rats presented more number of platform-crosses (P < .05). The swimming latency in the PTSD+EGCG25 group was approximated to that in the PTSD+FLX group. The PTSD group did not differ significantly from the other groups in the mean swimming speed. SPS procedure carefully impaired spatial cognitive memory in the water maze test. EGCG treatment displayed attenuated SPS-stimulated memory damage in the MWM test.

Effects of EGCG administration on recognition memory were assessed using the novel ORT measuring the time taken to sniff familiar and novel objects during a 3-min choice trial
The PTSD-stimulated rats demonstrated considerable motor function as measured by the locomotion and exploration as total number of rearing in the OFT. We found no significant difference among PTSD-stimulated, saline-treated, and EGCG-treated rats in terms of locomotor activity [F(5,39) = 0.356, P = .875] or the total number of rearing [F(5,39) = 0.779, P = .572].
Effects of EGCG on SPS-stimulated contextual freezing behavior
The freezing time was significantly increased after exposure to SPS in the contextual freezing measurement (P < .05 on days 7 and 14; Fig. 3). Although repeated EGCG administration failed to reverse the increase in freezing time in the stress-stimulated rats on day 7, the percentages of freezing times were significantly decreased in the group treated with EGCG at 25 mg/kg on day 14 (P < .05). These results show that the actuality of a continuous fear response to the context was linked to traumatic experiences, and repetitive EGCG treatment relieved contextual freezing behavior in the rats.
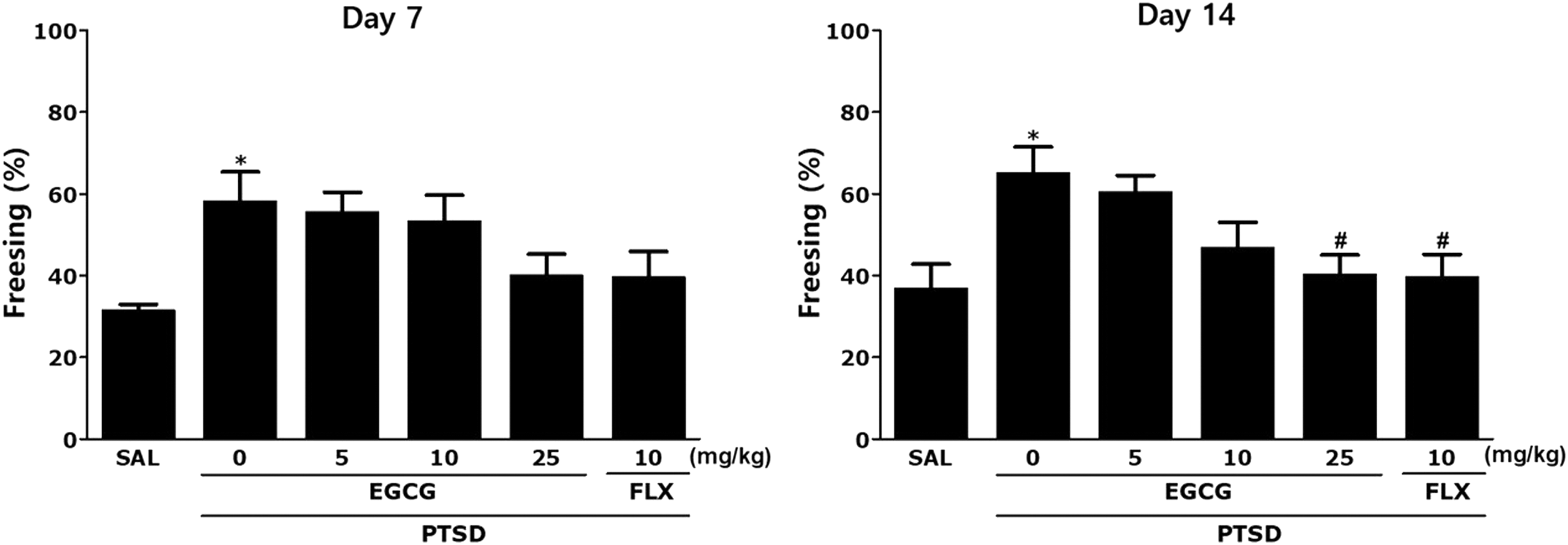
Effects of EGCG on freezing behavior after exposure to SPS in rats The percentages of freezing time were determined on days 7 and 14. *P < .05 versus SAL group; # P < .05 versus PTSD group.
Effect of EGCG on SPS-stimulated alternations in plasma CORT, CRH, and ACTH levels
ELISA analysis indicated that SPS exposure during 14 days significantly increased the plasma CORT, CRH, and ACTH levels in the rats by 237.01%, 321.48%, and 234.61%, respectively, compared with those in the SAL group (P < .05 and P < .01; Fig. 4A–C). But, administration of EGCG decreased the SPS-stimulated increase in plasma CORT levels, although this result was only minimally statistically significant. But, the administration of EGCG decreased the SPS-stimulated increase in plasma CRH and ACTH levels compared with the PTSD group, respectively (P < .05). These results indicated that the inhibition of PTSD-like symptoms of EGCG was linked to decreased HPA axis stress hormone levels. The suppression of the increase in plasma CORT, CRH, and ACTH levels by EGCG treatment also well showed the potential of therapeutic agents in the SPS-stimulated PTSD-like symptoms in rats.
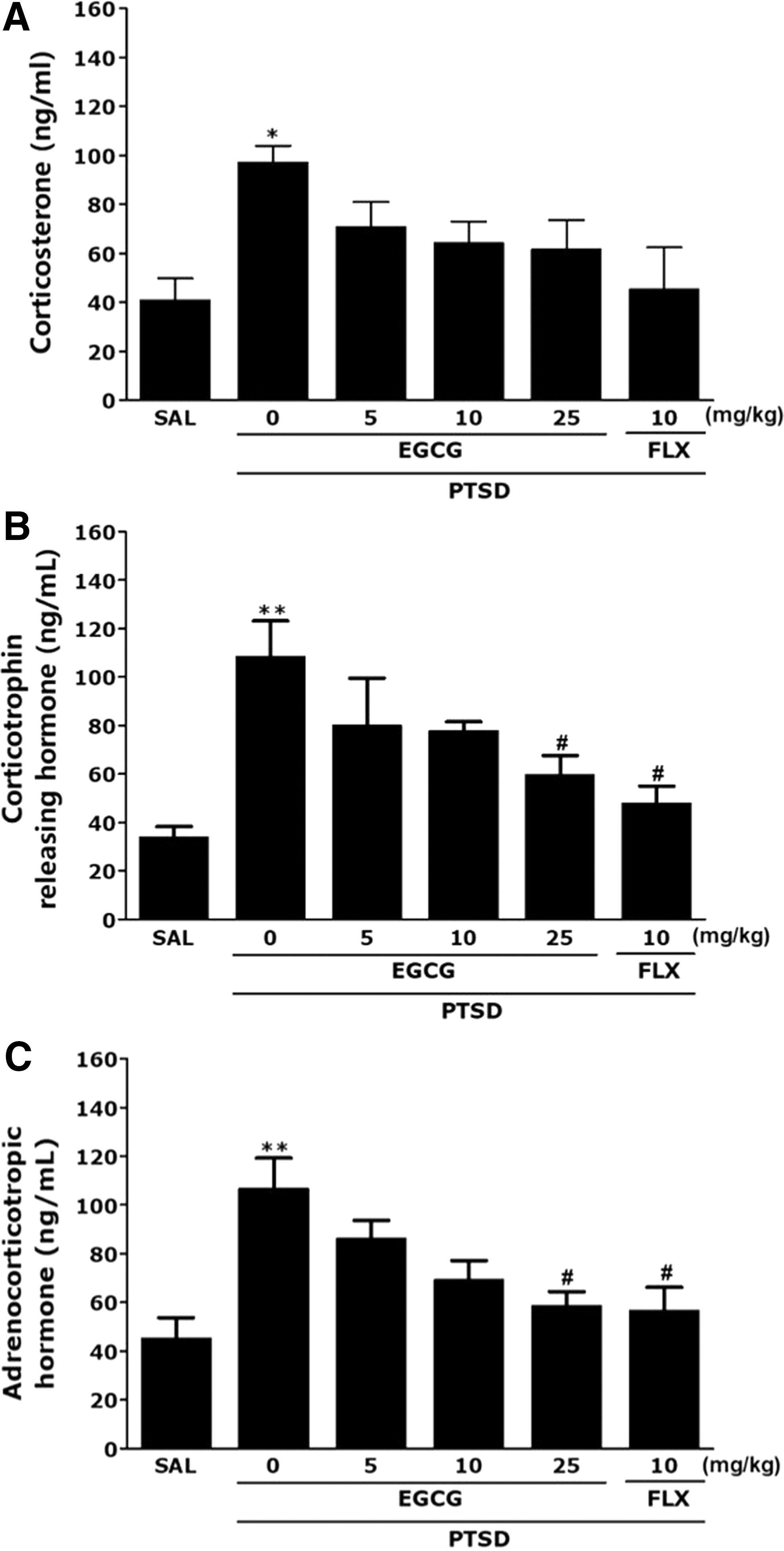
Effects of EGCG administration on plasma CORT
Effects of EGCG on SPS-stimulated alternations in CREB and BDNF in the hippocampus
The post hoc test results revealed significantly decreased concentrations of CREB and BDNF in the hippocampus of PTSD groups compared with those of the SAL group (P < .01 and P < .05; Fig. 5A, B). Administration of EGCG significantly increased the SPS-stimulated decrease in BDNF level in the hippocampus compared with that in the PTSD group (P < .05).

Effects of EGCG administration on CREB and BDNF protein levels
To research the effect of EGCG on the expression levels of neurotrophic factors in the hippocampus of rats damaged by PTSD, the mRNA levels of CREB and BDNF were analyzed using RT-PCR whereas the mRNA of CREB and BDNF in the PTSD group significantly decreased compared with that of the SAL group (P < .05). The decreased expression of CREB and BDNF mRNA in the PTSD group was significantly re-established to levels similar to those in the SAL group by 25 mg/kg of EGCG (P < .05). Our results show that CREB and BDNF levels in the hippocampus of rats taking 25 mg/kg of EGCG were similar to those of rats taking 10 mg/kg of FLX.
Effects of EGCG on SPS-stimulated levels of neuroinflammatory mediators in the hippocampus
The post hoc test results indicated significantly increased levels of IL-1β and TNF-α in the hippocampus of PTSD groups compared with those in the SAL group (P < .01 and P < .05; Fig. 6A, B). Administration of EGCG significantly decreased the SPS-stimulated increase in IL-1β and TNF-α levels in the hippocampus compared with those in the PTSD group (P < .05).
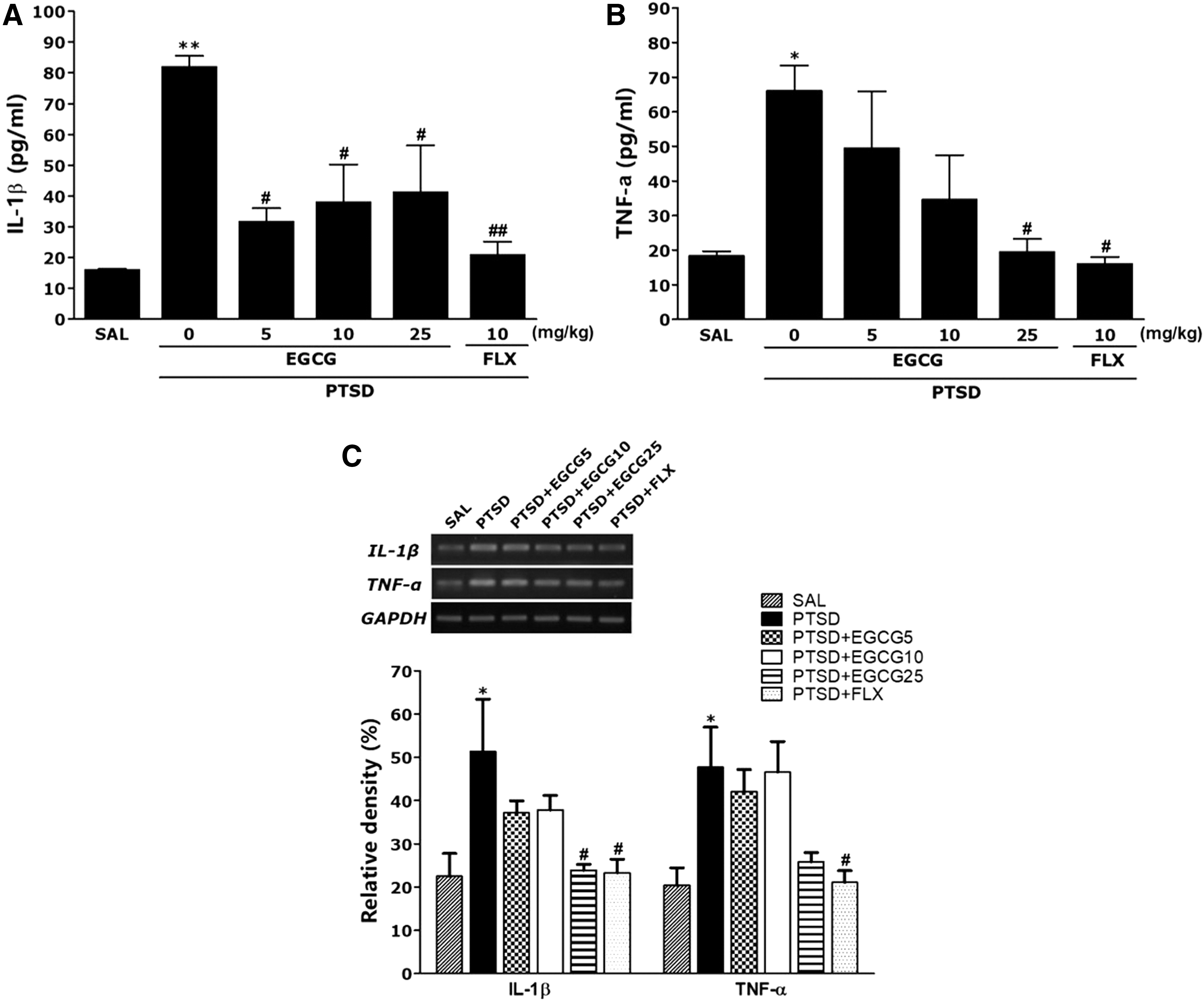
Effects of EGCG administration on IL-1β and TNF-α protein levels
To investigate the effect of EGCG on the expression of neurotrophic factors in the hippocampus of rats damaged by PTSD, the mRNA expression levels of IL-1β and TNF-α were analyzed using RT-PCR, whereas the mRNA level of IL-1β and TNF-α in the PTSD group significantly increased compared with that of the SAL group (P < .05). The increased expression level of IL-1β mRNA in the PTSD group was significantly re-established to levels similar to those in the SAL group by 25 mg/kg of EGCG (P < .05).
Effects of EGCG on SPS-stimulated alternations in P4 and ALLO in the prefrontal cortex and hippocampus
The post hoc test results showed a significantly decreased concentration of P4 in the prefrontal cortex and hippocampus of PTSD groups compared with those of the SAL group (P < .01; Fig. 7). However, administration of EGCG significantly increased the SPS-stimulated increase in P4 level in the hippocampus compared with that in the PTSD group (P < .05). The post hoc test analysis showed a significantly decreased concentration of ALLO in the prefrontal cortex and hippocampus of PTSD groups compared with that in the SAL group (P < .01 and P < .05; Fig. 7). However, administration of EGCG significantly increased the SPS-stimulated increase in ALLO level in the prefrontal cortex and hippocampus compared with that in the PTSD group (P < .05).

Effects of EGCG administration on the levels of progesterone and allopregnanolone in the prefrontal cortex and hippocampus. *P < .05, **P < .01 versus the SAL group, # P < .05 versus the PTSD group.
Discussion
Our outcomes clearly indicated that EGCG treatment significantly improved spatial memory and learning disorders stimulated by the SPS procedure, as evidenced by rat behavior in the MWM test and ORT. SPS-stimulated rats also revealed increased proinflammatory cytokine and decreased hippocampal BDNF levels. Moreover, we found that EGCG was related to the prevention of neurosteroid biosynthesis in the brain and normalization of HPA axis dysfunction.
In a previous study of the memory impairment animal model, 25 mg/kg EGCG treatment for 4 weeks showed a neuroprotective effect. 17 Thus, the same dose of EGCG (25 mg/kg for 2 weeks) was used in this experiment, which is ∼7.5 times more than the typical dose for human consumption. Human consumption of green tea daily is ∼2 mg/kg (1.2 g × 1% yield from green tea leaf per 70 kg). 21 Each rat's average water consumption per day was about 3–4 mL. Therefore, the total dose of EGCG intake was ∼15 or 60 mg per rat. The total dose of EGCG that reached the brain was ∼2–3 nmol/g according to a published study. 22 We investigated the dose-dependent effects of EGCG and found that 25 mg/kg of EGCG was the most adequate dose for preventing SPS-stimulated negative effects in the MWM test and ORT. In this study, the optimal dose was previously reported.
To discover the effects of EGCG on two different types of memory, spatial learning and memory and process of recognition, behavioral tests such as ORT and MWM test, respectively, were used. Our results indicated that cognitive memory was damaged under SPS-stimulated situations. This impairment was significantly more noticeable after SPS exposure, as verified by the no significant differences in the exploration time for familiar objects, deterioration of process of recognition in the exploration time for novel object, and reduction in the index of stimulus recognition. These results showed an intense stimulation and damage in the brain after exposure to memory impairing events, contributing to decreases in episodic memory and recognition ability. Therefore, the SPS procedure well showed deterioration of memory function of recognition in the novel object and the index of stimulus recognition. Administration of EGCG significantly increased the exploration time for novel object and ameliorated the impaired object recognition memory after the SPS procedure. The MWM is a hippocampal-dependent spatial memory test, used specifically to demonstrate cognitive impairment and to investigate constant spatial learning ability and reference memory function in rats. The SPS-infused rats exhibited significantly longer escape latencies to reach the platform and presented damages in spatial learning during the acquisition trial of the MWM test. In the SPS-stimulated rats that received EGCG, the rats exhibited faster learning and shorter escape latencies than the nontreated group. Furthermore, the SPS-infused rats without EGCG administration performed poorly in successive testing in probe trials 24 h after acquisition compared with control. EGCG improved these behavioral alternations and re-established spatial learning and memory and recognition of novelty in SPS-stimulated rats. Long-term treatment with the FLX showed similar effects. Thus, the results in this study confirm our hypothesis that EGCG ameliorates spatial memory impairment stimulated by the SPS procedure.
Significant individual differences in total number of rearing and locomotor activity were not shown between the groups, suggesting that EGCG did not affect psychomotor ability, motor damage, or sensory-motor performance in the OFT. In addition, memory impairment observed in the SPS-stimulated rats was probably not contributed to differences in their locomotion activities. Consequently, the alterations in behavioral performance in the MWM test may be associated with improved memory, not differences in psychomotor function or active responses.
The SPS-stimulated freezing behavior also associated that PTSD is typified by an exaggerated reaction to a mild stressor or the reminder of the trauma to which the response is suitable for the original traumatic condition. 23 Therefore, SPS exposure led to the acquisition of conditioned fear responses to trauma-related stimuli. 24 Thus, the present results support that EGCG could ameliorate fear memory stimulated by traumatic stress.
In in vivo system, the enforced elevation of CORT concentrations can influence cognition impairment by learning and memory ability, and this might be associated with the progression of traumatic stress in humans. 20 EGCG prevented the SPS-stimulated increase in CORT levels, suggesting that EGCG inhibits stress-stimulated HPA axis dysfunction and associated behavioral deficit, as well as decreases in CREB and BDNF expression in the hippocampus. We indicated that EGCG treatment prevented the increase in HPA axis inhibition. Therefore, our results indicated that the effects of EGCG are arbitrated by an increase in BDNF level through the CREB modulations, and might be associated with an improvement in the performance on memory-related tests and neuronal function.
In addition, we investigated the levels of CREB and BDNF, an important factor related to cognition and memory. 25 CREB and BDNF have been reported to be downregulated during chronic stress in the hippocampus of rats, and memory-related drug treatment prevented the decrease in CREB and BDNF levels stimulated by stress. 25 An association of impaired cognition and memory with reduced levels of CREB and BDNF has also been reported in patients with PTSD. 25,26 To verify EGCG-related action mechanisms of learning and memory betterment, the effects of EGCG on BDNF level in the hippocampus, estimated to be an essential role neuromodulator in the production of learning and memory, were examined. SPS-stimulated memory impairment significantly reduced BDNF mRNA expression levels in the hippocampus and resulted in poor performance in hippocampus-dependent tasks. 4,27 CREB is also believed to play a decisive role in the production of memories. 4 However, in this study, no significant individual differences in CREB levels were observed after treatment with EGCG, which suggests that EGCG did not affect CREB level. However, SPS-stimulated cognition impairments are associated with significant decrease in CREB mRNA level in the hippocampus as well as poor performance on hippocampus-dependent spatial test and process of recogntion. 25 In this study, EGCG treatment significantly reversed SPS-stimulated decreases in CREB mRNA expression, which suggests that the useful results of EGCG were associated with increases in CREB mRNA expression that may be mediated to performance in memory test and enhanced neuronal function. Unfortunately, the present findings indicate that there is no correlation between gene and protein function.
This study also indicates that SPS significantly increased the expressions of IL-1β and TNF-α in the hippocampus, essentially causing neuroinflammatory action in the brain. Hippocampus deficits commonly stimulated memory impairment and reduction of learning ability and stimulated poor reinforcement of declarative memory in animals and humans. 26 Thus, continuous increases in expression of proinflammatory cytokines stimulated by SPS have been closely connected to psychiatric diseases related to impaired working memory. 25 Alternation of IL-1β and TNF-α levels is regulated dynamically by several immune cells during inflammatory action in the hippocampus. 27 In addition, EGCG continually decreased SPS-stimulated IL-1β and TNF-α protein, finally resulting in restoration of the chronic inflammation and continuous brain dysfunction. 28,29 In conformity with the memory dysfunction and inflammation hypothesis, PTSD is the result of chronic inflammation and irreversible and selective dysfunction in the brain. 28,29 Therefore, our results suggest that the anti-inflammatory efficacy of EGCG significantly reverses the SPS-stimulated deficits in learning and memory function and increased levels of proinflammatory mediators such as TNF-α and IL-1β.
Altered levels of P4 and ALLO have also been involved as one of the most powerful factors for PTSD pathological condition in the hippocampus and prefrontal cortex, which is associated with emotional processing, fear conditioning, and explicit memory. 30 –32 In addition, ALLO, a major metabolite of P4, has been associated with decreased levels of context-related fear memory in the PTSD. 33 These findings indicated that, like FLX, EGCG significantly inhibited the reduction in the expressions of P4 and ALLO in the hippocampus and prefrontal cortex.
In conclusion, the present results indicate the hypothesis that EGCG ameliorated SPS-stimulated memory and behavioral deficits in rats. Administration of EGCG significantly weakened the problems of SPS-stimulated damage significantly, as suggested by ameliorated cognitive ability and object recognition memory during behavioral tests, prevented decreases in BDNF levels, and normalized HPA axis dysregulation. EGCG also inhibited SPS-simulated alternation of proinflammatory cytokines such as IL-1β and TNF-α in the hippocampus. In addition, our results also reveal that EGCG produces a therapeutic effect on PTSD-like stress response that was led by modulation of the HPA axis and biosynthesis of neurosteroids. Therefore, our results suggest that EGCG may have efficacy as a functional food good material for amelioration of object recognition memory, improvement of learning and memory, and prevention of neuronal modulations related to PTSD.
Footnotes
Author Disclosure Statement
The authors declare no potential conflicts of interest.