
Abstract
Diet and exercise are recommended both as a prophylactic and as a therapeutic approach for patients with established coronary artery disease. We previously reported that sesame oil (SESO) and its aqueous extract (SOAE) showed antiatherosclerotic and anti-inflammatory properties. We also observed that genes involved in reverse cholesterol transport (RCT) might be activated. In this study, we tested whether post-treatment with SESO or SOAE would reduce preexisting atherosclerosis by enhancing RCT. Female low-density lipoprotein receptor knockout (LDL-R−/−) mice were fed an atherogenic diet for 3 months, followed by post-treatment with either control or SESO or SOAE for 1 month. Plasma lipids and atherosclerotic lesions were quantified at the end of the study. RNA was extracted from the aortic tissues and used for real-time polymerase chain reaction analysis. SESO and SOAE post-treatment significantly reduced atherosclerotic lesions in LDL-R−/− mice compared to controls. No significant change in plasma cholesterol, triglyceride, or LDL cholesterol levels was observed. Aortic gene analysis showed that the SESO/SOAE post-treatment reduced inflammatory gene expression and induced genes involved in cholesterol metabolism and RCT. This is the first study that demonstrates that post-treatment with SESO and SOAE could be an effective treatment for preexisting atherosclerosis and inflammation. The study also may suggest that reducing inflammation might be conducive to an accelerated regression of lesions.
Introduction
C
Sesame oil (SESO) has been reported to help reduce high blood pressure and lower the amount of medication needed to control hypertension. 5 –7 Other beneficial effects include reduction in plasma cholesterol, low-density lipoprotein (LDL) cholesterol, and triglyceride (TRG) levels. 8 Our earlier studies of sesame oil and its aqueous extract (SESO/SOAE) diet-fed LDL-R −/− mice showed significantly reduced lipid levels, inflammation, and atherosclerotic lesions compared to high-fat diet-fed control animals. 9,10 In this study, we tested whether SESO or SOAE post-treatment would facilitate reverse cholesterol transport (RCT) and “promote” regression of atherosclerosis.
Materials and Methods
Chemicals
All chemicals were purchased from Sigma (St. Louis, MO, USA).
Preparation and analysis of aqueous extract
SOAE was prepared and used as described previously. 11 Briefly, SOAE was prepared using SESO and distilled water. Aqueous portion was separated by filtration and lyophilized. Lyophilized sample was reconstituted with pyrogen-free water. Absence of lipid portion and residual protein was confirmed by thin layer chromatography (TLC) and sodium dodecyl sulfatepolyacrylamide gel electrophoresis (SDS-PAGE) analysis. In addition, absence of possible endotoxin contamination of SOAE was confirmed using Limulus assay.
Animals
Twenty-five 4-week-old female LDL-R −/− mice on a C57BL/6J background (B6.129S7-Ldlr tm1Her/J strain), weighing 18–20 g, were obtained from Jackson Laboratory (Bar Harbor, ME, USA). After 2 weeks of acclimatization, five animals (n = 5) were sacrificed to obtain baseline parameters. The remaining animals were fed an atherogenic (high fat) diet for 3 months and were divided into three groups followed by 1 month post-treatment as group 1 on a normal diet, group 2 was on SESO diet, and group 3 was on normal diet supplemented with SOAE (0.75 mg/day/mouse). The animals were regularly monitored, and a weekly record of body weight was maintained up to 120 days. All procedures were performed according to a protocol approved by the Institutional Animal Care and Use Committee of the University of Central Florida.
Diet
The normal purina diet (TD.150278) with 1% soybean oil, normal diet reformulated with either 1% SESO (TD.150515) or SOAE (prepared in-house: 0.75 mg/mouse/day), and an atherogenic diet with 2% cholesterol and 17% high fat (TD.04287) were purchased from Harlan Teklad (Madison, WI, USA). The composition of the atherogenic diet was identical to that described previously. 12 Diets were stored at 4°C to avoid oxidation.
Collection of plasma and organs
After 15 weeks of feeding, mice were fasted overnight and anesthetized with 1–2% isoflurane. Fasting blood samples were collected into EDTA tubes by heart puncture. Plasma was separated as described previously 12 and stored at −80°C. The liver was perfused with ice-cold phosphate buffered saline (PBS), weighed, and the tissue stored for RNA isolation.
Isolation and quantification of aortic lesions
Isolation of the aorta and quantification of aortic lesions were performed as described previously. 9,10,12 Detailed en face pictures of the aorta along with a ruler were obtained using a digital camera. Lesion areas were marked on photographs via direct microscopic observations. The lesion area was quantified using ImageJ software. 13 The ruler in the pictures was used to determine the pixel to mm2 conversion factor. After imaging, aortas were stored at −80°C for RNA extraction.
Plasma lipid analysis
Plasma lipid profiles of total cholesterol, triglyceride (TRG), high-density lipoprotein cholesterol (HDL-C), and LDL cholesterol (LDL-C) and glucose levels were determined using a Cholestech L*D*X analyzer (Cholestech Corp., Hayward, CA, USA).
cDNA synthesis and real-time polymerase chain reaction
Total RNA from aortic tissue was isolated using TRIzol™ reagent. Total RNA of 1 μg was then reverse-transcribed into cDNA using the SuperScript™ III First Strand Synthesis system (Invitrogen, Carlsbad, CA, USA). cDNA (50 ng) samples were used to perform quantitative real-time polymerase chain reaction (RT-PCR) by iQ™5 iCycler Multicolor Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) with SYBR Green (Invitrogen). Mouse oligonucleotide primers for RT-PCR were purchased from Invitrogen. PCR was carried out with ABCA1, ABCG1, CD4, CD36, CD68, DOCK2, LOX1, MCP-1, MMP9, P-selectin, and SR-A1, with mouse-specific primers (Supplementary Table 1; Supplementary Data are available online at
Statistical analyses
Values are presented as mean ± standard deviation (SD), and statistical analyses were performed using Student's t-test, with P < .05 as the level of significance. Significance between groups was calculated using one-way ANOVA-bonferroni multiple comparison test using Prism Pad software (version V), with P < .05 considered significant.
Results
Body weight analysis in mice
A slight significant decrease in body weight (∼6%) was observed in SOAE diet treated animals compared to the control and SESO diet treated animals (Supplementary Fig. S1A, B). No significant changes were observed in liver weight in all the groups.
No significant changes in plasma lipid levels in SESO/SOAE diet post-treated animals
No significant difference was identified in visual observation of plasma following centrifugation in between the groups (data not shown). As shown in Figure 1A, B, the plasma lipid profile analysis showed an insignificant decrease in TRG and glucose levels in SOAE diet-fed animals compared to controls and SESO diet-fed animals. An insignificant increase in HDL cholesterol levels was observed in SESO diet animals. Elevated levels of HDL cholesterol in SESO diet-fed animals suggest its pronounced effect on HDL.
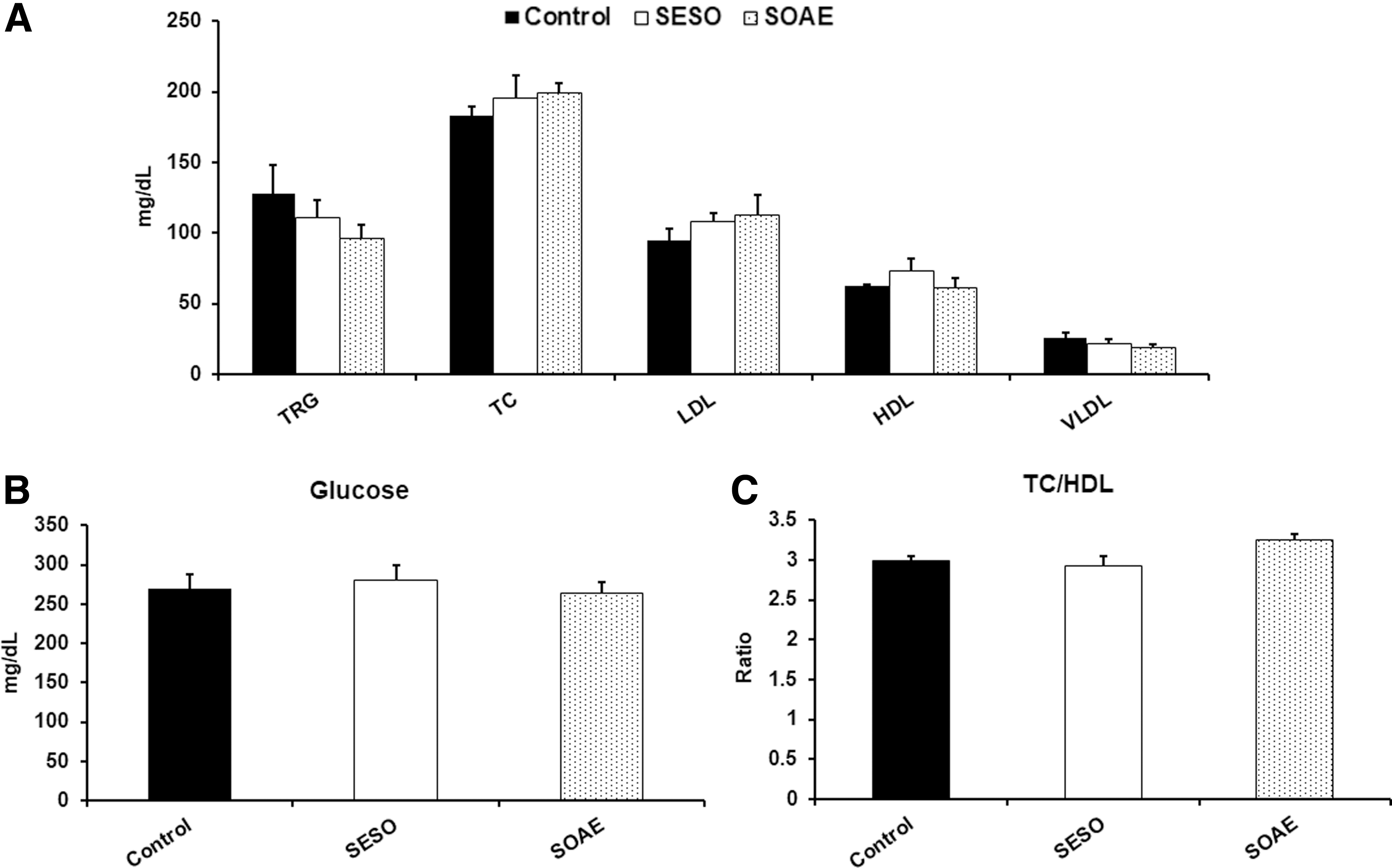
Plasma lipid levels in SESO/SOAE diet-treated animals.
Regression of atherosclerotic lesions in SESO/SOAE treated animals
Atherosclerotic lesion regression was analyzed in experimental animals, and the extent of lesion regression was quantified by measuring the lesion surface area. We observed a significant increase in lesion regression in SESO/SOAE treated animals compared to control animals. Figure 2A shows significant lesions in control animals which had prominent lesions in the aortic arch, and in some instances, lesions extended up to the abdominal aorta. These lesions were generally not present in those animals treated with SESO/SOAE (Fig. 2B, C). Quantitation of the lesions also showed that the lesion sizes (in mm2) of SESO/SOAE treated animals were significantly reduced compared to those of the controls (Fig. 2D). Average lesion area of all the animals is represented in Figure 2E. Overall, the SESO/SOAE treated animals showed that lesion regression was induced by 30–50% compared to control animals (Fig 2F; mean ± SD (mm2), **P < .01; ***P < .001).

Regression of atherosclerotic lesions in SESO/SOAE diet-treated animals. Representative images of atherosclerotic lesions in
Gene expression in mouse aorta
Aortic gene expressions were analyzed using real-time PCR. A nonsignificant increase in RCT related ABCA1 gene expression was observed in SESO/SOAE post-treated animals, whereas significantly reduced levels of ABCG1 expression (Fig. 3A) were observed compared to control animals. Similarly, and surprisingly as pre-treatment (unpublished) 1-month post-treatment with SESO/SOAE significantly reduced the expression of monocyte/macrophage markers and scavenger receptors CD68 and SR-A1 (Fig. 3B), whereas CD36 was reduced in SOAE diet-fed animals only. In addition, a significant reduction was observed in inflammatory markers such as CD4, MCP1, and P-selectin (Fig. 3C) in experimental animals compared to control animals. A significant difference in DOCK2 and LOX1 expressions was observed between control and experimental animals in the aortic arch segments (Fig. 3D). SESO/SOAE post-treatment decreased CD68 levels in the aortic arch by 48% and by 77% (*P < .05), respectively.
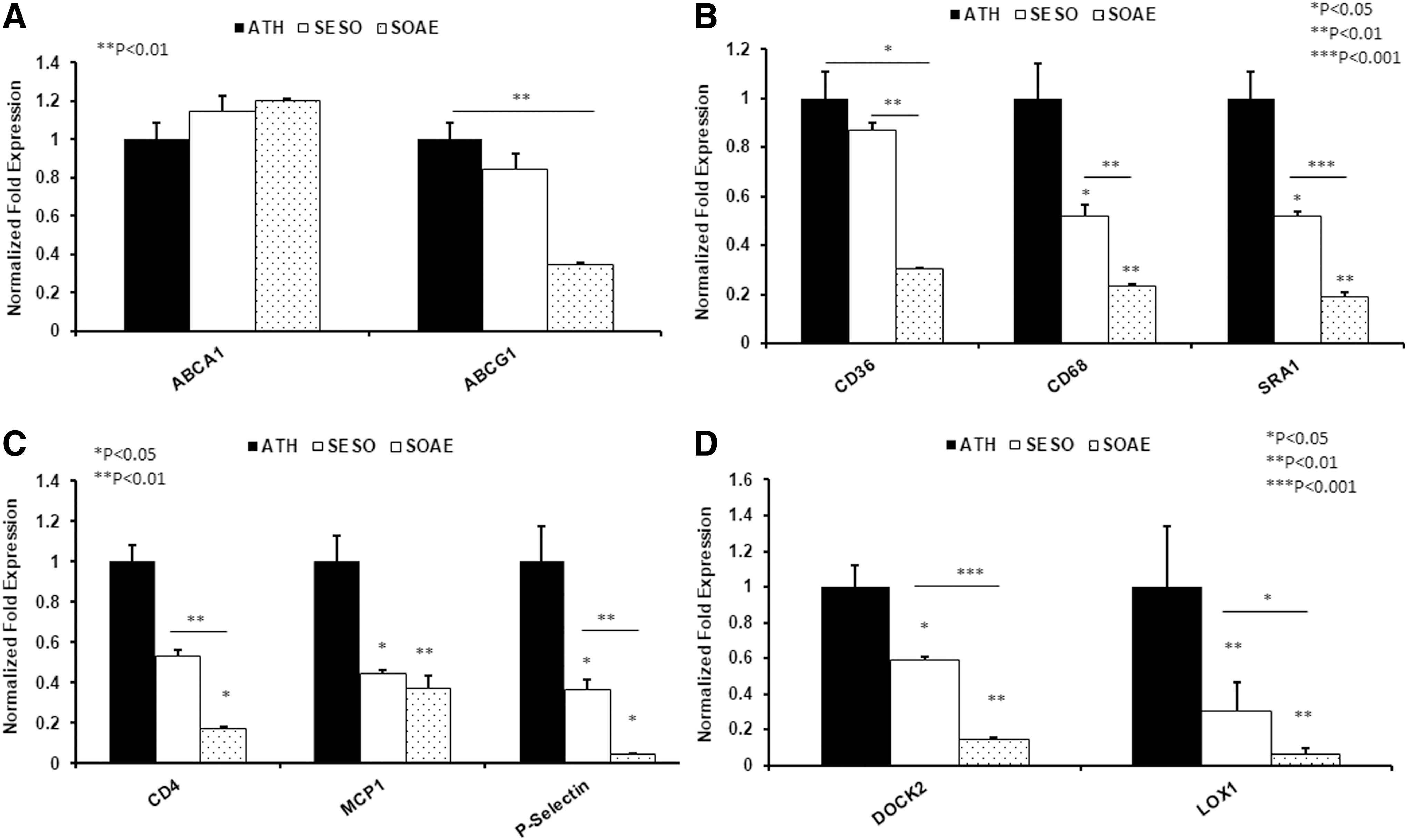
Gene analysis from mice aorta. mRNA level of several genes was analyzed in aorta of LDL-R
−/− mice after 3 months of feeding with high fat diet followed by 1 month normal/SESO/SOAE diet. Bar diagrams represent
Discussion
In the present study, we show for the first time that the SESO/SOAE diet causes reduction of preexisting aortic atherosclerotic lesions in LDL-R −/− animals in which lesions were induced by high fat diet. To our knowledge, this is the first study to demonstrate a reduction in preexisting lesions by SESO/SOAE diet in an LDL-R deficient animal model.
Pioneering studies by Rudel and coworkers have clearly established that dietary PUFA are more effective in preventing atherosclerosis than MUFA, despite the putative resistance of MUFA to oxidation. 14,15 The mechanism(s) by which PUFA decrease atherosclerosis has been studied extensively. As a result, numerous mechanisms have been proposed. These include (1) increased conversion of cholesterol to bile acids, (2) increased RCT, and (3) decreased inflammation. It is likely that all these play a vital role in the effects seen in intact animals when SESO/SOAE diet is fed. Although no mechanism has been reported, by acting as ligands for peroxisome proliferator-activated receptors (PPARs), they may activate orphan receptors and thus facilitate RCT. Similarly, they may suppress inflammation via PPAR gamma-mediated effects.
Enhanced RCT with increased expression of ABCA1 in SESO and SOAE post-treated animals was observed compared to control animals, whereas significantly reduced levels of ABCG1 expression were observed in these animals. The increased ABCA1 and reduced ABCG1 in SOAE post-treated animals suggest that SESO/SOAE might promote enhanced RCT toward lipid poor APOA1 than to intact HDL. PUFA are known to increase the conversion of cholesterol to bile acids by way of cholesterol 7alpha hydroxylase (CYP7A1). Evidence suggests that the orphan nuclear receptors farnesoid X receptor (FXR) and liver X receptor (LXR) are negative and positive regulators of CYP7A1 transcription. 16,17 Thus, a loop consisting of increased conversion of cholesterol into bile acids would be expected to increase, as well as an increase in RCT mediated by LXR. The potent ligands for FXR activation are the bile acids chenodeoxycholic, deoxycholic, and lithocholic acid, while the ligands for LXR activation are oxysterols. We previously reported that oxidized fatty acids “mimic” bile acids in their properties 18 and could potentially affect FXR, thus lowering the conversion of cholesterol to bile acids. Conversely, the components of SESO/SOAE could activate LXR and promote RCT.
Reduced mRNA levels of SR-A1 were observed in aortic lesions of SESO/SOAE diet-fed animals, suggesting the ability of SESO/SOAE to prevent further foam cell formation. SESO contains lignans, which are known to complex cholesterol from the gut and prevent cholesterol absorption. Similarly, SOAE might be contributing to the prevention of cholesterol absorption. This might be one of the reasons why there were no changes in blood lipids. We also observed a significant decrease in the mRNA level of CD68, a marker for monocyte/macrophages, in SESO/SOAE diet animals compared to normal diet-fed animals. The decrease of CD68 in the aortic arch area containing lesions is a confirmation that the presence of foam cell forming macrophages in SESO/SOAE animals was reduced. In addition, reduced LOX1 expression also confirmed the further accumulation of macrophage lipids. An insignificant decrease in MMP9 gene expression was observed (data not shown), whereas a significant decrease in DOCK2 expressions was observed in both SESO/SOAE diet-fed animals. MMP9 and DOCK2 play a key role in smooth muscle cell migration and proliferation and have been considered as major pathological factors in atherosclerosis. Similarly, a significant reduction in the levels of CD4, MCP-1, and P-selection was observed in SESO/SOAE post-treated animals compared to control animals (Fig. 3C), suggesting the possibility that reduced expression of other chemotactic factors may have contributed to reduce further accumulation and presence of macrophages and other inflammatory cells.
In summary, SESO/SOAE diet reduced lesions and inflammation in LDL-R −/− mice with preexisting atherosclerosis. More importantly, this reduction in lesion burden appears to not require LDL receptor and is independent of changes in plasma cholesterol.
Footnotes
Acknowledgments
This study was supported by the National Institutes of Health grant 5R01AT004106-05. The authors thank Mitsushita Doomra and Dr. Deepshika Bhardwaj for their assistance during animal feeding.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.