
Abstract
P2Y2 and P2Y4 receptors are physiologically activated by uridine 5′-triphosphate (UTP) and are widely expressed in many cell types in humans. P2Y2 plays an important role in inflammation and proliferation of tumor cells, which could be attenuated with the use of antagonists. However, little is known about the physiological functions related to P2Y4, due to the lack of selective ligands for these receptors. This can be solved through the search for novel compounds with antagonistic activity. The aim of this study was to discover new potential antagonist candidates for P2Y2 and P2Y4 receptors from natural products. We applied a calcium measurement methodology to identify new antagonist candidates for these receptors. First, we established optimal conditions for the calcium assay using J774.G8, a murine macrophage cell line, which expresses functional P2Y2 and P2Y4 receptors and then, we performed the screening of plant extracts at a cutoff concentration of 50 μg/mL. ATP and ionomycin, known intracellular calcium inductors, were used to stimulate cells. The calculated EC50 were 11 μM and 103 nM, respectively. These cells also responded to the UTP stimulation with an EC50 of 1.021 μM. Screening assays were performed and a total of 100 extracts from Brazilian plants were tested. Joannesia princeps Vell. (stem) and Peixotoa A. Juss (flower and leaf) extracts stood out due to their ability to inhibit UTP-induced responses without causing cytotoxicity, and presented an IC50 of 32.32, 14.99, and 12.98 μg/mL, respectively. Collectively, our results point to the discovery of potential antagonist candidates from Brazilian flora for UTP-activated receptors.
Introduction
P2
P2Y2 and P2Y4 receptors are physiologically activated by uridine 5′-triphosphate (UTP) and promote an increase in intracellular calcium through PLC/IP3. 2 They are expressed in many cell types, such as epithelial cells, monocytes, macrophages, neutrophils, cardiomyocytes, and organs, including the brain, heart, kidneys, liver, spleen, and muscle. 3 They have effects on chloride secretion in epithelial cells from the airway and eyes, and also induce water secretion in bowel epithelial cells. These functions are important to promote surface lubrification and mucus hydration, which can improve disease treatment, such as dry eye disease and chronic constipation. 2,4
P2Y2R also plays a role in neuroprotection, chemokinesis, and in the proliferation of hepatocytes and tumor cells. 5 –8 P2Y2 is also involved in the production of inflammatory modulators, such as cyclooxygenase-2 (COX-2) and prostaglandin E2 (PGE2), and neutrophil accumulation in lungs and liver during inflammation. 9 –13 Thus, P2Y2 antagonists might alleviate inflammation signals.
P2Y4R participates in Na+, K+, and Cl− regulation processes, but its functions are not completely characterized, as it is expressed in the same tissues and cell types as P2Y2R, and activated by the same physiological ligand, UTP. 14 –17 Therefore, the lack of selective ligands impairs the discovery of new functions associated with these receptors. However, this problem can be solved by the search for novel compounds with antagonistic activity on P2Y2 and P2Y4 receptors.
Natural products are an important source for the discovery of new molecules with antagonistic activity. They offer the advantage of millenary use in the treatment of several diseases, such as inflammation, parasitoses, pain, and recently, cancer, constituting the basis of traditional oriental medicines. 18 –20 Among the 13 drugs approved for global marketing between 2005 and 2007, five were classified as natural products, whereas the others were semisynthetic or derived, which reinforces natural product importance for clinical use. 21 In addition, the literature reports that some biological species express compounds with antagonistic activity on P2 receptors, such as emodin from Rheum officinale Bail and amentoflavone from Rheedia longifolia Planch & Triana on P2X7R. 22,23 However, due to the large number of samples for testing, implementation of high-throughput screening (HTS) methodologies is necessary to meet this demand, since they have the advantage of evaluating many compounds in a short period of time. 24,25
In this scenario, intracellular calcium measurement techniques emerge as a tool for the screening of new antagonists, since P2Y2 and P2Y4 receptors are coupled to the calcium signaling pathway. 26 Kaulich et al. tested a series of 40 flavonoids using calcium assays to discover novel lead antagonists for P2Y2R. They found that certain flavonoids could inhibit P2Y2R and suggest using their structures to develop new compounds with antagonistic activity. 27 Recently, Ito et al. applied an intracellular calcium measurement protocol through a HTS campaign to search for novel antagonists for the P2Y6 receptor and found one compound, TIM-38, with potential activity. 28
Thus, the aim of this study was to improve a protocol based on intracellular calcium measurements and apply it in the discovery of natural product compounds with antagonist activity on P2Y2 and P2Y4 receptors.
Materials and Methods
Reagents
Chemicals were purchased from Sigma Chemical Co. (St. Louis, MO, USA), including ATP, UTP, ADP, UDP, αβ-meATP, Suramin, DMEM, and RPMI culture mediums, NaCl, KCl, Na2HPO4, KH2PO4, MgCl2, CaCl2, Glucose, DMSO, MTT ((3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) tetrazolium, Probenecid, and Ionomycin. Fetal bovine serum was obtained from Cultilab (Campinas, SP, Brazil) and Fluo-4 AM was obtained from Life Technologies (CA, USA).
Extracts
Extracts used in this study were collected from Brazilian biomes (Amazon, Cerrado, Caatinga, Atlantic Forest, Pantanal, and Pampas), obtained through methanol extraction, and deposited at Bioprospecting Platform from the Chemistry of Natural Products Laboratory.
The stems from Joannesia princeps Vell. (blind code: JA2) were collected in the Atlantic biome, in Linhares, in the state of Espírito Santo, Brazil. The stems were crushed and mixed with a 1:1 mixture of CH2Cl2-CH3OH to obtain the extract deposited at voucher number EX294. Peixotoa A. Juss were collected in the Cerrado biome, in Tiradentes, located in the state of Minas Gerais, Brazil. Peixotoa A. Juss flower (blind code: RA3) and leaf (blind code: RB3) extracts are deposited at voucher numbers EX9355 and EX9356, respectively. A botanical specimen was deposited at the Federal University of São João del Rei Herbarium under code HUFSJ1103. The flowers and leaves were crushed and homogenized with ethanol to obtain the crude extracts. For the experiments, they were resuspended in a 1 mL Dulbecco's PBS solution containing 0.5% DMSO. The extracts were tested using a single-blind method, that is, the professional responsible for performing screening experiments did not know the name of the biological species that originated the samples. 29 Because of this, they were identified by a code.
Cell culture
J774.G8 is a murine macrophage cell line that expresses P2 receptors. The J774.G8 cell line was kindly provided by Dr. Vinícius Cotta from the Laboratory on Thymus Research at the Oswaldo Cruz Institute, Rio de Janeiro, Brazil. The cells were routinely maintained in culture with Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, at 37°C in a 5% CO2 atmosphere. The medium was changed every 3 days and the cells were adjusted to a concentration of 2 × 106 cells per 150-cm2 cell culture flask (Corning, NY, USA). Cell viability was analyzed by the Trypan Blue assay. For this, 10 μL of cell sample was diluted 1:100 in 0.04% Trypan Blue solution. After dilution, 10 μL of the sample were added into the Neubauer chamber and observed under optical microscope Nikon Eclipse TE 2000-S (Nikon, Tokyo, Japan) to count viable cells. Calcium assays were conducted only when the viability range was above 90%.
Calcium assays
J774.G8 cells were plated at a concentration of 2 × 105 cells/well in a 96-well black-wall, clear-bottom plate (Corning, NY, USA), and maintained in culture with DMEM supplemented with 10% fetal bovine serum, at 37°C in a 5% CO2 atmosphere for 24 h. Subsequently, the medium was replaced by 100 μL of Dulbecco's PBS (DPBS) (136 mM NaCl, 2.68 mM KCl, 8 mM Na2HPO4, 1.46 mM KH2PO4, 0.5 mM MgCl2, 0.9 mM CaCl2, 5.55 mM Glucose, and 2.5 mM Probenecid–pH 7.4) containing Fluo-4 AM [2 μM] and the cells were incubated for 60 min at 37°C in a 5% CO2 atmosphere. Extracellular Fluo-4AM was then removed by three consecutive washes with 200 μL DPBS and the cells received a final volume of 150 μL DPBS. Subsequently, cells were treated with extracts [50 μg/mL] and PPADS [300 μM], Suramin [300 μM], and Reactive Blue-2 (RB-2) [20 μM] antagonists for 30 min. Finally, the assay plate was placed into the FlexStation III equipment (Molecular Devices, CA, USA), which performed the agonist addition to each well, diluted in 50 μL DPBS. The mobilization of intracellular calcium was measured in real-time prior, during, and after the addition of the agonists at λ (excitation) of 485 nm and λ (emission) of 525 nm. Total measurement runtime was 90 sec and agonists were added at the 20-sec mark. Readings were carried out every 1.52 sec and a total of 60 readings were obtained for each well. The ΔFU, that is, the difference between the fluorescence peak associated with the maximum concentration of intracellular calcium ([Ca2+]i) and basal fluorescence of calcium was determined according to the protocol of Hansen and Bräuner-Osborne (2009). 26 The wells whose signal did not return to approximately 30% of the baseline after stimulation were discarded as quality control.
Cell loading observation under a fluorescence microscope
J774.G8 cells were loaded with Fluo-4AM as described previously and observed under a Nikon Eclipse TE 2000-S fluorescence microscope (Nikon, Tokyo, Japan), using 20 × objectives. Fluo-4 was excited using a mercury lamp and a set of filters: exciter: 480/30 and barrier: 535/40. Ionomycin was added after 40 sec of observation and pictures were captured with an Infinity 3 Microscope Digital Camera (Lumenera, Ontario, Canada).
RNA extraction, Reverse transcriptase polymerase chain reaction, and electrophoresis
mRNA was isolated from the J774.G8 cell line from a concentration of 5 × 106 cells using TRIzol (Life Technologies, CA, USA), according to the manufacturer's specifications. Then, 20 μL of mRNA was reverse transcribed at 42°C for 1 h and at 70°C for 5 min using 1 μL of reverse transcriptase enzyme. Next, 4 μL of cDNA samples were used with 4 μL of a primer sequence specific for each P2 receptor subtype, as shown in Table 1. These samples were used for the RT-PCR analysis in a 50 μL reaction volume containing 5 μL of Buffer 10 × , 4 μL Mg2+, 0.4 μL dNTP, 0.2 μL Taq polymerase, and 4 μL of water. Cycling conditions were: 94°C for 45 sec, followed by 35 cycles at 54°C (P2Y11), 55°C (P2Y6), 57°C (P2X1), 58°C (P2X3, P2Y14, and P2Y13), 60°C (P2Y2, P2X2, P2X5, P2X6, P2Y12, P2Y1, P2Y4, and GAPDH) or 65°C (P2X4 and P2X7) for 1 min and 72°C for 45 sec. PCR products were submitted to electrophoresis on a 2% agarose gel and visualized by ethidium bromide staining under a transilluminator.
Primer Sequence of P2 Receptors Used in RNA Extraction, Reverse Transcriptase Polymerase Chain Reaction, and Electrophoresis Experiments
Animals
The experiments with animals were approved by Oswaldo Cruz Institute Ethics Committee on the use of Animals, under identification code L-037/2017, approved on November 24, 2017, and followed the ethical principles in animal experimentation from the Brazilian College of Animal Experimentation. Healthy male C57BL/6 mice weighing between 25 and 30 g were obtained from the Oswaldo Cruz Central Bioterium. The animals were housed under 12-h light/12-h dark cycle conditions with free access to food and water.
Peritoneal macrophage isolation
Animals were euthanized by CO2 asphyxiation. Subsequently, 5 mL of RPMI medium was injected into the peritoneal cavity. Peritoneal cells were collected and macrophages were isolated by centrifugation at 378 g for 10 min. Cell viability was analyzed by the Trypan Blue assay. Peritoneal macrophages were plated at a concentration of 4 × 105 cells/well in a 96-well black-wall, clear-bottom plate and maintained in RPMI culture supplemented with 10% fetal bovine serum, at 37°C in a 5% CO2 atmosphere for 24 h. Calcium assays were carried out using the same conditions described previously.
Cell viability measurement
J774.G8 cells were plated at a concentration of 2 × 105 cells/well in a 96-well plate (Corning, NY, USA), and maintained in culture with DMEM supplemented with 10% fetal bovine serum, at 37°C in a 5% CO2 atmosphere for 24 h. Next, cells were treated with extracts [50 μg/mL] and the antagonists, PPADS [300 μM] and RB-2 [20 μM] for 1, 6, or 24 h. Cells treated with Triton-X (0.1%) were considered negative viability controls, whereas positive controls were obtained with cells that did not receive any treatment. Subsequently, the medium was replaced by 180 μL of DMEM without phenol red and each well received 20 μL of MTT solution [100 μg/well]. The plate was then incubated for 3 h at 37°C in a 5% CO2 atmosphere and then centrifuged at 312 g for 1 min. Finally, supernatants were collected and formazan crystals were dissolved in 100 μL DMSO. 30 The absorbance of the wells was measured on a FlexStation III equipment, using λ: 570 nm.
Data analyses
Each sample was measured in triplicate and all experiments were performed on at least 3 independent days. All data are presented as mean ± SDM, whereas the curves are presented as mean ± SEM. To test if samples follow a Gaussian distribution, the D' Agostino and Pearson normality test was used. If data followed a Gaussian distribution, an appropriate parametric test was applied, if not, an appropriate nonparametric test was used. The applied tests were specified in figure legends. P values of .05 or less were considered significant. Graphs and statistical analyses were performed using the GraphPad Prism version 7 software (GraphPad Software, San Diego, CA, USA). The quality of the calcium assay was assessed by calculating the z′-factor. The z′-factor is a valuable tool to evaluate the robustness and suitability of HTS assays. z′-factor values above 0.5 consider the assay as excellent and equal to 1, an ideal assay, whereas values below 0.5 consider that the assay should be reformulated.
31
This parameter was calculated using the following equation:
where cells stimulated with UTP at [10 μM] were considered positive control (concentration used in screening experiments) and cells stimulated with DPBS only were considered negative control.
Results
Establishment of a protocol for the detection of intracellular calcium mobilization
Our primary aim was to discover new antagonists for P2Y receptors activated by UTP, so a protocol for the detection of intracellular calcium mobilization was first established. Fluo-4 is a calcium indicator widely applied in HTS assays of metabotropic receptors that induce increase in intracellular calcium. 32 The literature states a range of Fluo-4 used in calcium assays, between 2 and 4 μM. 33 –36 To use low doses of the dye, the first step was to optimize Fluo-4 concentration in our assay conditions, making it more economical for HTS. In this context, we investigated concentrations ranging from 1 to 8 μM, using ATP [100 μM] to stimulate the cells due to its activity as a main physiological P2 receptor agonist, widely expressed in J774.G8 cells. As shown in Figure 1A, no significant difference in the quantification of calcium responses for concentrations between 1 to 4 μM was observed. Concentrations of 6 and 8 μM have shown significant difference only compared with 1 μM.
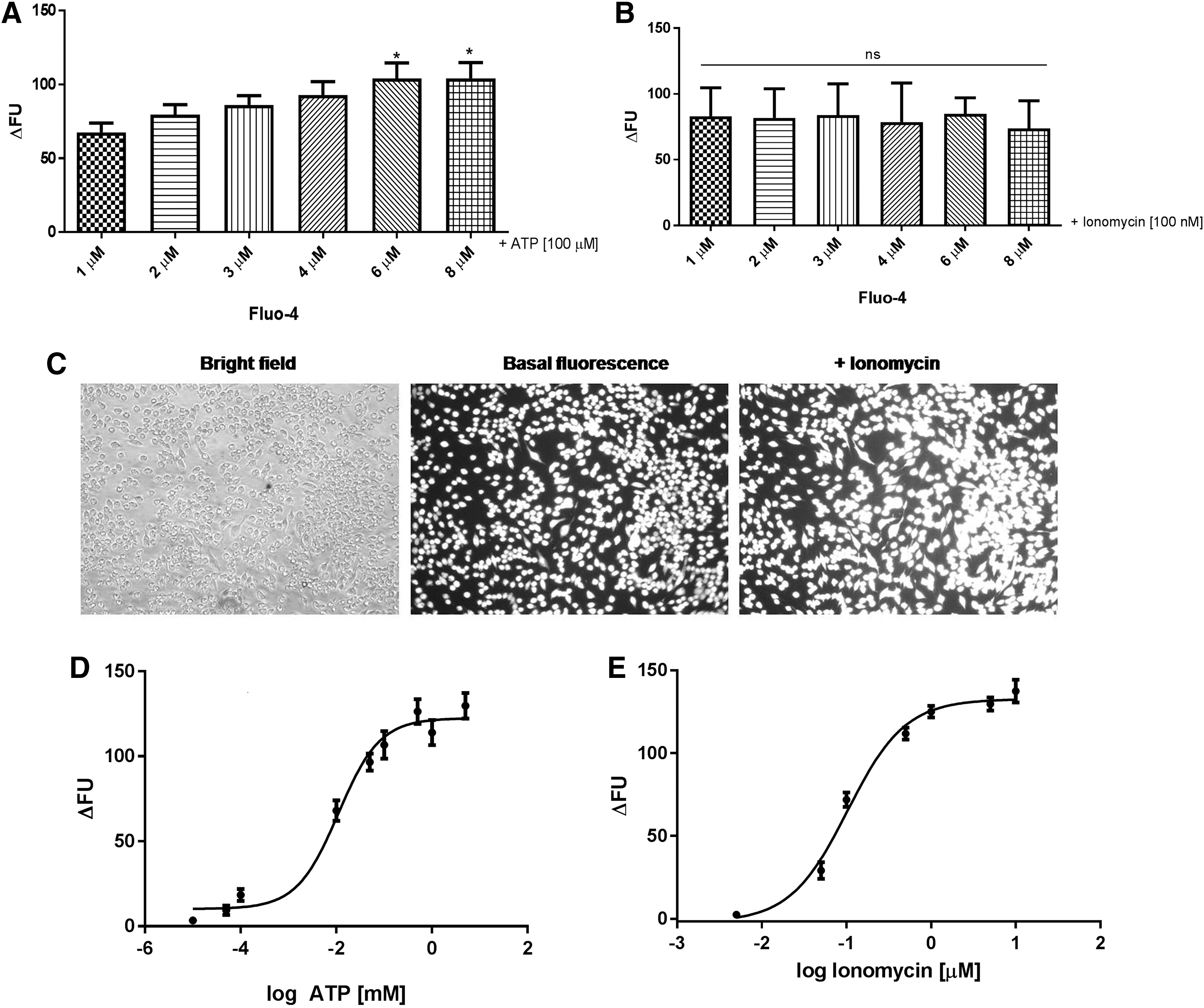
Measurement of [Ca2+]i after ATP and ionomycin stimulation in cells stained with Fluo-4. J774.G8 cells stained with different Fluo-4 concentrations were stimulated with [100 μM] ATP
A similar result was obtained in cell stimulation with ionomycin, a calcium ionophore, which promotes a massive transport of this ion from the extracellular to intracellular medium. All concentrations evaluated have a similar profile for calcium response quantification (Fig. 1B). Thus, the selected Fluo-4 concentration to use in our experiments was 2 μM, as calcium responses in this concentration did not differ from higher concentrations, and is cited in the literature.
As we could observe in Figure 1C, cells loaded with Fluo-4 at 2 μM shown a homogenous dye scattering in their cytosol and, after stimulus with ionomycin, become more fluorescent.
Next, two assays were performed to verify Fluo-4 sensibility at the established concentration (2 μM). Cells were stimulated with increasing ATP and ionomycin concentrations, and Fluo-4 fluorescence increased in a concentration-dependent manner, as shown in Figure 1D and E. EC50 for ATP and ionomycin were 11 μM and 103 nM, respectively.
Therefore, in this primary step, Fluo-4 concentrations at 2 μM were ideal for our assay, since cells presented homogenous dye loading and sensibility in responding to calcium stimuli in increasing concentrations.
Quality assessment of calcium responses induced by P2 receptors
To characterize the quality of calcium responses induced by P2 receptors through ATP stimulation acquired on the FlexStation III equipment, we analyzed original records. Original records indicated that DPBS addition alone did not cause signal variations, as expected (Fig. 2A). ATP stimulus, on the other hand, increased intracellular calcium levels, as seen by the fluorescence peak shown in Figure 2B. This effect was reversed with the addition of EGTA, a calcium chelating agent, indicating that the fluorescence is associated with calcium, as shown in Figure 2C. Cells stimulated with ionomycin also increased their fluorescence (Fig. 2D). No decline in intracellular levels during the selected time frame for this experiment, that is, 90 sec, was observed. Therefore, these experiments validated our calcium measurement protocol using the FlexStation III equipment.
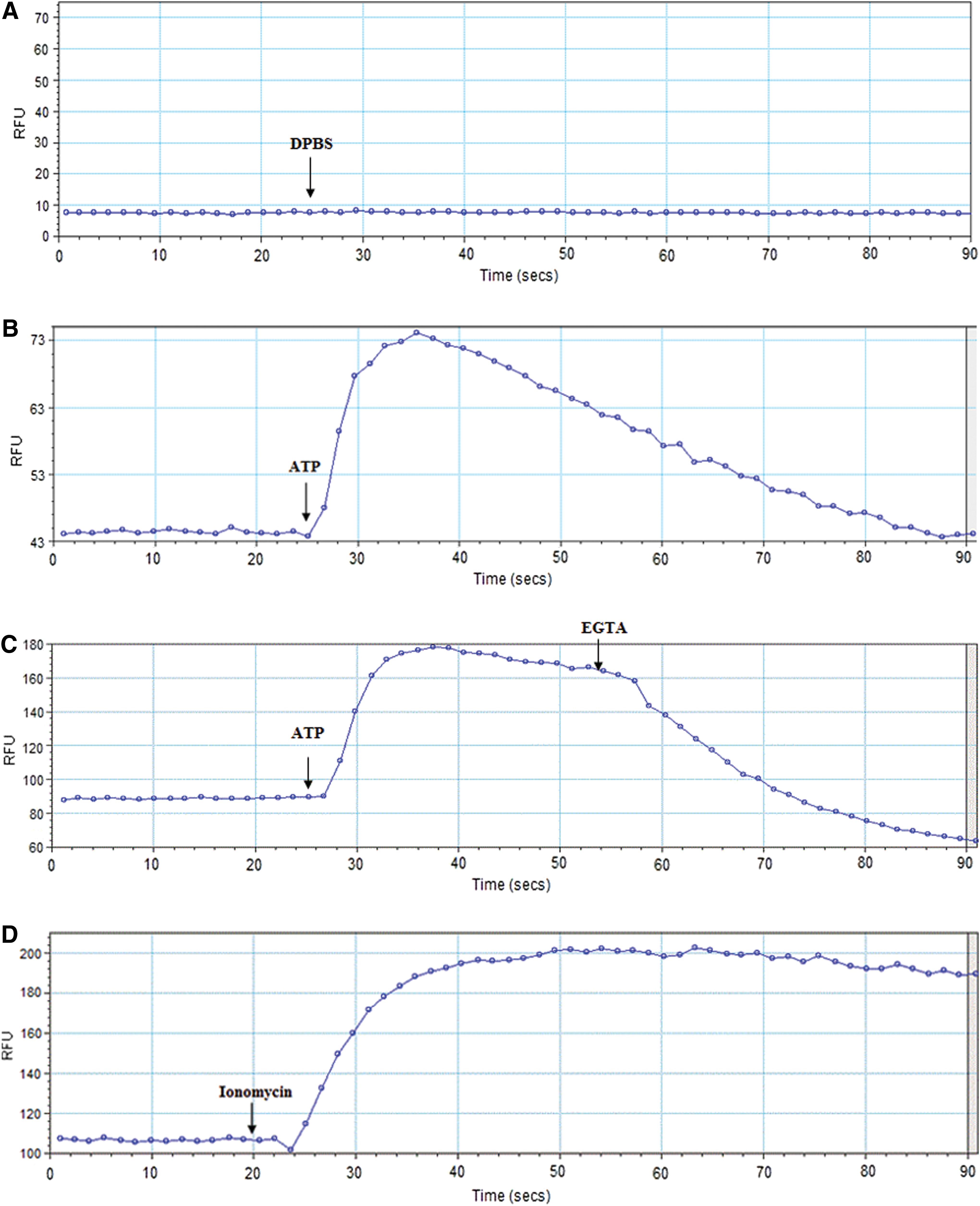
Original records of calcium signals obtained on a FlexStation III. Original records of calcium signals obtained on a FlexStation III equipment after addition of 50 μL of DPBS
Characterization of P2 receptors in J774.G8 cells
Before beginning the screening experiments, we attempted to identify which P2 receptors (P2R) were expressed in this cell line to guide our assay, avoiding bias, since some P2R subtypes were activated by the same agonists. First, the RT-PCR technique was performed, aiming to verify which P2R are expressed in J774.G8 cells. As observed in Figure 3A and B, cDNA fragments corresponded to P2X1 (775 bp), P2X7 (186 bp), P2Y1 (683 bp), P2Y2 (850 bp), P2Y4 (544 bp), P2Y6 (480 bp), P2Y11 (274 bp), and P2Y12 (360 bp).
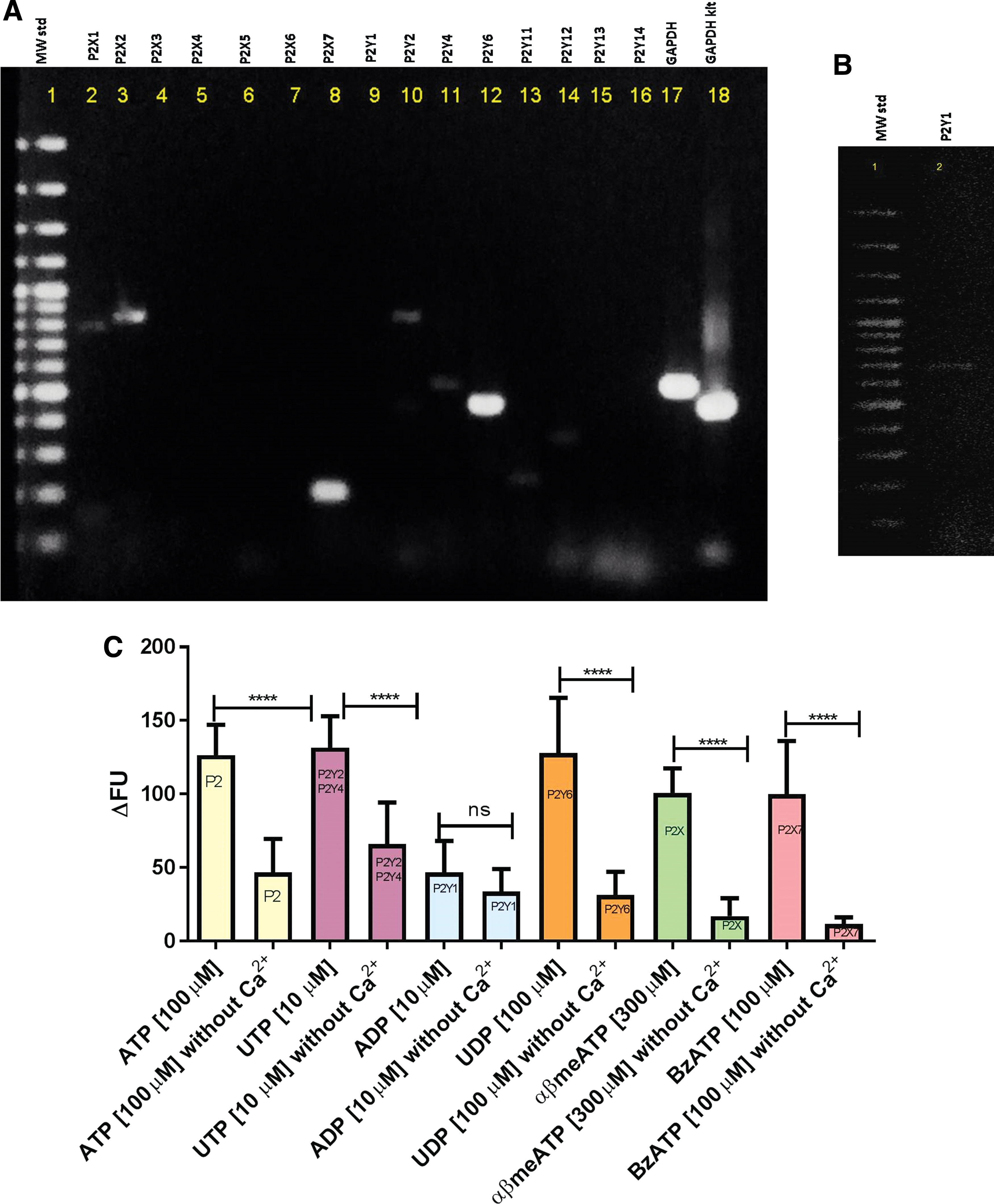
P2 receptors subtypes expressed in J774.G8 cell line. In
All these receptors, except P2Y12R, are associated with intracellular calcium increase signaling pathways through opening of a nonselective cation ion channel (P2X) or PLCβ/IP3 system activation (P2Y). Thus, we further investigated the response of P2R subtypes expressed in J774.G8 cells using selective agonists. As shown in Figure 3C, this cell line expresses P2X and P2Y receptors, such as P2Y2 and P2Y4, activated by UTP, P2Y1, and P2Y6, which were activated by ADP and UDP, respectively. Cells also express P2X7 that promoted calcium response after stimulation with BzATP (P2X7 selective agonist). ATP and αβmeATP, nonselective agonists, were used for observing general P2 and P2X receptor calcium responses, respectively. These results confirm the expression of P2 receptors observed in RT-PCR, including UTP-activated P2Y receptors, that is, P2Y2 and P2Y4, which are our target subtypes. Interestingly, we observed that all agonists, with the exception of ADP, presented lower responses without extracellular calcium, possibly indicating the participation of store-operated calcium channels. In the case of ATP and αβmeATP, this response could be regarding ionotropic channels.
Characterization of UTP-induced calcium responses
Since our cells express UTP-activated receptors, that is, P2Y2 and P2Y4, they were stimulated with increasing concentrations of this agonist, to observe UTP response profile (Fig. 4A). The determined EC50 was of 1.021 μM. Next, we obtained the P2Y2 and P2Y4 antagonist profiles by performing an antagonism assay using Reactive-Blue 2 (RB2), Suramin, and PPADS. 2 As shown in Figure 4B, RB2, Suramin, and PPADS partly inhibited UTP-induced intracellular calcium mobilization. To observe the antagonistic profile of these inhibitors of P2Y2 and P2Y4 receptors, we pretreated cells with different Suramin, RB2, and PPADS concentrations and obtained IC50 of 173.3, 22.98, and 74.24 μM, respectively (Fig. 4C–E). We also observed that Suramin and RB2 partially block intracellular UTP-induced calcium mobilization.
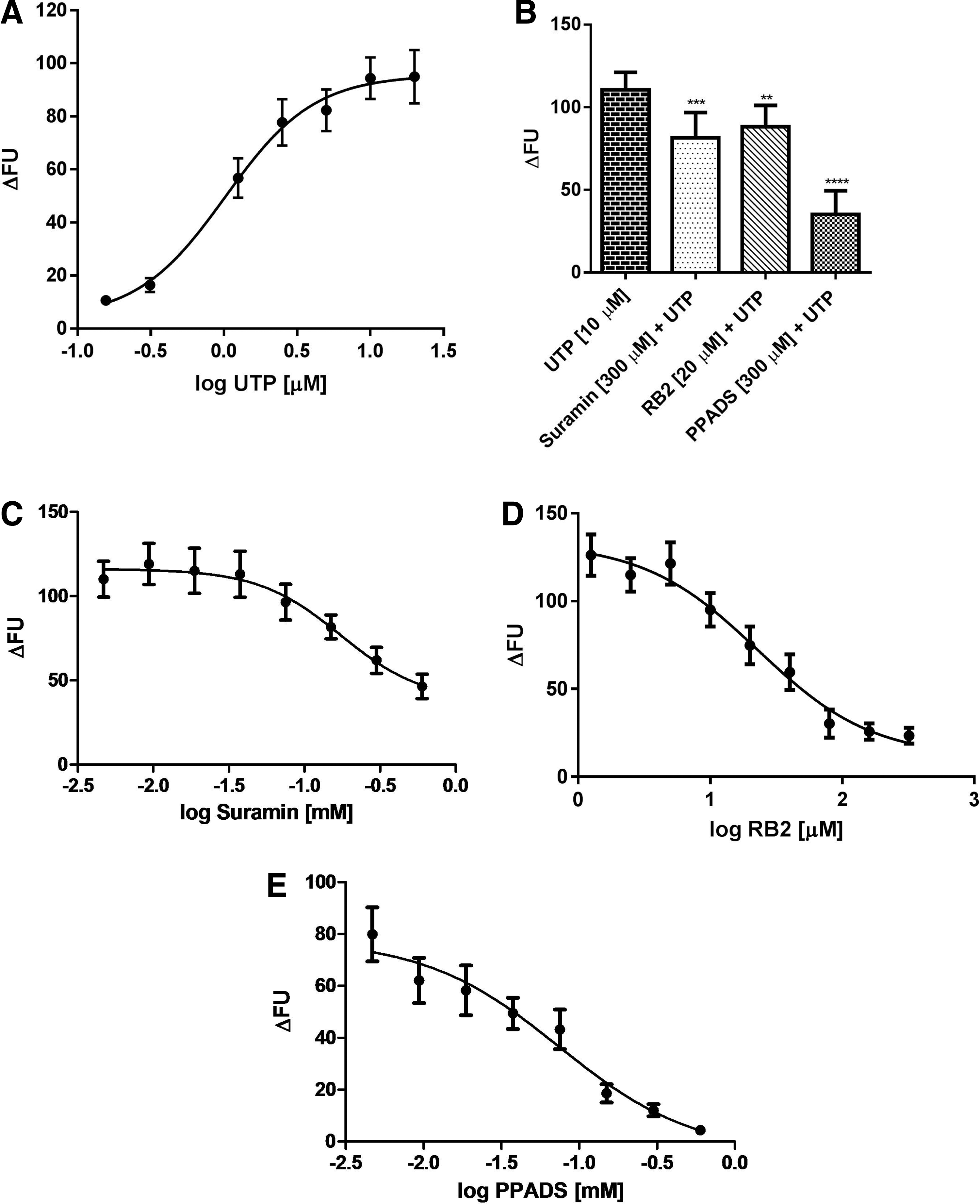
Calcium mobilization induced by UTP can be partially inhibited by P2Y2 and P2Y4 nonselective antagonists. UTP concentration–response curve was obtained from J774.G8 cells stained with 2 μM of Fluo-4. Data are presented as Mean ± SEM of three independent experiments performed in triplicate
After characterization of UTP calcium responses and P2Y2 and P2Y4 antagonist inhibition profile, we calculated the z′-factor of our assay. The z′-factor is a statistical parameter used to measure assay robustness. After applying the mathematical z′-factor equation, we obtained a value of 0.645. Therefore, the z′-factor of our assay, which is above 0.5, is within the optimal range (z′ ≥ 0.5). Taken together, these evaluated characteristics suggest that our protocol was appropriate for use.
Mini HTS application
Finally, we performed the screening of 100 natural product extracts, summarized in Table 2. Nine extracts partially inhibited calcium mobilization induced by UTP, namely: JA2 (Joannesia princeps Vell. - stem), JA5, JB2, JB7, P2B6, P5C1, P5F3, RA3 (Peixotoa A. Juss - flower), and RB3 (Peixotoa A. Juss - leaf). The same experiment was repeated using peritoneal macrophages from mice, a primary cell that expresses P2R (Fig. 5A). When using these cells, we observed that only J. princeps Vell. extract was able to significantly inhibit UTP-calcium responses. The mean of J. princeps Vell in peritoneal macrophages was less than in J774.G8 cells (Table 2) (21.07 ± 18.24 compared with 52.97 ± 29.48), which could reflect differences between cell lines and primary cells. Peixotoa A. Juss extracts (RA3 and RB3) were not been tested in peritoneal macrophages due to sample scarcity. Subsequently, cells were treated with increasing concentrations of J. princeps Vell. and Peixotoa A. Juss (flower and leaf) extracts, and the obtained IC50 were equal to 32.32, 14.99, and 12.98 μg/mL, respectively (Fig. 5B–D). Aiming to verify if calcium response inhibition was the result of possible extract cytotoxicity, viability tests were performed using the MTT technique. As observed in Figure 5E, only leaf extract caused a slight decrease in cell viability, but although nonsignificant. Taken together, we characterized J. princeps Vell. and Peixotoa A. Juss extracts as potential antagonist candidates for UTP-stimulated receptors.
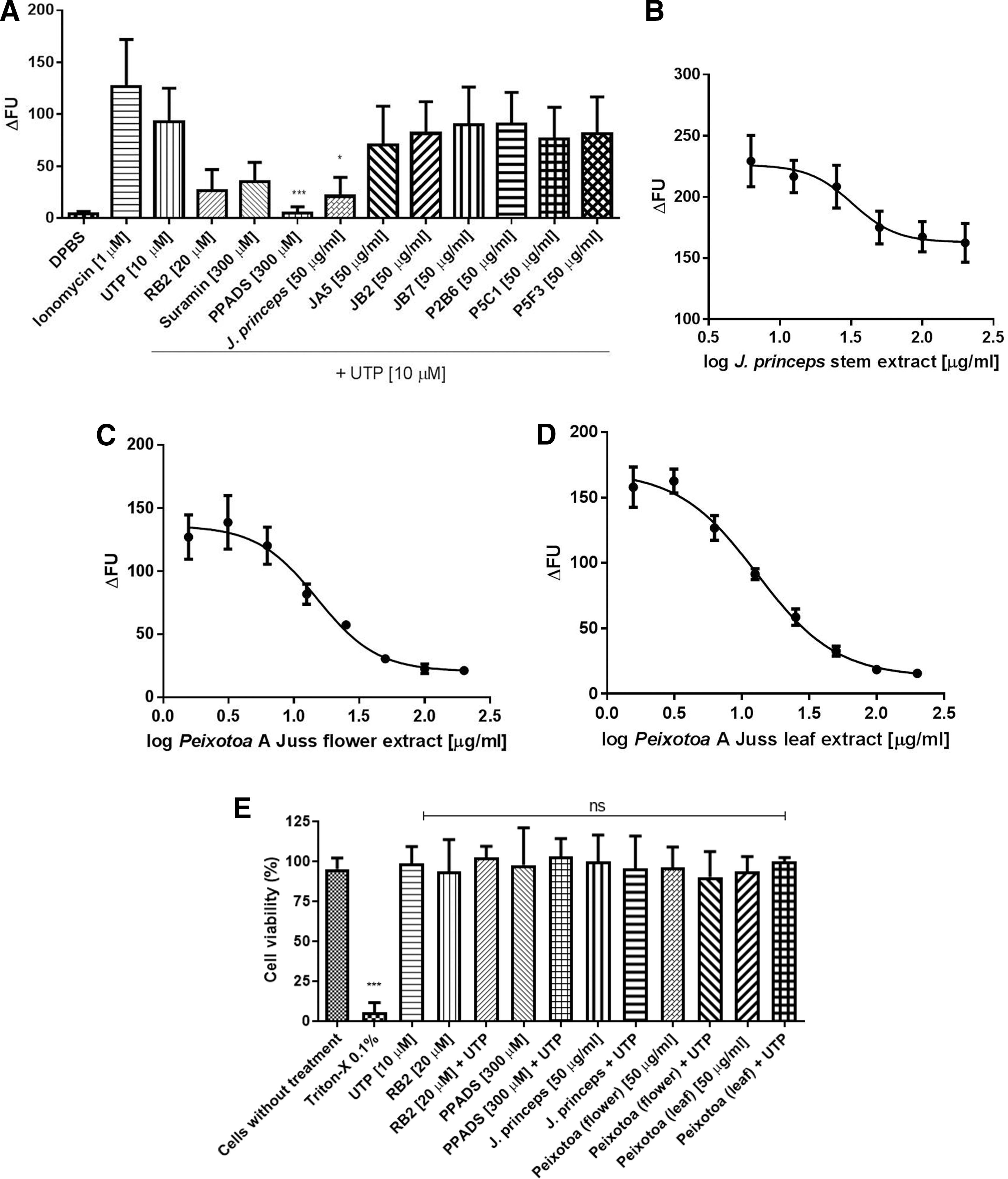
J. princeps and Peixotoa A. Juss extracts partially inhibited calcium responses induced by UTP. Peritoneal macrophages stained with 2 μM of Fluo-4 were pretreated with RB2 [20 μM], Suramin [300 μM], PPADS [300 μM], and extracts [50 μg/mL] during 30 min. Then, cells were stimulated with UTP [10 μM] and monitored for 90 sec. Data are presented as Mean ± SDM of three independent experiments performed in triplicate. Data analyses were made using one-way ANOVA and Holm-Sidak post-test; P < .05. (*) indicates significant differences in relation to UTP
Mini High-Throughput Screening Application with Extracts from Brazilian Plants: Inhibition Effect on Calcium Mobilization Induced by UTP Through P2Y2 and P2Y4 Receptors
J774.G8 cells labeled with 2 μM Fluo-4 were pretreated with extracts [50 μg/mL] for 30 min. Then, cells were stimulated with UTP [10 μM] and were monitored for 90 sec. Mean ± SDM of three independent experiments. Data analyses were made using one-way ANOVA and Tukey post-test; P < .05.
Asterisks Indicates significant differences in relation to UTP.
Joannesia princeps Vell. stem extract.
Peixotoa A. Juss. flower extract.
Peixotoa A. Juss. leaf extract.
Discussion
P2Y2 and P2Y4 are purinergic receptors physiologically activated by UTP, which promote an increase of intracellular calcium through PLCβ/IP3 when activated. 2 They are expressed in many cell types, such as epithelial cells and leukocytes, and even in organs, such as the brain, heart, kidneys, liver, spleen, and muscle, 3 where they play important roles. Likewise, these P2Y receptors are associated with certain diseases, such as cystic fibrosis, dry eye disease, Alzheimer, and cancer. 2,4,5 Thus, pharmaceutical companies have been developing drugs that act on these P2Y receptors, especially P2Y2, to treat these related diseases. 37 –39 However, the lack of selective ligands that act on P2Y2 and P2Y4 receptors compromises the discovery of new functions associated with these receptors, which can be solved by the search for new antagonist compounds. 17 Therefore, the aim of this study was to improve a methodology for the detection of intracellular calcium mobilization to discover new molecules with antagonistic activity on UTP-activated P2Y receptors. For this reason, we established a protocol to evaluate intracellular calcium mobilization.
First, the calcium indicator concentration was optimized. Fluo-4 is widely applied in HTS assays of metabotropic receptors that induce an increase of intracellular calcium. 32 We chose 2 μM as the final concentration, due to the apparent homogenous cellular loading in cells and the fact that no significant differences were observed compared with other concentrations. In addition, it is important to use the lowest concentration that results in an adequate signal, thereby reducing reagent consumption and expenses during screening. 31 Gee et al. also demonstrated that the use of the lowest concentrations of calcium indicators can reduce the buffer effect on calcium, as well as minimize the levels of toxic products, such as formaldehyde and acetic acid, which are produced through the hydrolysis of acetoxymethyl esters. 40 Furthermore, using this concentration, we demonstrated that cells were able to react to different stimuli, which increase intracellular calcium levels in a concentration-dependent manner, such as P2R agonists, including ATP and UTP. The EC50 found for ATP (11 μM) is slightly higher than that reported by Li et al. when they performed calcium experiments using the FlexStation III (1.1 μM and 58 nM for HEK293 and CHO-K1 cells, respectively). 32 This could be a sum of responses from various ATP-activated P2R, such as P2Y2, P2Y11, P2X1, P2X2, and P2X7 subtypes. The latter could have displaced the EC50, since its activation occurs at concentrations higher than 100 μM. Meanwhile, the EC50 found for ionomycin (103 nM) was lower than that reported by Valentin et al. (2011), that is, 1.5 μM when they performed a calcium assay using a microplate reader with human endothelial cells from the umbilical vein (HUVEC). 41
We also observed some expected calcium signal characteristics, such as the uniformity of the basal signal, a signal peak after the ATP stimulus, indicating an increasing of [Ca2+]i and its return to basal levels after a response time, which, in calcium events, usually occurs in seconds. In addition, a decrease of calcium responses in the presence of EGTA, a calcium chelating agent, was also observed. A similar response profile was obtained through calcium imaging (data not shown). Ionomycin stimulation also induced an increase in intracellular calcium levels, although with no return to basal levels during the experiment period (90 sec). However, a longer measurement time concerning the ionomycin response could be required to observe this return. However, increasing the well-measurement time could lead to a decrease in the number of samples to assess in an established period of time. Therefore, we opted to maintain the set time at 90 sec.
Moreover, we calculated the z′-factor, a measure for assay robustness, obtaining a value of 0.645, indicating that our protocol was adequate for use. Ito et al. performed an HTS campaign to identify novel P2Y6R antagonists using P2Y6-1321N1 cells, and obtained a z′-factor of 0.80, 28 whereas Valentin et al. (2011) used primary cells (HUVEC) to perform a HTS based on calcium measurement and obtained a z′-factor of 0.60. 41 These findings suggest that the use of transfected cells could improve the performance of the assay. However, the use of transfected cells can add costs to the screening. Thus, we suggest that it be applied in a second research moment, to verify hit responses.
J774.G8 murine macrophage cell line expresses P2Y activated by UTP, targets of the screening assay proposed herein. Using the RT-PCR technique, we detected cDNA for P2X1, P2X2, P2X7, P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, and P2Y12 subtypes. These P2R subtypes have been described in macrophages in many studies applying molecular and pharmacological methods. 42 –45 Furthermore, all are associated with intracellular calcium increase signaling pathways through the opening of a nonselective cation ion channel (P2X) or PLCβ/IP3 (P2Y), with the exception of P2Y12. Therefore, we characterized these subtypes using the following agonists: ATP, a nonselective P2 receptor agonist; UTP, a P2Y2 and P2Y4 receptor agonist; ADP for the P2Y1 receptor; UDP for the P2Y6 receptor; BzATP, a selective P2X7 agonist; and αβmeATP for P2X receptors. These data showed the functional expression of these receptors. Calcium responses decreased in the absence of extracellular calcium, suggesting the participation of store-operated calcium channels and ionotropic receptors in the amplification of intracellular calcium mobilization. 46
We also obtained the profiles of P2Y2 and P2Y4 agonist and antagonists. J774.G8 cells were stimulated with increasing concentrations of UTP and the EC50 obtained was 1.021 μM. It was similar to that cited by King et al.: 1.1 μM for P2Y2 and 0.20 μM for P2Y4 receptors. 47 In relation to the antagonists, we observed that Suramin and RB2 partially block intracellular UTP-induced calcium mobilization, whereas PPADS was more effective. King et al. cited that PPADS (100 μM) and RB2 (100 μM) inhibited approximately 30% of P2Y4R, whereas Suramin is ineffective. 47,48 von Kügelgen cited that P2Y2 is blocked by Suramin and RB2, but not by PPADS. 48 We also hypothesize that the participation of store-operated calcium channels could have influenced their inhibition at some level.
It is worth mentioning that our study is one of the pioneers in the search for novel antagonists using natural products, which present significant diversity concerning number of species and chemical structures, both acquired over millions of years of evolution. Thus, we tested 100 extracts from a natural library at a cutoff concentration established at 50 μg/mL. First, nine extracts demonstrated partial inhibition of UTP-induced calcium responses in J774.G8 cells. After a repetition of this experiment with a primary cell, that is, peritoneal macrophages, which express P2Y2 and P2Y4 receptors, 44 we found that only J. princeps Vell. extract inhibited this activity. Two other extracts (Peixotoa A. Juss leaf and flower) were not tested in these cells due to sample scarcity. However, aiming at observing the occurrence of this inhibition at higher doses, we treated J774.G8 cells with increasing concentrations of the assessed extracts, and observed that calcium responses were inhibited in a concentration-dependent manner, suggesting potential ability to block UTP-activated P2Y receptors. We then asked if inhibition could be due to cell toxicity. To answer this question, we performed a cell viability assay applying the MTT technique and observed no cytotoxicity.
J. princeps Vell. is a tree approximately 20 m in height belonging to the Euphorbiaceae family. In Brazil, it is found in the Atlantic Rainforest and Caatinga biomes, and can also be found in some regions of Africa and Asia. It is popularly known as “cotieira” in Brazil, with economic value in landscaping, wood extraction, and medicinal applications. 49,50 The only part of the plant used for medicinal purposes is its seed. The seed oil is used as a laxative, whereas its extract displays antihelminthic activity. 50 Recently, Donato-Trancoso et al. demonstrated that J. princeps Vell. seed oil can improve cutaneous wound closure in an experimental mice model. 51 Sousa et al. cite that the use of J. princeps Vell. in folk medicine is indicated to treat menstrual and digestive disorders, pernicious fever, microbial diseases, syphilis, scrofulous, and swelling. 50 In relation to chemical profile, Achenbach and Benirschke, in their work using J. princeps Vell. root bark, stem bark, and leaves, demonstrated that this species presents compounds belonging to the following classes: sesquiterpenes, bis-sesquiterpenes, diterpenes, triterpenes, and steroids. 52 Waibel et al. demonstrated that the methanol extract of the seeds presents secondary metabolites, such as lignans, neolignans, and sesquineolignans. 53 Recently, Camero et al. isolated 28 secondary metabolites from J. princeps Vell. leaves for the first time, including gallic acid derivatives, gallotannins, flavonoids, α-ionones, glycosylated monoterpenes, lignan, and other phenolic derivatives. 54 However, scarce reports in the literature are available concerning J. princeps Vell. stem phytochemistry, which was used in this work.
Peixotoa A. Juss is a bush belonging to the botanical family Malpighiaceae, which, in Brazil is represented by 32 genera and 300 species. The Peixotoa A. Juss genus comprises shrubs, subshrubs, and creepers, which grow on rocky and earthy substrates. It is a native plant presenting widespread distribution in Brazil. 55 Only one species belonging to this botanical genus presents folk medicinal use reported in the literature. The roots of Peixotoa cordistipula A Juss, popularly known as “joão-da-costa” are used to treat spine disorders in the west-central region of Brazil. 56 The scarcity of information about this botanical genus is also reflected in the phytochemistry field, since no studies are available in the literature concerning the characterization of its metabolites. Some information about the chemical profile of other Malpighiaceae family members is, however, available. For example, De Frias and collaborators evaluated the chemical profile of leaves of Banisteriopsis anisandra A. Juss., belonging to the Banisteriopsis genus, and found alkaloids, anthraquinones, flavonoids, and tannins. 57 Andrade et al. analyzed the chemical profile of another Malpighiaceae family member, Byrsonima crassifolia, which belongs to the Byrsonima genus, and found that barks produce the following secondary metabolites: flavonoids, anthraquinones, reducing sugars, phenolic compounds, tannins, and cardiotonic glycosides. 58 Therefore, based on the presence of metabolites in these two genera, it is possible that Peixotoa A. Juss may also synthesize some of these substances.
Despite this information, scarce studies demonstrating the potential medicinal use of other plant parts, such as leaf, stem, root, and flower, are available. In the present study, we demonstrate the potential antagonistic activity of a stem extract from J. princeps Vell. and flower and leaf extracts from Peixotoa A. Juss on P2Y2 and P2Y4 receptors, inhibiting UTP-induced calcium responses in a dose–concentration manner without causing cytotoxic effects. However, it is important to understand and characterize the active molecules from these extracts responsible for the observed inhibitory effect, which will be carried out in further studies by our group.
In conclusion, our results point to the discovery of potential antagonists for P2Y2 and P2Y4 receptors from Brazilian botanical species J. princeps Vell. and Peixotoa A. Juss. This discovery may significantly contribute to purinergic pharmacology and the application of P2 receptor antagonists in clinical therapy.
Footnotes
Acknowledgments
This work was supported by grants from Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Instituto Oswaldo Cruz (IOC). The authors thank Dr. Vinícius Cotta, who kindly donated the J774.G8 cell line, Fátima Regina Marques for technical assistance with extracts, and the Program for Technological Development in Tools for Health PDTIS-FIOCRUZ/RPT-FIOCRUZ for the use of its facilities.
This article is based on the author's dissertation and thesis, which is available in a repository in Brazil. The authors thank the Oswaldo Cruz Institute and the Postgraduate Program in Cellular and Molecular Biology for providing all the necessary conditions to complete this work.
Author Disclosure Statement
No competing financial interests exist.