
Abstract
Mojarra of Nile tilapia (Oreochromis niloticus) skeleton was used as protein source for the preparation of protein hydrolysates and peptide fractions with angiotensin-converting enzyme (ACE) inhibitory activity. The flour presented a content of 34.92% protein and a brightness (luminosity, L*) of 82.29. Protein hydrolysates were obtained from the protein-rich flour with the enzymes Flavourzyme® and Alcalase® reaching degree of hydrolysis (%DH) of 52% and 67% at 100 min of reaction, respectively. Both hydrolysates showed low-molecular-weight (MW) peptides estimated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis. The hydrolysates obtained with Flavourzyme at 60 min and at 80 min with Alcalase showed greater ACE inhibitory activity with IC50 values of 0.238 and 0.344 mg/mL, respectively. The peptide fraction A (MW >10 kDa) with Flavourzyme and fraction B (MW = 10–5 kDa) with Alcalase obtained by ultrafiltration of hydrolysates with higher DH presented IC50 of 0.728 and 0.354 mg/mL, respectively, whereas peptide fraction C (MW = 5–3 kDa) with both enzymes hydrolysates with greater ACE inhibitory activity showed IC50 values of 0.470 and 0.634 mg/mL. The components obtained in this study could be used as functional ingredients in the design and development of functional foods.
Introduction
The renin–angiotensin system is the main regulator of blood pressure in humans and it starts when renin acts on angiotensinogen, which convert into angiotensin-I (Ang I) a decapeptide inactive. 1 Angiotensin-converting enzyme (ACE) (dipeptidyl carboxypeptidase I, kinase II, E.C.3.4.15.1) is an ectoenzyme of vascular cells that is secreted in plasma and plays an important role in cardiovascular homeostasis. ACE binds to the C-terminal dipeptide of the inactive decapeptide Ang I to catalyze its conversion to the octapeptide vasoconstrictor angiotensin-II (Ang II), but also inactivates bradikinin a vasodilator nonapeptide, thereby increasing the blood pressure. 2 Protein hydrolysates have been used to obtain peptide fractions containing bioactive peptides, which are small peptide chains containing 2–15 amino acid residues. 3
Peptides have been obtained from protein hydrolysates of both animal and vegetable origin; some of these exhibit ACE inhibitory activity, which could be an alternative for the treatment of arterial hypertension. 4 ACE inhibitors of natural origin can control blood pressure and may have minimal side effects, whereas ACE inhibitors of synthetic origin can cause side effects such as cough, taste disturbance, and skin rashes, and also present risks in the health of pregnant women such as deformations in the fetus and even fetal death. 5
In the past 20 years, an emerging area is the search and identification of bioactive compounds that may be present in marine and aquatic by-products, 6 since research have reported physiological and bioactive properties of protein hydrolysates obtained from fish by-products such as the head, skin, viscera, and skeletons of different species of fish, 7 as well as the role that these play as immunomodulators, the effect on the decrease of cholesterol, its role in the control of hypertension, among others. 8 Mojarra of Nile tilapia (Oreochromis niloticus) is one of the most popular species cultured in freshwater aquaculture. However, their high demand has increased dramatically in global market, so more tilapia-processing industries have been increasing generating more by-products and causing environmental pollution; however, these by-products have a significant amount of protein 3 that could be used as a source focused on obtaining new potentially bioactive components of high biological value.
For these reasons, in the present study mojarra of Nile tilapia (O. niloticus) skeleton was used as a protein source for obtaining protein hydrolysates and peptide fractions with inhibitory activity of the ACE.
Materials and Methods
Materials
The mojarra of Nile tilapia (O. niloticus) was acquired in a tank nursery with plastic membranes from the city of Loma Bonita, Oaxaca, Mexico. The reagents used were of analytical grade and were acquired from the brands J.T. Baker (Phillipsburg, NJ, USA), Bio-Rad Laboratories, Inc. (Hercules, CA, USA), and Merck (Darmstadt, Germany). The enzymes Alcalase® and Flavourzyme® were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Mojarra tilapia skeleton flour
To obtain the flour, the method reported by Petenuci et al. 7 was used with some modifications. The mojarras of Nile tilapia were washed with purified water and the scales, viscera, and fillets were removed, and the obtained skeletons were washed with distilled water and they underwent a scalding (ratio 33.3 g/100 mL) for 50 min at 80°C, then another washing was performed with distilled water and the skeletons were cleaned manually to remove the residual meat and subsequently dried in a convection oven (RIOSSA mark) at 65°C for 24 h. After drying, the skeletons were milled in a coffee mill (Krupps Model GX4100; Millville, NJ, USA) until the flour was able to pass through a No. 40 mesh (effective aperture 0.425 mm) was obtained.
Chemical analysis and energy value
The chemical analysis of flour was performed according to the official methods described by AOAC, 9 which comprise the following analysis: moisture (method 925.09), crude protein (method 954.01), crude fat (method 920.39), and ashes (method 923.03). The energy value (kcal/g) was calculated by multiplying the protein content by four and the total fat by nine and adding these results. Subsequently, the data were multiplied by 4.184 to convert them into kilojoules per gram. 7
Color determination
The color was determined using an UltraScan® colorimeter (HunterLab, Hunter Associates Laboratory, Inc., Reston, VA, USA). The parameters measured were L* (luminosity), a* represents the chromatic scale from −green to +red (redness), and b* represents the chromatic scale from −blue to +yellow (yellowing). 10
Proteolytic enzymes
Enzymes used were as follows: Alcalase, a preparation of commercial proteases endoprotease activity produced by Bacillus licheniformis. 11 Its specific activity is 2.4 Anson Units (AU) per gram. An AU is defined as the amount of enzyme that, under standard conditions, digests hemoglobin at an initial rate that generates a quantity of soluble product of trichloroacetic acid (TCA) that produces the same color with the Folin reagent as a milliequivalent of tyrosine released per minute. The specific activity of Alcalase for this study was adjusted to 0.3 AU/g by dilution with distilled water. Furthermore, the Flavourzyme enzyme is a complex of proteases from Aspergillus oryzae having endo- and exoprotease activities. Its activity was 1.0 leucine aminopeptidase units (LAPU)/g. One LAPU is defined as the amount of enzyme that hydrolyzes 1 μmol of p-nitroanilide leucine per minute. In this study Flavourzyme was adjusted to 50 LAPU/g by dilution in distilled water. 12
Enzymatic hydrolysis
For enzymatic hydrolysis of Nile tilapia (O. niloticus) skeleton flour the method described by Hou et al. 13,14 was used with some modifications. A suspension of the flour was performed (5 g of protein in 100 mL of water). Different reaction times (0, 20, 40, 60, 80, and 100 min) were evaluated, hydrolysis was performed by individual treatments with each of the aforementioned enzymes, and the response variable was the percentage of the degree of hydrolysis (%DH). The digestion was carried out in a 100 mL beaker equipped with a magnetic stirrer, a thermometer, and a potentiometer, with an enzyme substrate ratio of 10% v/v, which was based on the protein content in the meal. The pH = 8.0 was adjusted to 60°C for Alcalase and pH = 7.0 to 50°C for Flavourzyme. Samples at 20, 40, 60, 80, and 100 min were taken to determine %DH, respectively, to finish the hydrolysis proteases heating at 85°C for 10 min and the inactivated hydrolysates were filtered through a filter 0.45 μm to remove the insoluble substrate and residual enzyme. The filtered material was stored frozen at −40°C in an ultrafreezer (Model No. DF8517; IlShin BioBase Co. Ltd., Korea) for further analysis.
Degree of hydrolysis
The %DH was determined using the method of Kim et al.
15
and reported by Qian et al.
16
This value was estimated by the amount of soluble nitrogen in TCA at 10% and this ratio versus the amount of total nitrogen in the protein suspension is as given in the equation:
Electrophoresis (sodium dodecyl sulfate–polyacrylamide gel electrophoresis)
Electrophoresis was performed in polyacrylamide gel with sodium dodecyl sulfate (SDS-PAGE) and the method of Laemmli 17 was followed with some modifications. Protein hydrolysates were mixed in a 1:2 (v/v) ratio solution with a buffer: 20% SDS, 2.5% glycerol, 1% β-mercaptoethanol, 0.02% bromophenol blue, and deionized water. The gel system consisted of 0.21% SDS and 20% acrylamide resolving (pH 8.8), and 5% acrylamide stacking gel (pH 6.8). Total gel thickness was 0.75 mm, with 10 cm of resolving gel and 2 cm of stacking gel. Protein bands were stained by gel immersion in Coomassie brilliant blue R-250 (Cat: 161-0400; Bio-Rad, Richmond, CA, USA) in a solution of 40% methanol, 7% acetic acid, and 53% deionized water. Broad range protein molecular weight (MW) standards were used (Cat: 161-0317; Bio-Rad), conformed by protein standards: myosin: 200 kDa, β-galactosidase: 116.25 kDa, phosphorylase b: 97.4 kDa, albumin: 66.2 kDa, ovalbumin: 45 kDa, carbonic anhydrase: 31 kDa, trypsin inhibitor: 21.5 kDa, lysozyme: 14.4 kDa, and Aprotinin: 6.5 kDa. Gel images were generated with the Gel Doc 1000 Image Analysis System (Bio-Rad) and analyzed with the Molecular Analyst Software (Bio-Rad).
ACE inhibitory activity
The ACE inhibitory activity (in vitro) was determined in all the protein hydrolysates obtained with Alcalase and Flavourzyme at different reaction times and in peptide fractions of the hydrolysates that had the best ACE inhibitory activity and where the highest DH was reached. The method used was reported by Hayakari et al.
18
with some modifications. In an ACE-I-containing hydrolysate, hippuryl-
Peptide fractions by ultrafiltration
The hydrolysates that reached the highest DH and highest ACE inhibitory activity were fractionated using a high-performance ultrafiltration (UF) cell (Model 2000; Millipore, Marlborough, MA, USA) according to Cho et al. 19 Five fractions were prepared using four membranes with MW cutoffs (MWCO): 1, 3, 5, and 10 kDa. Soluble fractions were prepared by centrifuging the hydrolysates through the MWCO membranes beginning with the largest cartridge (10 kDa). The retentate and permeate were collected separately, and the retentate was recirculated into the feed until the maximum permeate yield was reached, as indicated by a decreased permeate flow rate. Permeate from the 10 kDa membrane was then filtered through the 5 kDa membrane with recirculation until maximum permeate yield was reached. The 5 kDa permeate was then processed with the 3 kDa membrane and the 3 kDa permeate with the 1 kDa membrane. This process minimized contamination of the larger MW fractions with smaller MW fractions while producing enough retentates and permeates for the following analysis. The five ultrafiltered peptide fractions were prepared and designated as fraction A: >10 kDa (10 kDa retentate); fraction B: 5–10 kDa (10 kDa permeate–5 kDa retentate); fraction C: 3–5 kDa (5 kDa permeate–3 kDa retentate); fraction D: 1–3 kDa (3 kDa permeate–1 kDa retentate); and fraction E: <1 kDa (1 kDa permeate).
Statistical analysis
All results were analyzed using descriptive statistics with a central tendency and dispersion measures. One-way analysis of variance was applied to evaluate protein-rich flour hydrolysis data and in vitro ACE inhibitory activity, and a Duncan multiple range test was used to determine differences between treatments, with a significance level of 5%. All analyses were carried out according to Montgomery, 20 using the Minitab version 16 software.
Results
Proximal chemical composition and color
Flour was obtained from the mojarra of Nile tilapia (O. niloticus) complete skeleton, with a yield of 45.97% of the total fish used. The proximal chemical composition of the flour is given in Table 1, showing a protein and fat content of 34.92% and 5.2%, respectively. On the contrary, the flour in the study showed an ash content of 40.18% and the energetic value of the skeleton flour of the mojarra of Nile tilapia evaluated was 780.23 kJ (186.48 kcal/100 g).
Chemical Analysis, Energy Value, and Color Parameters from Mojarra of Nile Tilapia (Oreochromis niloticus) Skeleton Flour
The data are the average of three replicates ± standard deviation.
Except moisture, all other components are on dry basis.
The color parameters influence the general acceptability of food products; therefore, the determination of this parameter is necessary to dispose of its use in foods for human consumption. The skeleton mojarra of Nile tilapia flour (O. niloticus) rich in protein (Table 1) showed a luminosity value (L*) = 82.29, a redness value (a*) = 0.47, and a yellowing value (b*) = 15.26.
Degree of hydrolysis
Enzymatic hydrolysis with Flavourzyme and Alcalase enzymes (Fig. 1) at different reaction times managed to obtain extensive hydrolysates (DH >10%). DH obtained with Flavourzyme enzyme was 34–52% showing a statistically significant difference (P < .05) with respect to the enzyme Alcalase with DH of 41–67%. All the curves showed a rapid initial phase during which a large number of peptide bonds were hydrolyzed and subsequently this rate of hydrolysis decreased and a stationary phase was reached, where the hydrolysis apparently did not take place, forming a plateau after 80-min reaction with both enzymes.

Percentage DH of the protein hydrolysates obtained from mojarra of Nile tilapia (Oreochromis niloticus) skeleton flour rich in proteins at different reaction times with the enzymes Flavourzyme® and Alcalase®. The data are the average of three replicates ± standard deviation. a-dDifferent letters in the same kinetics indicate significant difference (P < .05). DH, degree of hydrolysis.
Electrophoresis (SDS-PAGE)
The electrophoretic pattern of protein hydrolysates obtained with Flavourzyme and Alcalase are given in Figure 2. In both gels, lane 1 shows the standard of high MW proteins, in lane 2 the skeletal of Nile tilapia flour without hydrolysis (time 0) is shown, and in lanes 3–7 are the hydrolysates obtained at different reaction times. In lane 2, protein bands between 200 and 14.4 kDa were observed, whereas lanes 3–7 show bands between 115 and 12.45 kDa (Fig. 2a) of the hydrolysates with Flavourzyme. On the contrary, the hydrolysates with Alcalase in lane 2 protein bands between 200 and 14.4 kDa were observed, and in lanes 3–7 there were bands between 97 and 18 kDa (Fig. 2b).
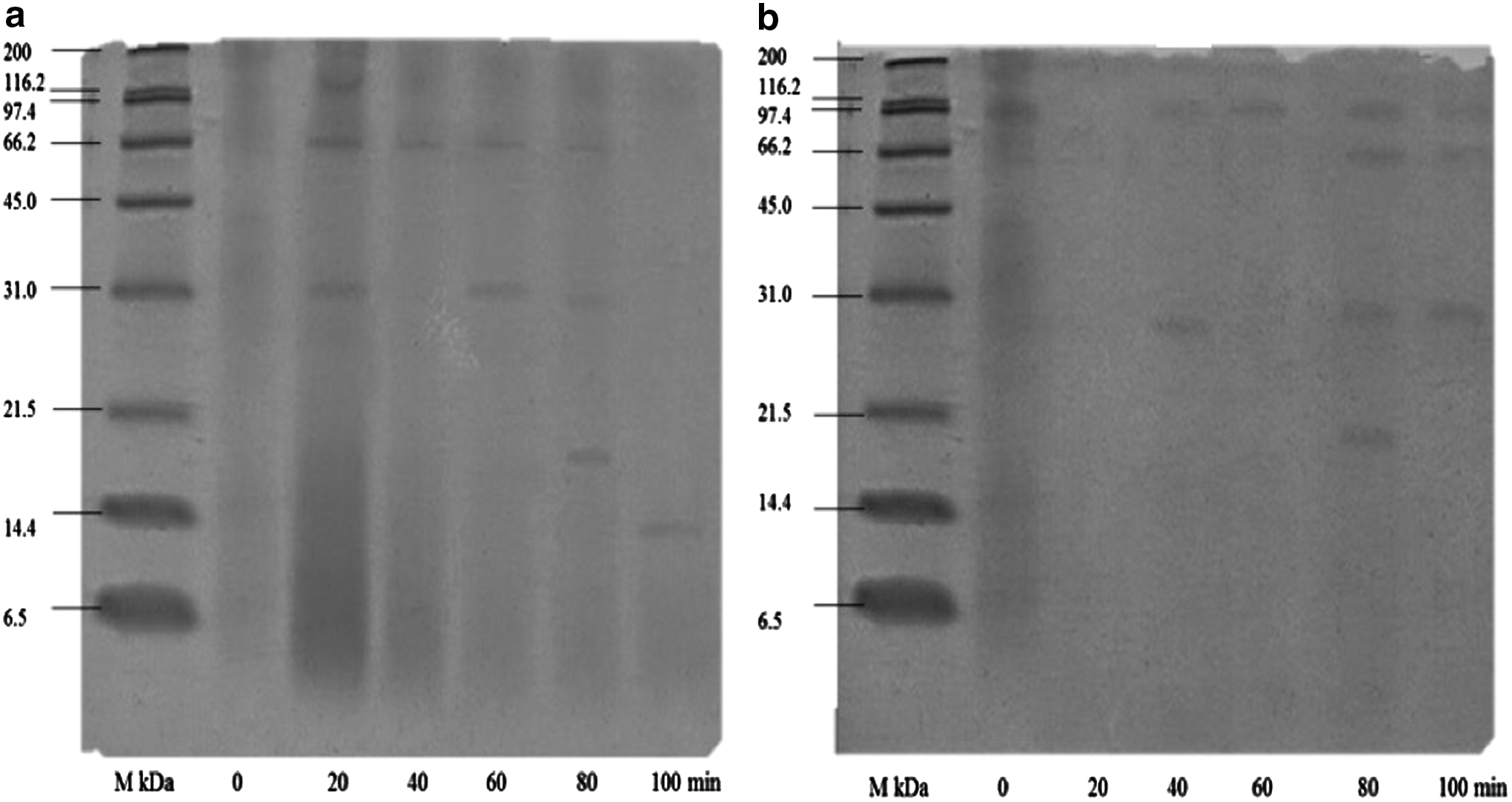
Electrophoretic pattern of protein hydrolysates obtained from mojarra of Nile tilapia (Oreochromis niloticus) skeleton flour rich in proteins at different reaction times with the enzymes
In vitro ACE inhibitory activity of protein hydrolysates
The ACE inhibitory activity of the protein hydrolysates of O. niloticus meal obtained with Flavourzyme and Alcalase are given in Table 2. The hydrolyzate obtained with Flavourzyme at 60 min of reaction showed a value of IC50 = 0.238 mg/mL, followed by hydrolyzate at 80 min with an IC50 = 0.298 mg/mL and the obtained at 20, 40, and 100 min of reaction with a range of IC50 values of 0.527–0.926 mg/mL, showing significant statistical difference (P < .05) with the hydrolysates obtained with Alcalase; the hydrolyzate at 80 min had greater activity with IC50 = 0.344 mg/mL, whereas the hydrolysates at 20, 40, 60, and 100 min the interval of IC50 was 0.400–0.891 mg/mL.
Angiotensin-Converting Enzyme Inhibitory Activity In Vitro of Protein Hydrolysates Obtained from Mojarra of Nile Tilapia (Oreochromis niloticus) Skeleton Flour
The data are the average of three replicates ± standard deviation.
Lowercase letters different in the same column indicate significant difference (P < .05).
Uppercase letters different in the same row indicate significant difference (P < .05).
In vitro ACE inhibitory activity of the peptide fractions obtained by UF
Hydrolysates selected to be fractionated by UF were those that presented higher DH and greater ACE inhibitory activity, the treatments being as follows: hydrolyzed with Flavourzyme at 60 and 100 min, and the hydrolysates with Alcalase at 80 and 100 min, respectively (Table 3). Peptide fractions of the hydrolysates with Flavourzyme 100 min showed the highest activity in fraction A (MW >10 kDa) with an IC50 value = 0.728 mg/mL, fraction B (MW = 10.05 kDa) and fraction C (MW = 5–3 kDa) with IC50 values = 0.755 and 0.761 mg/mL, respectively, whereas the lowest activity was in fractions D and E (MW = 3–1 kDa and MW <1 kDa, respectively). On the contrary, the peptide fractions of the hydrolysates with Alcalase with the best activity was fraction B (MW = 10–5 kDa) with an IC50 = 0.354 mg/mL, whereas for peptide fractions A, C, D, and E (MW >10 kDa, MW = 5–3 kDa, MW = 3–1 kDa, and MW <1 kDa, respectively), the range of IC50 values was 1.13–7.08 mg/mL.
Angiotensin-Converting Enzyme Inhibitory Activity In Vitro of the Peptide Fractions Obtained by Ultrafiltration from Protein Hydrolysates of Mojarra of Nile Tilapia (Oreochromis niloticus) Skeleton Flour
The data are the average of three replicates ± standard deviation.
Lowercase letters different in the same column indicate significant difference (P < .05).
Uppercase letters different in the same row indicate significant difference (P < .05).
ACE, angiotensin-converting enzyme; DH, degree of hydrolysis.
On the contrary, the ACE inhibitory activity shown by the peptide fractions of the hydrolysates had the highest activity (60 and 80 min for Flavourzyme and Alcalase, respectively) (Table 3). The highest inhibitory activity of the ACE for the peptide fractions with both enzymes evaluated was fraction C (MW = 5–3 kDa) with a value of IC50 = 0.470 and 0.634 mg/mL, respectively, whereas for fractions A, B, D, and E (MW: >10, 10–5, 3–1, and <1 kDa) the activity was in the range of IC50 values between 0.685 and 1.62 mg/mL, respectively.
Discussion
The data yield was similar to those found by Petenuci et al. 7 where they had a yield of 41% backbone flour of Nile tilapia. For protein these values were higher than those reported by Visentainer et al. 21 for head flour tilapia with 14.1% and by Hou et al. 14 with values of 18.36% and 18.8% for bone structure and Alaska pollock meat, respectively. Nguyen et al. 22 reported protein values of 14.8%, 16.0%, and 17.4% in by-products (head, viscera, and tail) of yellowfin tuna (Thunnus albacares), and Naqash and Nazee 23 reported values of 12.1% in tropical flying fish backbone (Exocoetus volitans). However, other authors such as Petenuci et al. 7 and Liaset et al. 24 found higher protein values in backbone flour of Nile tilapia of 40.8% and in the structure of Atlantic salmon (Salmo salar L.) of 48%. The fat content found in this study was within the range of 0.6–25.3% of reported fat content for various products and fishery by-products. 7,13,25 These variations in the composition may be because of the feeding of the fish, size of fish, sex, age, geographical area of their catch, and the time of year they were caught. 26 The ash content was higher than values reported by Visentainer et al. 21 with 4.76% in heads tilapia and by Gbogouri et al. 27 found in fresh salmon heads triturated with a content of 2.6%.
Petenuci et al. 7 had values of 18.3% in backbone flour of Nile tilapia. This may be because of variation in the method of obtaining the flour and drying the same as this reduced the water content that caused an increase in the concentration of other components as mentioned by Petenuci et al. 7 It should be noted that the high ash content of the flour in the study could be a rich source of minerals and trace elements that would give an added value to this by-product to be used as an ingredient in some edible product for animals. The energy values can be compared with the energy values of 1636 kJ (391 kcal/100 g) found by Petenuci et al. 7 in backbone tilapia flour. The values found in this study suggest that it could be added to different dishes such as soups and broths, helping them to significantly increase their energy value.
The L* value found in this study was similar to that reported by Šližytė et al. 28 with a value of L* = 85.7 in protein hydrolysate obtained with the enzyme Protamex to 10 min of reaction of the complete skeleton of cod (Gadus morhua), whereas Elavarasan et al. 29 reported values of L* = 89.32 and 80.16 in freshwater carp (Cirrhinus mrigala) hydrolysate freeze-drying and oven, respectively. On the contrary, Hou et al. 13 found a lower luminosity in defatted protein horse mackerel (L* = 72.52). The parameters a* and b* showed that the tilapia flour evaluated was less red (a* = 0.47) and yellow (b* = 15.26) than those found by other authors such as Sathivel et al., 30 who obtained values of a* (9.0–9.9) and b* (30.0–30.2) in head hydrolyzates of red salmon (Oncorhynchus nerka) with the Alcalase and Flavourzyme enzymes, respectively.
This may be because of the fact that the species evaluated showed no retention of lipids and fat-soluble pigments as a result of the treatment to which the material was subjected to during the obtaining of flour. However, Dong et al. 31 mentioned that when there is a greater reddening and yellowing it may be because of the oxidation of pigments and Maillard reactions when subjected to an enzymatic hydrolysis, or could be because of the release of hemoglobin from the spines when they are subjected to a cut as mentioned by Šližyte et al. 28
Similar behaviors in enzymatic hydrolysis curves of protein products and fishery by-products have been reported by Nguyen et al. 22 in hydrolyzed by-products of yellowfin tuna (head, viscera, and tail), by Guerard et al. 32 from the stomach of yellowfin tuna, and by Kristinson and Rasco, 33 in muscle and head of salmon. On the contrary, Klompong et al. 34 analyzed the influence of the enzyme concentration on the degree of hydrolysis from muscle yellowstripe scad (Selaroides leptolepis) using Flavourzyme and Alcalase at concentrations of 0.25–10% and obtained the highest DH using an enzyme-substrate concentration of 10% for each enzyme, where DHs were 25% and 45% in the first 20 min of reaction, respectively.
These results are comparable with DH obtained in this study after 20-min reaction because with the same enzymes DH 34% and 41%, respectively, were obtained using the same enzyme/substrate ratio of 10%. It should be mentioned that the highest DH was obtained with Alcalase and this may be because of the fact that this enzyme has an endoprotease activity, so it has a broad specificity to peptide bonds hydrolyze from C-terminal end of the peptide chain, causing the release of peptides containing hydrophobic amino acid residues such as Phe (F), Tyr (Y), Trp (W), Leu (L), Ile (I), Val (V), and Met (M). 35 On the contrary, several authors report that alkaline proteases including Alcalase, show higher activity than acidic and neutral proteases such as Flavourzyme. 31,34
The results showed that as the DH increased, the production of low MW peptides increased, especially those with a molecular weight <31 kDa with both enzymes, achieving extensive hydrolysates and even some protein bands could not be retained by this method. “Similar results have been reported by several researchers as Raghavan et al. 8 who obtained peptides with Flavourzyme enzyme with a molecular weight <30 kDa from tilapia fillets and these peptides could not be retained by separating gel with the method of SDS-PAGE electrophoresis employed.
On the contrary, Theodore et al. 36 obtained hydrolysates from catfish with the Protamex® enzyme, an alkaline enzyme similar to Alcalase with DHs of 5%, 15%, and 30%. These authors reported a DH of 5% of heavy myosin chains (MW ∼200 kDa) which were completely hydrolyzed to low MW peptides and they observed bands of light chains (∼10–20 kDa) and actin (∼42 kDa). As the DH increased up to 30%, the number of bands corresponding to peptides between 20–45 kDa decreased significantly, since the bands <20 kDa disappeared and were not revealed by the SDS-PAGE method, indicating an extensive hydrolysis. Klompong et al. 37 obtained protein hydrolysates from yellow stripe trevally (S. leptolepis) with Alcalase and Flavourzyme with DHs of 5, 15, and 25% and reported that the hydrolysates with Flavourzyme (DH = 5%) found peptides or proteins with a MW > 60 kDa; however, in the hydrolysates with Alcalase, those peptides/proteins were not found, and when the DH was 25% with both enzymes, the peptides were not retained because of their low molecular weight.
These results of ACE inhibitory activity are within those reported by other authors such as Matsui et al. 38 who found IC50 values between 0.3 and 1.2 mg/mL from sardine muscle hydrolysates; Nakajima et al. 39 reported a value of IC50 = 0.078 and 0.138 mg/mL, respectively, in hydrolysates of Atlantic salmon and coho salmon (silver) using the enzyme Termolysin®; Balti et al. 40 obtained with cuttlefish (Sepia officinalis) using several proteolytic enzymes such as Alcalase and reported values of IC50 = 1.84 mg/mL; and Elavarasan et al. 29 reported IC50 values of 1.15 and 1.53 mg/mL for inhibition of the ACE in protein hydrolysates of freshwater fish (C. mrigala) dried by lyophilization and oven, respectively. The difference found in the inhibitory activity of the ACE could be because of the fact that during the course of the hydrolysis peptides are formed that show ACE inhibitory activity and these are targets of the enzymes still present in the medium and can be again hydrolysates, thereby causing a modification in the composition and amino acid sequencing of the said components, as mentioned by Pedroche et al. 12
Similar results were found by Samaranayaka et al. 41 who obtained hydrolysates from Pacific hake (Merluccius productus) using two Pepsin® and Pancreatin® digestive enzymes, and these were fractionated by UF with membranes of 10, 3, and 1 kDa, and reported an IC50 = 0.883 mg/mL for the fractions >10 kDa that had less ACE inhibitory activity obtained in this study for the fraction of the same MW obtained with the Flavourzyme enzyme.
The values found in this study can be compared with values reported by Jung et al. 4 from protein hydrolysates of the fish yellowfin sole (Limanda aspera) skeleton obtained with α-Chimotrypsin® that were fractionated with membranes of different molecular sections of 30, 10, and 5 kDa, and found a value of IC50 = 0.883 mg/mL for the fraction <5 kDa, because in this study similar values were found in the fractions of low MW (fractions C and D with Flavourzyme to 60 min and the fractions C, D, and E with Alcalase to 80 min).
Salampessy et al. 42 isolated and characterized peptide fractions from the skin of fish leatherjacket (Meuchenia sp.) from hydrolysates with Flavourzyme and obtained IC50 values between 1.77 and 2.04 mg/mL in the peptide fractions <5 kDa and the fractions of 5–10 kDa IC50 values were between 1.86 and 3.05 mg/mL; these activity values were lower than this study and the protein-rich tilapia flour evaluated could be a good source of peptides with inhibitory activity of the ACE. These changes in the inhibitory activity could be the result of the amino acid composition and sequence on bioactive peptides present in the different peptide fractions obtained, because these amino acids have ACE inhibitory activity, and that the said activity will not depend on the size of the peptide fractions but of the aforementioned factors, and the protein source, the enzymatic system, and operating conditions used in this study. 43,44
In conclusion, the use of the enzymes Flavourzyme and Alcalase individually achieved extensive hydrolysates from the protein-rich flour of the mojarra of Nile tilapia (O. niloticus) skeleton. The electrophoretic patterns obtained by SDS-PAGE of these hydrolysates showed low MW protein bands, thus suggesting the presence of bioactive peptides on hydrolysates. The hydrolysates showed ACE inhibitory activity in all their reaction times, the hydrolyzate having greater activity with Flavourzyme at 60 min and with Alcalase at 80 min of reaction. On the contrary, the peptide fractions obtained by UF that showed greater activity were peptide fraction C (5–3 kDa) with both enzymes. So they could have applications as a functional ingredient in the design and development of specialized foods.
Footnotes
Acknowledgments
The author Berenise Borges-Contreras (CVU: 392816) thanks the Consejo Nacional de Ciencia y Tecnología (CONACYT) for the scholarship granted to study the Master's Degree in Food Sciences of the Technological Institute of Tuxtepec. Likewise, the author thanks the PROMEP (today PRODEP) for financing the project entitled: Antihypertensive and antioxidant activity of peptide fractions obtained from the protein hydrolyzate of the tilapia fish skeleton (O. niloticus). Project key: PROMEP/103.5/10/5379.
Author Disclosure Statement
No competing financial interests exist.