
Abstract
This study focused on the evaluation of the chemopreventive potential of tissue in vitro culture of the “Mela Rosa Marchigiana” apple (MRM callus) that allows the amplification of secondary metabolites. The MRM pulp and MRM callus chemopreventive potential was evaluated in terms of antiproliferative activity, inhibition of tumorigenesis in soft agar cultures, cell cycle and western blotting analyses in CaCo2 and LoVo colon cancer cell lines and in JB6 promotion-sensitive (JB6 P+) cells. MRM callus induced a strong concentration-dependent inhibition of colon cancer cell proliferation and suppressed 12-o-tetra-decanoyl-phorbol-13-acetate-induced tumorigenesis of JB6 P+ cells in soft agar cultures. MRM callus inhibited the phosphorylation of JNK, p38, and eIF2alpha. Our data indicate that the MRM callus exerts a good antiproliferative and antitumorigenic potential through the MAP kinase inhibition and could provide natural compounds with chemopreventive properties.
Introduction
Cancer is a complex disease in which genetic and environmental exposure and lifestyle serve to initiate tumor development and progression. 1 For this reason, researchers focused on chemoprevention, the use of synthetic or natural agents to inhibit, prevent, or slow induction and progression of cancer. 2
Colon cancer is the third most prevalent cause of cancer death in the world; due to the long precancerous stage of colorectal cancer and to the direct contact between cancer cells and chemopreventive dietary agents, chemoprevention could be a very promising strategy. Several studies considered the role of fruit and vegetable nonnutrient compounds in chemoprevention, to test their abilities to interfere with tumorigenesis. 3 Apple consumption counteracts various degenerative disorders, such as ischemic heart disease, inflammation, and cancer. 4,5
The preventive effects of bioactive phytochemicals contained in apple juice has been demonstrated in colon cancer. 6,7 Apples or apple products (apple juice, extracts, dehydrated apples) could exert biological effects to prevent or reduce the proliferation of colorectal cancer, both in in vitro and in vivo models and in case–control studies, by which the risk of colorectal cancer risk resulted inversely correlated with the number of consumed apples. 8 –13 Those beneficial effects have been attributed to the apple's bioactive molecule mixture, which is more effective than a single bioactive molecule in exerting their biological activities. 14 –16
We have demonstrated that a specific apple is particularly rich in polyphenols and that its juice leads to the strongest antiproliferative activity against breast cancer cells, in comparison to the five other apple varieties, suggesting that polyphenols play an important role. 17
Although access to fresh fruits is determined by seasonal availability, drying processing techniques and in vitro culture (callus) induction allow for the extension of the shelf life of fruits. Callus production is used to amplify fruit and vegetable secondary metabolites that have beneficial effects for human health; several studies focused on the optimization of this technique to obtain bioactive components from a variety of natural foods. 18 –24
We have previously demonstrated that the callus derived from the pulp of the Italian “Mela Rosa Marchigiana” apple (MRM apple) (Fig. 1) produces a higher amount of triterpenic acids than those contained in the apple itself, in particular maslinic acid (MA) and tormentic acid (TA). 25

Triterpenoids exert antiproliferative activity against several human cancer cell lines. 26,27 and MA has a strong antiproliferative activity against human colon cancer cells, HT29 and CaCo2, activating apoptotis. 28,29
The aim of this study was to evaluate MRM callus and MRM pulp antiproliferative effects in CaCo2 and LoVo human colon cancer cell lines and in preneoplastic JB6 Cl 41–5a promotion-sensitive cells (JB6 P+ cells) and to investigate MRM callus antitumorigenic potential in JB6 P+ cells.
Materials and Methods
Cell cultures and chemical reagents
Human colorectal adenocarcinoma cell lines, CaCo2 and LoVo, and murine skin epidermal JB6 Cl 41–5a promotion-sensitive (JB6 P+) cells were purchased from the American Type Cell Culture Collection (ATCC, Rockville, MD, USA). Cells were cultured in Dulbecco's Modified Eagle Medium (CaCo2 and LoVo cells) or Eagle's Minimum Essential Medium (EMEM) (JB6 P+ cells) media, supplemented with 10% or 5% of heat-inactivated fetal bovine serum, respectively, and 2 mM L-glutamine, 1 × MEM Nonessential Amino Acid Solution, 0.1 mg/mL streptomycin, 0.1 U/L penicillin, and 1 mM Na-pyruvate. The 12-o-tetradecanoylphorbol-13-acetate (TPA) was dissolved in dimethyl sulfoxide.
All cell culture materials and Maslinic Acid ((2α,3β)-Dihydroxyolean-12-en-28-oic acid, MA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). The TA, obtained from the MRM callus, has been kindly provided by ABR (Active Botanicals Research) (Brendola, VI, Italy).
Plant material and tissue culture
MRM apples were collected from an orchard in the Marche region of Italy.
After sterilization with ethanol 90%, apples were opened under the laminar flow cabinet with a sterile blade and cultures were conducted as previously described. 25 Cultures were incubated in the dark at 25°C ± 2°C and subcultures were obtained after 28 days in the same media. The callus obtained by subcultures (Fig. 1B) was stored at −80°C and lyophilized before the extraction.
Extraction of MRM callus and MRM pulp
Lyophilized MRM pulp and MRM callus was ground into powder and extracted in sterilized Milli-Q H2O (1.94 g in 8.95 mL and 1.2 g in 18.4 mL, respectively) under magnetic stirring, followed by centrifugation at 3600 g for 15 min at 4°C. The supernatants were filtered with a 0.45 μm membrane, aliquoted, and stored at −20°C.
Anchorage-dependent growth assay
CaCo2 and LoVo cells (1 × 104 cells/well) and JB6 P+ cells (2 × 104 cells/well) were seeded in 96-well plate; after overnight incubation, cells were treated in triplicate with MRM pulp, MRM callus, MA, and TA (only CaCo2 and LoVo cells) at final concentrations of 0–5% v/v of MRM pulp and MRM callus, 0–50 μg/mL of MA and 0–100 μg/mL of TA. JB6 P+ cells were stimulated with 10 ng/mL TPA. After 72 h of treatment, cell viability was evaluated using the CellTiter 96 Aqueous Non-Radioactive Cell Proliferation Assay (Promega, Madison, WI, USA), as previously reported. 17
Anchorage-independent transformation assay
JB6 P+ cells were cultured in 35-mm dishes (1 × 104 cells/dish) to investigate MRM pulp, MRM callus, and MA effects on TPA-induced cell transformation. The intermediate agar layer was composed of 0.3% agar–10% FBS-EMEM and cells, with different concentrations of MRM pulp, MRM callus (0%, 0.10%, 0.5%, 5% v/v), and MA (0, 0.3, 1.5, 15 μg/mL), with a bottom layer of 0.6% agar–10% FBS-EMEM. Cells were maintained in an incubator for 21 days, stained with 0.1% Crystal Violet, counted, and photographed with an inverted microscope.
Flow cytometric analysis
CaCo2 cells were seeded in six-well plates (5 × 105 cells/well) and then treated with 2.5% and 5% v/v of MRM pulp, 2.5 and 5% v/v MRM callus, or with 12.5 and 25 μg/mL MA, for 24 h. After treatment, cell cycle analysis was performed as previously reported. 17 Cytofluorimetric acquisitions and sample analysis were performed with a Partec PAS flow cytometer (Partec, Münster, Germany) and FlowJo 8.6.3 software (TreeStar, Inc., Ashland, OR, USA), respectively.
Immunoblot analysis
CaCo2 cells were seeded in six-well plates (5 × 105 cells/well) and treated for 4 and 8 h with MRM pulp, MRM callus, or MA. JB6 P+ cells were seeded in six-well plates (2.5 × 105 cells/well), starved in 0.1% FBS-EMEM for 24 h, and then subjected to a 1-h pretreatment with MRM pulp, MRM callus, or MA, after which were stimulated with TPA 10 ng/mL for 30 min. After treatments, cells were lysed for western blotting analysis as previously reported. 30
The following primary antibodies have been used: phospho-p38 MAPK, p38 MAPK, phospho-SAPK/JNK, SAPK/JNK, phospho-p44/42 MAPK (ERK1/2), p44/42 MAPK (ERK1/2), phospho-eIF2alpha, and Cleaved Caspase-3 (Cell Signaling Technology, Beverly, MA, USA). Protein bands were detected incubating blots with horseradish peroxidase-conjugated secondary antibody and then with Clarity™ Western ECL Substrate (Bio-Rad Laboratories, Inc., Hercules, CA, USA); the immunoreactive bands were detected with chemiluminescence film (Amersham Hyperfilm ECL; GE Health Care, Little Chalfont, United Kingdom). Equal proteins loaded were confirmed using anti-Actin antibody (Sigma-Aldrich, St. Louis, MO, USA).
Statistical analysis
All the data are expressed as mean ± SEM of at least three different experiments. Significant differences between the mean values were analyzed using Prism5 software, with 1- or 2-way ANOVA followed by Dunnett's or Bonferroni post-hoc tests. P-values minor than .05 were considered as significant.
Results
MRM pulp and MRM callus antiproliferative effects in human colon cancer cells
Anchorage-dependent growth inhibition MTS assay was performed in CaCo2 and LoVo cells. As shown in Figure 2A, in both cancer cell lines MRM pulp and MRM callus led to a concentration-dependent reduction of cellular proliferation; MRM callus significantly reduced the proliferation of CaCo2 and LoVo cells starting from a concentration of 0.625% v/v (P < .001) and from 1.25% v/v (P < .05), respectively. Moreover, the MRM callus was more effective than MRM pulp (P < .001), leading to a IC50 of 3.7 ± 0.5 or 6.6% ± 0.6% v/v in CaCo2 and LoVo, respectively (Table 1).
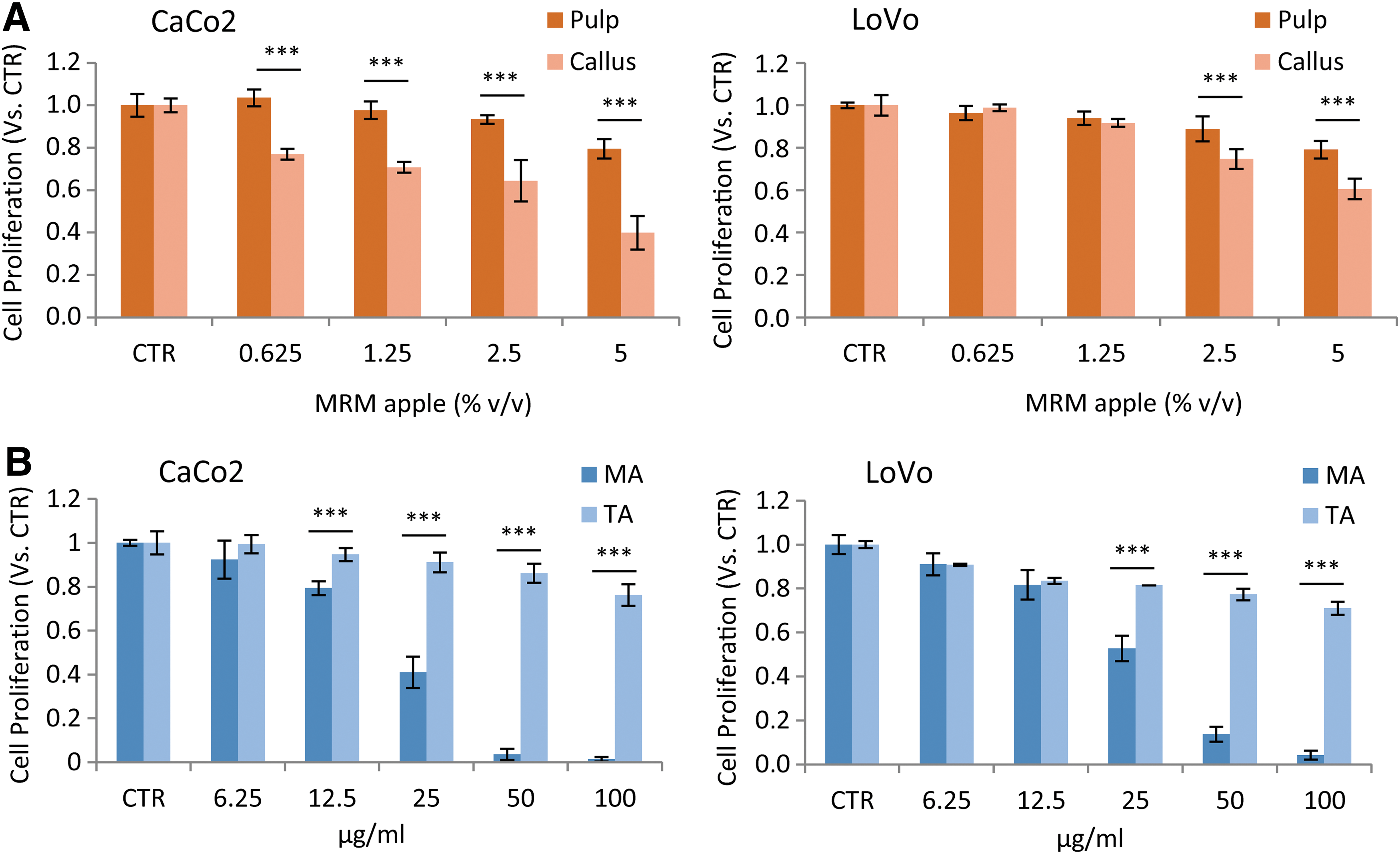
Effect of MRM pulp, MRM callus, MA, and TA on CaCo2 and LoVo cell proliferation. CaCo2 and LoVo cells were treated with MRM pulp, MRM callus
IC50 Values
Half maximal inhibitory concentrations of MRM pulp, MRM callus (% v/v) MA and TA (μg/mL) obtained in CaCo2 and LoVo cells.
(IC50) were calculated by nonlinear regression analysis (see Anchorage-dependent growth assay section)
MRM, Mela Rosa Marchigiana; MA, maslinic acid; n.c., not calculable; TA, tormentic acid.
Based on previously reported chemical analysis, 25 the amounts of MA and TA–the most representative MRM callus triterpenic acids–contained in 3.7% and 6.6% v/v of MRM callus were 11.3 and 20 μg/mL of MA and 33.6 and 59.6 μg/mL of TA, respectively.
To understand whether MA and TA were responsible for MRM callus antiproliferative effects in the colon cancer cell lines tested, different concentrations of MA and TA were considered (Fig. 2B). MA resulted in a concentration-dependent reduction of cell proliferation: the obtained MA IC50 was of 20.9 ± 0.8 and 24.9 ± 0.9 μg/mL (Table 1), 1.85-fold and 1.25-fold higher than the MA contained in the IC50 concentration of MRM callus, in CaCo2 and LoVo cells, respectively. The TA showed a lower antiproliferative activity than MA, inhibiting 23.8% and 29.0% of cell proliferation at the highest concentration tested (100 μg/mL) in CaCo2 and LoVo cells, respectively. For this reason only MA was considered for further experiments.
Analysis of cell cycle perturbations
The analysis of cell cycle was performed in CaCo2 cells. Results showed that only MA at a concentration of 25 μg/mL led to a perturbation of cell cycle, with an increased cellular population in the G1 phase, from 45.87% ± 2.19% of the control to 61.23% ± 0.81%, and a decreased cellular population in the S phase, from 33.67% ± 1.27% of the control to 18.63% ± 0.64% (P < .001) (Fig. 3A). These results showed that only MA produces significant changes in cell cycle profile; the link between MA-induced cell growth inhibition and cell cycle arrest in G1 phase was confirmed by the western blotting analysis of phosphorylated ERK 1/2, which resulted to be downregulated only by MA (Fig. 3B).

The apoptotic pathway involvement was assessed by hypodiploidy analysis, which revealed that MRM pulp and MRM callus did not induce alterations; these data were confirmed by the analysis of cleaved caspase-3 through western blotting, in which only MA activated the apoptotic pathway (data not shown).
MRM pulp and MRM callus antiproliferative effects in TPA-stimulated preneoplastic JB6 P+ cells
JB6 P+ cells were stimulated with 10 ng/mL TPA and simultaneously treated with different concentrations of MRM pulp or MRM callus for 72 h. Morphological examination of cells resulted in significant changes only in cotreated TPA-MRM callus cells, starting from a concentration of 1.25% v/v and being more evident at a concentration of 5% v/v (Fig. 4A). In non-TPA-stimulated cells, we found no significant proliferation reduction after treatment with MRM callus. Notably, MRM callus induced a reduction of 100% of TPA-stimulated proliferation at a concentration of 5% v/v (P < .001). On the other hand, MRM pulp only partially reduced JB6 P+ cell proliferation and TPA-induced proliferation (Fig. 4B).
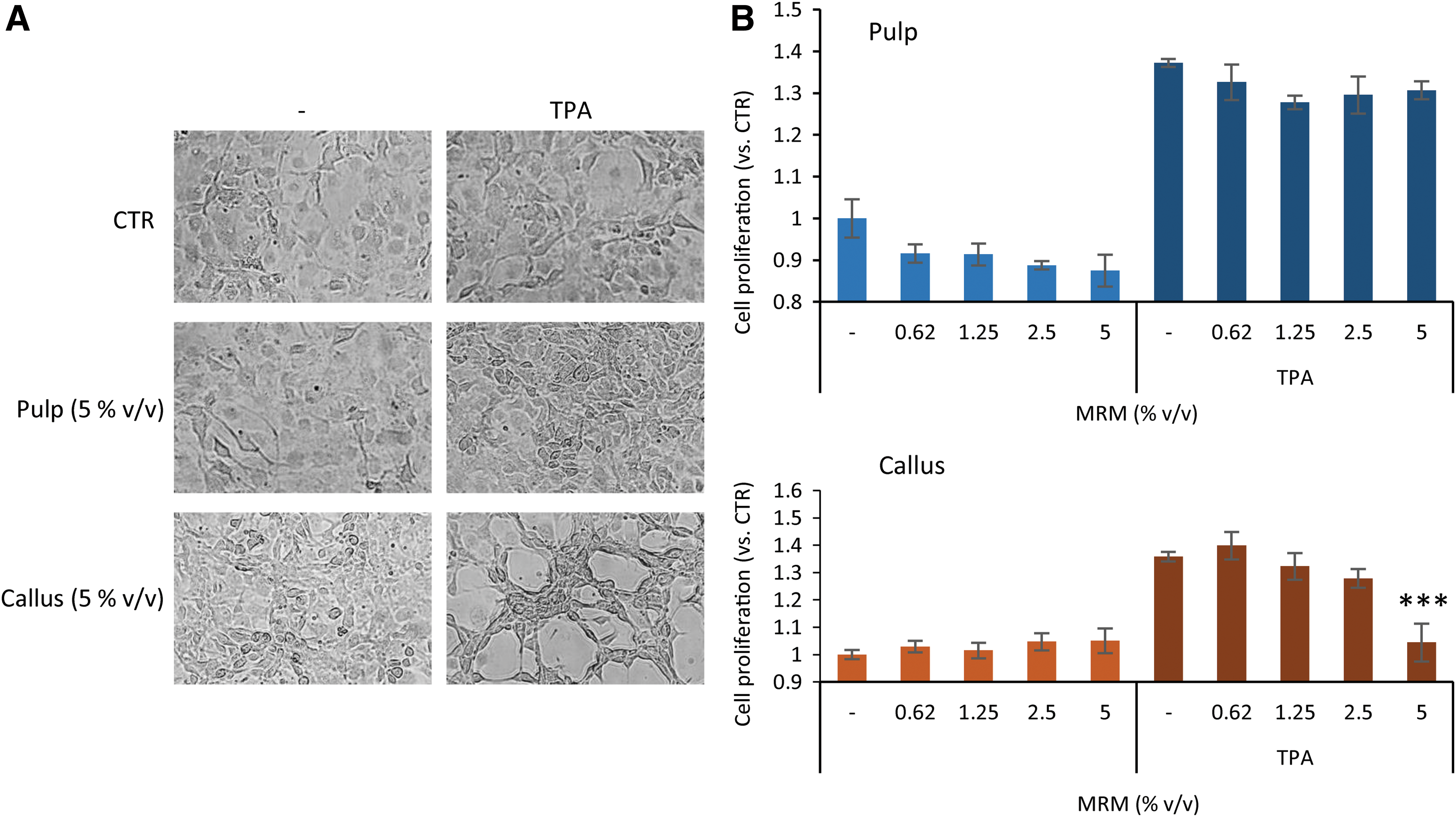
Effect of MRM pulp, MRM callus, and MA on JB6 P+ cell proliferation. Morphological examination
MRM pulp and MRM callus effects on TPA-induced JB6 P+ cell transformation
The ability of JB6 P+ cells to form colonies in anchorage-independent cultures under different treatments was monitored (Fig. 5A). Results showed that both MRM pulp and MRM callus significantly decreased JB6 P+ cell's colony formation ability in TPA presence or absence, starting from a concentration of 0.10% v/v. Interestingly, MRM callus was more effective than MRM pulp: MRM pulp resulted in a reduction of colony number of about 7.5% in nonstimulated cells and 34.8% in TPA-stimulated JB6 P+ cells at 0.1% v/v; moreover, to inhibit more than 90% of colony formation, a concentration of 5% v/v is necessary (P < .01 and P < .001, respectively); MRM callus led to a reduction of about 81.9% in nonstimulated cells and completely inhibits colony formation in TPA-stimulated JB6 P+ cells at 0.1% v/v.

Effect of MRM pulp, MRM callus, and MA on in vitro tumorigenesis of JB6 P+ cells. Anchorage-independent cell growth assay. Cells were treated with MRM pulp, MRM callus (0–5% v/v), or MA (0–15 μg/mL), stimulated with TPA (10 ng/mL), cultured in soft agar for 21 days, and colonies containing more than 20 cells were counted.
Furthermore, to understand whether the MA was responsible for the effect of MRM callus in the suppression of TPA-induced colony formation, different concentrations of MA were tested in soft agar cultures. MA did not lead to statistically significant differences in colony number in nonstimulated cells (Fig. 5B); in TPA-treated cells, a final concentration of 0.3 μg/mL MA, corresponding to 0.1% v/v of MRM apple callus, did not have the same effect observed in MRM callus-treated cells, leading to a small reduction in colony number (P < .05) (Fig. 5B).
MRM pulp and MRM callus' effects on TPA-induced phosphorylation of MAPK pathways in JB6 P+ cells
MAP kinase phosphorylation was evaluated by western blotting analysis. Figure 6 shows that only MRM callus inhibited TPA-induced JNK phosphorylation and avoided the accumulation of phosphorylated p38. However, ERK phosphorylation level did not change in MRM pulp, MRM callus, or MA treatment. To better understand the involvement of p38 in MRM callus effects in JB6 P+ cells' growth inhibition in soft agar conditions, the phosphorylation of eIF2alpha was considered. Our results showed that TPA-induced phosphorylation of eIF2alpha was inhibited only by MRM callus, confirming the involvement of p38.

Western blot analysis of JB6 P+ pretreated with MRM pulp, MRM callus (5% v/v), or MA (15 μg/mL) for 1 h and stimulated with TPA (10 ng/mL) for 30 min. Phosphorylation of SAPK/JNK, ERK1/2, p38 MAPK (Thr180/Tyr182), and eif2alpha was analyzed. Total SAPK/JNK, p38, ERK1/2, and actin were used as loading controls for phospho-JNK, phospho-p38, phospho-ERK, and phospho-eIF2alpha, respectively. Densitometry values for specific proteins relative to unstimulated cells (set as one-fold) are included below the lanes.
Discussion
Several epidemiological studies suggested that increasing consumption of plant-based foods could be a strategy to prevent some chronic diseases, such as cardiovascular diseases, age-related function decline, and cancer. 31 Biologically active components in fruits and vegetables, particularly phytochemicals, nonnutrient plant compounds, such as flavonoids, phenolic acids, and carotenoids, can modulate many processes in the development of diseases, reducing their incidence. For these reasons, the scientific community has focused attention on micronutrients as potential cancer-preventive agents, studying diet-derived antioxidants in chemoprevention. 32
Apples represent a significant part of the European diet and, in Italy, are the most highly consumed fruit. Apples are rich in antioxidants and phenolic compounds and are a great source of polyphenols, flavonoids, and catechins, as well as triterpenoids and anthocyanins. 4
Given that fruits and vegetables contain biologically active components that can modulate processes of different diseases, several studies aimed to produce secondary metabolites useful to promote health, using in vitro cultures (callus) of fruit and plant tissues. 18 –20,22,23 We have previously demonstrated that in vitro cultures of the pulp of “Mela Rosa Marchigiana” apple (MRM callus) produces more triterpenic acids than those found in its pulp, resulting in an amplification of important secondary metabolites with antioxidant properties. 25
In this study we evaluated the antiproliferative activities of MRM callus in two human colon cancer cell lines (CaCo2 and LoVo cells), comparing them with the effects of MRM pulp. Our results demonstrated that MRM callus significantly inhibits CaCo2 and LoVo cell proliferation and that it is more effective than the pulp of the same apple. These results suggest that MRM pulp in vitro culture is a key to amplify the micronutrient content.
Apples' total triterpenoids have potent antiproliferative effects against human cancer cells 33 ; however, TA possesses lower CaCo2 permeability than MA. 34 Our results show that a low antiproliferative activity of TA in human colon cancer cells, maybe due to the low permeability of this compound. On the other hand, MA exerts an antiproliferative effect against human colon cancer cells 28,29 ; our results confirm this antiproliferative effects in CaCo2 and LoVo cells. Considering the antiproliferative effect given by the same amount of MA contained in MRM callus, MRM callus shows greater antiproliferative activity than MA alone, suggesting that the efficacy of MRM callus is not exerted only by MA, but that the whole MRM callus content contributes to the effect.
Moreover, MRM pulp and MRM callus effects on cell cycle perturbation were evaluated in colon cancer cell line CaCo2, showing that neither MRM pulp nor MRM callus impact cell cycle progression; on the contrary, MA leads to strong G1 phase cell accumulation and to a caspase-3 activation, confirming that MA is not the main compound exerting biological activity of MRM callus. Furthermore, MRM callus and MRM pulp antiproliferative activity was evaluated in the preneoplastic JB6 Cl 41–5a promotion sensitive (JB6 P+) cells, demonstrating that both MRM pulp and MRM callus are not toxic in preneoplastic nonstimulated JB6 P+ cells and that, interestingly, MRM callus completely inhibits TPA-induced JB6 P+ cell proliferation; MRM pulp, however, does not exert this effect.
To evaluate MRM pulp and MRM callus antitumorigenic potential, we considered their ability to inhibit JB6 P+ cell colony formation in anchorage-independent culture conditions (soft agar). Our results show that MRM callus blocks TPA-induced cell transformation, inhibiting colony formation in soft agar cultures; on the contrary, MRM pulp is less effective, and MA does not lead to statistically significant inhibition of TPA-induced colony formation.
These divergent results would suggest a synergistic effect between MA and other MRM callus secondary metabolites, which are not produced at the same level by MRM pulp. This result is in accordance with several epidemiological studies, 14 –16 suggesting that the whole phytocomplex MRM callus is involved in the resulting effects, representing the optimal environment for the produced metabolites to exert their biological activities.
To evaluate the molecular mechanisms involved in the ability of MRM callus to inhibit colony formation in soft agar cultures, we analyzed MAP kinase pathways, because of the positive correlation between p38, JNK, and ERK1/2 phosphorylation induced by TPA or Epidermal Growth Factor and colony formation in soft agar culture. 35 –39 Our results excluded ERK1/2 involvement, whereas an inhibition of p38 and JNK phosphorylation was exerted only by MRM callus in TPA-stimulated JB6 P+ cells, suggesting the involvement of p38 and JNK pathways in the ability of MRM callus to inhibit tumorigenic process in vitro. Moreover, the involvement of p38 was also considered through the analysis of a related protein, eIF2alpha; eIF2alpha is phosphorylated at a higher percentage in stomach, colon, and rectal tumors than in normal tissues. 40
eIF2alpha phosphorylation is an adaptive response to stresses and facilitates tumor cell survival and growth. 41 Zykova et al. showed the induction of eIF2alpha phosphorylation in stimulated JB6 P+ cells and demonstrated that p38 is required for this result. 42 Our results showed that phosphorylation of eIF2alpha is inhibited only in TPA-stimulated JB6 P+ cells treated with MRM callus, confirming the involvement of p38 in MRM callus efficacy. Notably, MRM callus and MA mechanisms of action are markedly different, underlying the importance of multitarget inhibitors; phytochemicals are known to inhibit several targets and their combination contained in the whole extracts provides synergistic or additive effects, increasing efficacy and chemopreventive potential. 43
Results from this study could clarify how MRM callus can modulate cancer initiation and proliferation, serving as a starting point for further research about the use of MRM callus in diet-derived antioxidant chemoprevention. Moreover, the prospect of using MRM callus in comparison with single compounds or the fruit itself must be considered; the opportunity to produce a variety of secondary metabolites from MRM apple in vitro cultures without depending on seasonal maturation or storage problems is highly advantageous.
However, further experiments would be necessary to assess the effect of the other compounds found in high amount in the MRM callus. Further studies will be conducted to evaluate whether the employed plant material could be used for the development of a nutraceutical product capable of contributing to the protection of people's health.
Footnotes
Acknowledgments
The authors would like to thank Dr. Elizabeth Tremmel, Dartmouth College, Hanover, NH, USA, for her linguistic revision of the article.
The authors are grateful to Dr. Andrea Carpi, (R&D Manager ABR) for kindly supplying tormentic acid used in this work.
Author Disclosure Statement
No competing financial interests exist.