
Abstract
Obesity is a worldwide epidemic, which is characterized by the excess accumulation of adipose tissue and to an extent that impairs both the physical and psychosocial health and well-being. There are several weight-loss strategies available, including dietary modification, pharmacotherapy, and bariatric surgery, but many are ineffective or not a long-term solution. Bioactive compounds present in medicinal plants and plant extracts, like polyphenols, constitute the oldest and most extensive form of alternative treatments for the prevention and management of obesity. Their consumption is currently increasing in the population due to the high cost, potential adverse effects, and limited benefits of the currently available pharmaceutical drugs. A great number of studies has explored how dietary polyphenols can interfere with the different mechanisms associated with obesity development. They suggest that these compounds can decrease energy and food intake, lipogenesis, and preadipocyte differentiation and proliferation, while increasing energy expenditure, lipolysis, and fat oxidation. Both quercetin, one of the most common dietary flavonols in the western diet, and epigallocatechin gallate (EGCG), the most abundant polyphenol in green tea, exhibit antiobesity effects in adipocyte cultures and animal models. However, the extrapolation of these potential benefits to obese humans remains unclear. Although quercetin supplementation does not seem to exert any beneficial effects on body weight, this polyphenol could prevent the obesity-associated mortality by reducing cardiovascular disease risk. An important consideration for the design of further trials is the occurrence of gene polymorphisms in key enzymes involved in flavanol metabolism, which determines a subject's sensitivity to catechins and seems, therefore, crucial for the success of the antiobesity intervention. Although the evidence supporting antiobesity effects is more consistent in EGCG than with quercetin studies, they could still be beneficial by reducing the cardiovascular risk of obese subjects, rather than inducing body weight loss.
Introduction
Currently obesity is one of the major public health problems; it is a worldwide epidemic with >35% of adults today considered to be overweight or obese. 1 A subject's body mass index (BMI, the weight in kilograms divided by the square of the height in meters) of 30 or higher is classified as obese. 2 Obesity is caused by an imbalance between energy intake and expenditure that results in adipose tissue dysfunction with adipocyte hypertrophy. 3 The main treatment options with sufficient evidence-based support are lifestyle intervention, pharmacotherapy, and bariatric surgery (Fig. 1). However, the lifestyle intervention shows low adherence and high rate of weight regain; pharmacotherapy may lose effectiveness in long-term treatments and bariatric surgery is invasive and expensive. 3
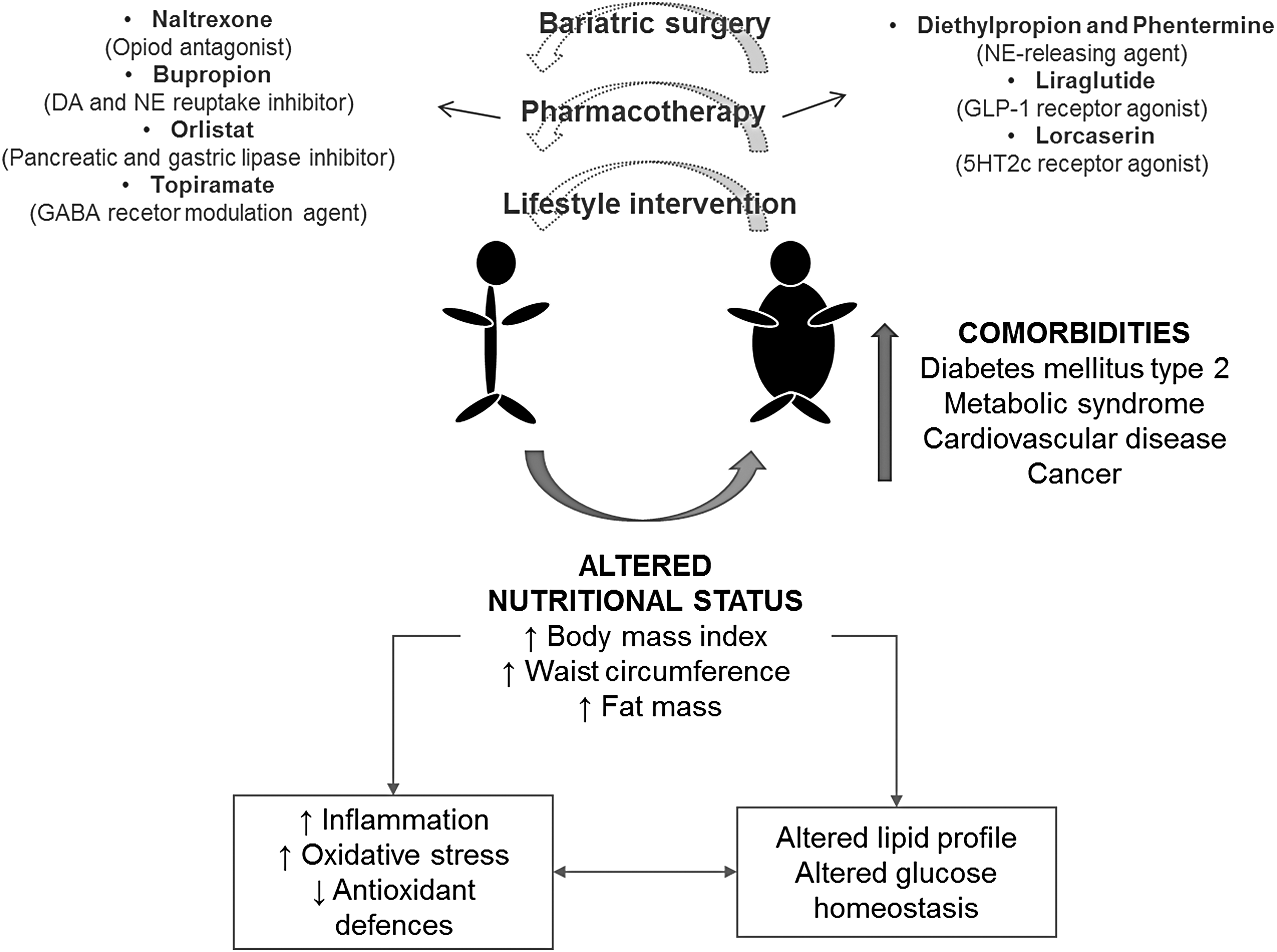
Obesity: physiopathological main features and treatment strategies. It is a low-grade inflammation disease with a high oxidative level. Treatments are scarcely effective as the first strategy to lose weight is focused on the modification of eating behaviors and physical activity. Obesity is associated with an increased risk of disease and death. DA, dopamine; GABA, γ-aminobutyric acid; GLP-1, glucagon-like peptide-1; NE, norepinephrine.
Obesity is comprised of several metabolic alterations accompanied by a state of chronic low-grade inflammation and an increased oxidative status that contributes to the development of health complications. Consequently, the intake of natural antioxidant and anti-inflammatory phenolic compounds seems to be a promising strategy for the prevention or management of obesity and its complications. Polyphenols constitute a ubiquitous group of secondary metabolites widely found in fruits, vegetables, cereals, legumes, nuts, and vegetable-derived foodstuffs, including wine, cocoa, coffee, tea, etc. Polyphenols are generally involved in plant defense against different types of pathogenic attacks and environmental stresses.
Epidemiological studies suggest that long-term consumption of polyphenol-rich diets is associated with lower risk of cardiovascular diseases, diabetes, osteoporosis, neurodegenerative diseases, and some type of cancers. 4 As there is an urgent need to find simple, effective strategies for improving weight loss, this review addresses the antiobesity effects of bioactive compounds that can be easily obtained through the diet, such as the polyphenols, quercetin and epigallocatechin gallate (EGCG).
Quercetin
Quercetin is ubiquitously present in fruits and vegetables, being one of the most common dietary flavonols in the western diet. According to the United States Department of Health and Human Services, the average daily intake of quercetin in humans is about 25 mg, 5 a value which is also supported by French and Finnish studies. 6,7 Onions are among the foods particularly high in this flavonol, bringing 0.03–0.28 mg/100 g of fresh weight (FW) with white and yellow, and red onion varieties exhibiting the highest content (ranging 1.31 mg/100 g FW). 8 –10 Quercetin has been reported to exert a wide range of biological effects, including anticarcinogenic, anti-inflammatory, and antiviral activities. In addition, this flavonol was shown to inhibit lipid peroxidation, platelet aggregation, and protect against mitochondrial dysfunction. 11,12 As a result, several studies also indicate that quercetin reduces health risks associated with obesity, such as heart disease, diabetes, and high blood pressure. 13 –15 However, its direct effect on the development and progression of obesity is still unclear.
Impact of quercetin on adipose tissue
The growth of adipose tissue, a hallmark of obesity, is due to the formation of new adipocytes from undifferentiated precursor cells (preadipocytes), further leading to an increase in adipocyte size (mature adipocytes) and eventually hypertrophy. 16 All treatments that modulate both size and number of adipocytes represent a promising approach for obesity prevention and management. Studies evaluating the growth of 3T3-L1 preadipocytes exposed to 100 μM quercetin show a highest inhibitory activity (71.5%) of this compound, compared with other polyphenols such as naringenin, naringin, hesperidin, rutin, and resveratrol. 17 This effect is probably due to its antioxidant properties, since quercetin exhibited the most potent antioxidant oxygen radical absorbance capacity of all the tested compounds. 17
Quercetin also inhibits the differentiation of preadipocytes to adipocytes by modulating a network of transcription factors and adipogenesis-related enzymes. Quercetin (10–100 μM) exerts anti-adipogenic activity in 3T3-L1 preadipocytes by activating the adenosine 5′ monophosphate-activated protein kinase (AMPK) pathway, specifically upregulating the levels of phosphorylated AMPK and its substrate, acetyl-CoA carboxylase (ACC). 18 Quercetin averted adipogenesis in 3T3-L1 preadipocytes, dose dependently (3.3–13 μM) decreasing the intracellular triglyceride content and glycerol 3-phosphate dehydrogenase activity (GPDH). 19 As this enzyme participates in lipid biosynthesis, the inhibition of GPDH activity decreases preadipocyte differentiation and lipid accumulation in mature adipocytes.
Two key modulators of adipocyte differentiation, peroxisome proliferator activator receptor γ (PPARγ) and CCAAT/enhancer-binding protein α (C/EBPα), promote the expression of adipogenic and lipogenic genes, including ACC that converts acetyl-CoA to malonyl-CoA, a fundamental intermediate component in fatty acid (FA) synthesis and an inhibitor of their oxidation, and the transcriptional factor sterol regulatory element-binding protein 1c (SREBP-1c). 20 In 3T3-L1, quercetin (3.3–13 μM) has been shown to decrease the mRNA level of PPARγ and C/EBPα and other FA anabolic genes highly expressed at the end stage of adipogenesis, like those coding for lipoprotein lipase (LPL) and adipocyte FA-binding protein (aP2). 19 LPL plays an important role in fat accumulation by hydrolyzing triglycerides (TG) from TG-rich lipoproteins, releasing free FA that pass through the endothelial wall and are uptaken by the adipocytes. 21 aP2 acts as a metabolism-regulating adipokine that plays a central role in the pathway linking obesity to insulin resistance and FA metabolism. 22
The effects of quercetin on lipolysis have also been studied in the mouse OP9 stromal cells, which rapidly differentiate into adipocytes. 23 In this model, 10 μM quercetin prevents adipogenesis by downregulating FA synthase (FAS), LPL, and aP2 expression and upregulating the expression of adipose triglyceride lipase (ATGL) and hormone-sensitive lipase (HSL). These latter enzymes play a role in lipolysis by catabolizing stored TG in lipid droplets; TGs are first hydrolyzed by ATGL into diglycerides, and ultimately in monoglycerides by HSL. 24
The effect of quercetin on TG accumulation varies according the to maturation state of the adipocyte and the concentration of this flavonol. At high concentration (10 μM), it reduces fat accumulation and LPL and FAS gene expression in mature adipocytes, whereas at a low concentration (∼1 μM), it decreases TG content as well as C/EBPβ and PPARγ gene expression in differentiating preadipocyte. 25 In addition, quercetin at high concentration (10 μM) has been also shown to inhibit osteoblastic differentiation and promote mesenchymal stem-cell-induced adipocyte differentiation. 26 Interestingly, quercetin 3-O-glucoside at 20 μM exerted a higher reduction of TG contents and lipid accumulation than quercetin, as well as a more significant decrease in the expression of the adipogenic markers, C/EBP-β, C/EBP-α, PPARγ, and aP2 in 3T3-L1 cells. 27 This indicates that glycosylation can affect the physicochemical and pharmacological properties of the flavonol. 27
Recently, quercetin (1 μM) was shown to be one of the more active molecule (among 15 various phenolic compounds) in reducing lipid accumulation in 3T3-L1 preadipocytes. 28 These findings indicate that the structure, concentration, and metabolism of the polyphenols as well as the model in which they are studied should be taken into account to determine their healthy properties and possible adverse effects, more particularly when their use as nutraceutical or pharmaceutical is being considered.
Another strategy to reduce the number of adipocytes and prevent preadipocyte differentiation is through apoptosis induction. In this aspect, 100 μM quercetin has been shown to stimulate apoptosis in 3T3-L1 cells by decreasing mitochondria membrane potential and the protein levels of poly (ADP-ribose) polymerase (PARP) and B cell lymphoma 2 (Bcl-2), and activated caspase 3, Bcl-2-associated X protein (Bax), and Bcl-2 homologous antagonist/killer (Bak). 17 Moreover, 3T3-L1 treatment with quercetin (10–100 μM) resulted in apoptosis induction and the concomitant decrease in extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinases (JNK) phosphorylation. 18 Noteworthy, it has been observed that JNK activation is associated with obesity and insulin resistance. 29,30 Recently, a quercetin-rich ethanolic extract prepared from purple corn silk was shown to decrease lipid accumulation and preadipocyte proliferation, in addition to inducing lipolysis and apoptosis in adipocytes. 31 In vitro effects of quercetin are summarized in Table 1.
Antiobesity Effects of Quercetin or Quercetin-Rich Extracts: In Vitro Studies
ACC, acetyl-coenzyme A carboxylase; AMPK, 5′ adenosine monophosphate-activated protein kinase; aP2, adipocyte protein 2; ATGL, adipose triglyceride lipase; Bak; Bcl-2 homologous antagonist/killer; Bax, Bcl-2-associated X protein; Bcl-2, B-cell lymphoma 2; C/EBPα, CCAAT/enhancer-binding protein alpha; C/EBPβ, C/EBP beta; ERK, extracellular-signal-regulated protein kinase; FABP4, fatty acid-binding protein 4; FAS, fatty acid synthase; GPDH, glycerol-3-phosphate dehydrogenase; HSL, hormone-sensitive lipase; JNK, c-Jun N-terminal kinase; LPL, lipoprotein lipase; PARP, poly (ADP-ribose) polymerase; PPARγ, peroxisome proliferator activator receptor gamma; SREBP-1c, sterol regulatory element-binding protein 1c; TG, triglycerides.
In addition to its effect on adipocyte cultures, quercetin markedly diminished transcript levels of inflammatory receptors and activation of their signaling molecules (ERK, p38 mitogen-activated protein kinase (MAPK), and nuclear factor (NF)-B) in cocultured myotubes/macrophages, and this was accompanied by reduced expression of the atrophic factors. 32 Indicating that quercetin may have a role in the prevention of obesity-induced skeletal muscle atrophy. 32
Impact of quercetin in animal models
Adipocyte lipogenesis and thermogenesis
Other mechanisms have been proposed for treating obesity, including the decrease of lipogenesis or the increase of lipolysis. For example, studies in rat adipocytes indicate that quercetin inhibits insulin-mediated lipogenesis by inhibiting the insulin receptor (IR) tyrosine kinase. 33 In mice fed a Western diet, quercetin (0.05% for 20 weeks) reduced visceral and liver fat accumulation, and PPARγ and SREBP-1c expression. It also improved hyperglycemia, hyperinsulinemia, and dyslipidemia. 34 Quercetin was also shown to modulate lipogenesis at the transcriptional level, preventing high-fat diet (HFD)-induced obesity and regulate hepatic expression of lipid metabolism-related genes, including Fnta, Pon1, PPARγ, Aldh1b1, Apoa4, Abcg5, Gpam, Acaca, Cd36, Fdft1, and Fasn. 35
Quercetin supplementation on mice fed a HFD significantly reduced their body weight gain, and prevented increased weight of the liver and white adipose tissue, compared with the unsupplemented animals. In fact, the hepatic fat accumulation and the size of lipid droplets in the epididymal fat pads were also reduced by the supplementation. Collectively, these results indicate that quercetin prevents HFD-induced obesity in C57B1/6 mice, being its antiobesity effects related to the regulation of lipogenesis at the transcriptional level. 35 In another study in HFD-fed rats, the increase in retroperitoneal and mesenteric fat weight was significantly reduced by supplementing the animal diet with 0.72% quercetin-rich onion peel extract. Concomitantly, the mRNA level of PPARγ and C/EBPα were downregulated and that of uncoupling protein-1 (UCP-1) was upregulated in the epididymal fat. 36
Low-grade inflammatory state
Obesity is associated with a chronic, low-grade inflammatory state mainly originated from the adipose tissue through the presence of activated macrophages, the increased secretion of proinflammatory adipokines, and the subsequent development of systemic inflammation, insulin resistance, and obesity-related metabolic complications. 37 Accordingly, preventing or decreasing this low-grade inflammation could be an interesting strategy to prevent obesity progression and development of comorbidities. 38 Taking these antecedents into account, the administration of quercetin (2 or 10 mg/kg of body weight for 10 weeks) reduced body weight gain in obese Zucker rats and exerted an anti-inflammatory effect in the visceral adipose tissue, increasing plasma adiponectin and reducing the plasma levels of nitrate plus nitrite and tumor necrosis factor (TNFα). 39 A reduction of the systolic blood pressure and plasma concentrations of TG, total cholesterol, free FA, and insulin were also reported in these animals. 39
Recently, the administration of quercetin (0.1% for 12 weeks) to mice fed a HFD was also shown to alleviate macrophage infiltration in the adipose tissue and modify their M1/M2 phenotype ratio, in addition to reduce the levels of proinflammatory cytokines through the activation of the AMPKα1/SIRT1 pathway. Glucose transporter type 4 (GLUT4) translocation and protein kinase B signaling were enhanced in the epidydimal adipose tissue of the animals. These events were accompanied by a decreased weight gain, loss of insulin sensitivity, and glucose intolerance. 40 In a similar study, quercetin (25 mg/kg bw/day) did not exert any effect on the plasma levels of cytokines, but downregulated the expression of genes involved in the immune and inflammatory response in the visceral and subcutaneous adipose tissue. 41 These data indicate that the anti-inflammatory effect of quercetin also affects target tissues of interest for obesity development.
On the other hand, quercetin was also reported to reduce obesity-induced hepatic inflammation by promoting macrophage phenotype switching. A 9-week administration of quercetin (0.5 g/kg diet) to HFD-fed mice decreased M1 marker genes (TNFα, NOS2) and increased M2 marker genes (Arg-1, Mrc1) in macrophages, resulting in lower hepatic concentrations of TNFα and interleukin (IL)-6 and higher of IL-10, an anti-inflammatory cytokine. 42 These results are supported by an in vitro coculture study using lipid-laden hepatocytes and macrophages exposed to quercetin. This induced the macrophage expression of heme oxygenase-1 (HO-1), suppressed the expression of M1 macrophage marker genes, and reduced the release of monocyte chemoattractant protein (MCP)-1. Consistently, these effects were blunted by using an HO-1 inhibitor and in macrophages deficient in nuclear factor E2-related factor 2 (Nrf2). 42 These results indicate that quercetin may act as a useful dietary factor protecting against obesity-induced steatohepatitis through Nrf2-mediated HO-1 induction.
In another study in mice fed HFD containing quercetin (0.2% w/w) for 10 weeks, the animals became obese but had significantly lower glycemia after food deprivation, lower concentrations of serum C-reactive protein, and decreased hepatic lipid accumulation. 43 The anti-inflammatory effect of quercetin has also been proposed as one of the main mechanism of action in the reduction of obesity-induced skeletal muscle atrophy, as it reduced the levels of inflammatory cytokines and macrophage accumulation in the skeletal muscle in obese mice fed a HFD for 9 weeks. 32 It also reduced the expression of the specific atrophic factors, Atrogin-1 and Muscle RING-finger protein-1 (MuRF1), in the skeletal muscle of the obese animals, protecting them against the reduction of muscle mass and muscle fiber size. 32
Taken together, these findings suggest that quercetin exerts anti-inflammatory effect against HFD-induced inflammation in several tissues, but only at high doses, as supplementation for 16 weeks with 0.02% quercetin was ineffective for attenuating the development of HFD-induced obesity and adipose tissue inflammation, nor in preventing the development of insulin resistance and hypercholesterolemia. 44 In vivo effects of quercetin in animal models are summarized in Table 2.
Antiobesity Effects of Quercetin: Animal Studies
Abcg5, ATP-binding cassette subfamily G member 5; AKT, serine/threonine-protein kinase; Aldh1b1, aldehyde dehydrogenase 1 family member B1; Apoa4, apolipoprotein A-IV; Atf2, activating transcription factor-2; Arg-1, arginase I; Cat, catalase; CD36, cluster of differentiation 36; Cpt1a; Carnitine palmitoyltransferase IA; CRP, C reactive protein; Fdft1, squalene synthase; Glut4, glucose transporter type 4; Gpam, glycerol-3-phosphate acyltransferase 1; Gpx1, glutathione peroxidase 1; GRO, human growth-regulated oncogene; GSH, reduced glutathione; IFNγ, interferon gamma; IL-1β, interleukin 1 beta; IL-6, interleukin 6; IL-10, interleukin 10; KC, keratinocyte chemoattractant; LDL, low-density lipoprotein; Mcp-1, monocyte chemoattractant protein-1; Mrc1, mannose receptor C-type 1; MuRF1, Muscle RING-finger protein-1; NOS2, nitric oxide synthase 2; pAKT, phospho AKT; pAMPK, phospho AMPK; Pon1, paraoxonase 1; PPARα, PPAR alpha; Scd1, stearoyl-CoA desaturase-1; Sirt1, sirtuin 1; TNFα, tumor necrosis factor alpha; TLR2, Toll-like receptor 2; TLR4, Toll-like receptor 4; UCP-1, uncoupling protein 1; VLDL, very LDL.
Impact of quercetin in overweight and obese subjects
Although the beneficial effects of quercetin in obesity have been widely studied in cell culture and animal models, there are only a few studies focused on investigating the antiobesity effects of quercetin in humans. So far, the health benefits of quercetin in overweight and obese subjects are unclear. Quercetin supplementation (100 mg/day/subject for 12 weeks) significantly decreased the total body fat, particularly in the percentage of fat in the arm, and decreased the BMI of overweight or obese subjects. 45 However, when the supplementation dose was increased (500 or 1000 mg/day combined with vitamin C and niacin for 12 weeks), the protective effect of quercetin on BMI or composition was no longer observed. 46
Another study in a BMI of overweight or obese subjects: showed that the administration of this polyphenol either pure (150 mg/day for 6 weeks) or as quercetin-rich onion peel extract (100 mg/day for 12 weeks) failed in improving their nutritional status (body weight, waist circumference (WC), fat mass, fat-free mass), serum lipid profile, and plasma TNFα and C reactive protein (CRP). 47,48 Moreover, a 12-week quercetin consumption (500 or 1000 mg/day) had no effect on oxidative stress and antioxidant capacity in obese subjects. 49 However, it reduced their systolic blood pressure and their plasma concentrations of atherogenic oxidized low-density lipoprotein (LDL). 47 Quercetin (162 mg/day for 6 weeks) also lowered systolic blood pressure in subjects with stage I hypertension. 50 Consistently, although quercetin (150 mg/day for 6 weeks) did not improve the nutritional status and plasma CRP, it was found to decrease serum lipid, blood pressure, and plasma TNFα in overweight/obese volunteers according to their ApoE genotype. 51
A study evaluated the effects of quercetin on normal-weight subjects with various ApoE genotypes; quercetin (150 mg/day/subject) decreased WC, postprandial triacylglycerol concentration, as well as fasting and postprandial systolic blood pressure. 52 While Egert et al. 51 observed an improvement only in ApoE3 homozygous subjects, in Pfeuffer et al., quercetin has a positive effect on both ApoE3 and ApoE4. This indicates that the beneficial impact of quercetin in cardiovascular risk depends on the nutritional status and the ApoE genotype of the subjects. ApoE is a polymorphic protein with three common isoforms in humans, ApoE2, ApoE3, and ApoE4, that mediates the binding of lipoproteins or lipid complexes in the plasma or interstitial fluids to specific cell surface receptors. 53 ApoE3 is the wild type and most common isoform, whereas the ApoE4 polymorphism is present in 25% of the Caucasian population and is associated with a significantly higher rate of cardiovascular diseases. 54
Interestingly, a single dose of quercetin (56 mg) did not prevent the increase of systolic and diastolic blood pressure, serum triglycerides, total cholesterol, and insulin and the decrease of HDL cholesterol induced by a single test meal rich in saturated FAs and carbohydrates in overweight-to-obese, hypertensive, subjects. 55 Currently, quercetin is being evaluated on a phase II clinical trial, addressing the inhibition of intestinal glucose absorption in nonhypertensive obese subjects and obese diabetic subjects. 56 It is possible that the results of this study may contribute to clarify the antiobesity effect of quercetin in normotensive subjects as general nutritional status may be controlled during the trial. In addition, these results will be of high relevance due to the size of the sample under study, as 100 obese patients are considered to be enrolled. This study will be completed in 2025. 56 In vivo effects of quercetin in humans are summarized in Figure 2.

Effect of long-term administration of quercetin in overweight and obese subjects. Quercetin has no effect in improving the nutritional status of the subjects and its effect on inflammatory status is unclear. The healthy effect of quercetin in obesity seems to be related to its prevention on comorbidity occurrence, for example in reducing the risk of cardiovascular diseases. ALT, alanine transaminase; AST, aspartate transaminase; CRP, C-reactive protein; FRAP, ferric reducing ability of plasma; GSH, reduced glutathione; LDL, low-density lipoprotein; ORAC, oxygen radical absorbance capacity; QUE, quercetin; TNFα, tumor necrosis factor-alpha.
In conclusion, results from in vitro and animal studies suggest that quercetin beneficially impacts the adipose tissue by modulating the expression of genes involved in the regulation of adipocyte differentiation and maturation, fat accumulation, and basal metabolism. In addition, quercetin also decreases the inflammatory state of the adipose tissue by decreasing macrophage infiltration and activation and improving the release of proinflammatory adipokines and adiponectin (Tables1 and 2). Accordingly, quercetin would protect against obesity development as a result of the lower adipogenesis, lipogenesis, and inflammation, and the higher basal metabolism in the adipose tissue. Interestingly, this polyphenol also appears to prevent the hepatic and muscular complications associated with obesity in animal models. Unfortunately, the extrapolation of these results to obese humans remains unclear.
Besides its marginal effect on body weight, quercetin exerted only limited anti-inflammatory and antioxidant effects in obese subjects. This is particularly unfavorable for quercetin, considering that obesity is characterized by being a low-grade inflammation (and in consequence, a low-grade oxidation) disease. Although dietary supplementation with quercetin does not seem to exert beneficial effects in body weight, it could be an interesting strategy to prevent obesity-associated mortality by reducing the risk of cardiovascular disease (Fig. 2).
Epigallocatechin Gallate
EGCG is a flavonoid from the flavan-3-ol subclass, highly present in green tea; a cup of this beverage carries 200–300 mg of this compound. 8 –10 More than 800 published studies have shown the effects of green tea on cancer, mostly based on its chemopreventive effects. 57 It is generally agreed that such effects are mediated by catechins comprising of 50–80% EGCG. 58 EGCG has also demonstrated other beneficial effects in Parkinson's and Alzheimer's diseases, stroke, obesity, and diabetes type 2. 58 –62
Impact of ECGC on adipose tissue
Adipocyte differentiation and proliferation
The antiobesity effects of EGCG have been studied focusing on several potential mechanisms of action, including the inhibition of preadipocyte differentiation, adipocyte proliferation and lipid accumulation, and the induction of thermogenesis, among others. It has been proposed that EGCG downregulates the major genes of the adipogenesis pathway by upregulating β-catenin through WNT/β-catenin pathway activation. 63 EGCG has been shown to inhibit the proliferation of 3T3-L1 preadipocytes and their differentiation into mature adipocytes by undergoing an arrest of cell cycle. 64 –66 These events were due to the reduction of forkhead box protein O1 (FOXO1) activation 66 and to its antimitogenic activity, decreasing the levels of phospho-ERK1/2, cyclin-dependent kinase 2 (Cdk2), and cyclin D1 proteins and Cdk2 activity. 65,67,68 EGCG has also been shown to induce apoptosis of mature adipocytes 69 through Cdk2- and caspase-3-dependent mechanisms. 67
Interestingly, EGCG was more effective than epicatechin, epicatechin gallate, and epigallocatechin in modulating the mitogenic 65,70,71 and apoptotic signals. 65,67,69 The observed catechin-specific effects suggested that EGCG may act through a different mechanism than epicatechin, epicatechin gallate, and epigallocatechin in regulating preadipocyte growth. In addition, EGCG structure may be more favorable for molecular interactions, which may explain its greater efficacy. EGCG contains the largest number of hydroxyl groups on its three aromatic rings among the tea catechins, and these groups may be important for hydrogen bonding in molecular interactions. 72 Also, EGCG has both gallyl and galloyl groups, which confer conformational flexibility that may also be important for interactions with other molecules. Further investigations of the chemical basis of the antimitogenic and proapoptotic activity on preadipocytes by catechins are needed to understand the differences in their mechanisms and activities.
Recently, it has been suggested that the disturbance of membrane lipid rafts by A-type EGCG dimer was involved in its inhibitory effects on 3T3-L1 preadipocyte differentiation. 73 A-type EGCG dimer significantly reduced the intracellular lipid accumulation in 3T3-L1 preadipocyte cells by targeting miR-27a and miR-27b as well as PPARγ in the early stage of differentiation of 3T3-L1 preadipocyte. 74 This effect is associated with the dimer/membrane interactions and the presence of galloyl moieties and A-type linkage within the structure of proanthocyanidins, which might be crucial for their inhibitory effect on adipogenesis. 74
On the other hand, EGCG inhibited insulin and insulin-like growth factor I (IGF-I) stimulation of preadipocyte proliferation by decreasing the phosphorylation of IR-β, IR substrate (IRS)1 and IRS2, and the MAPK proteins, RAF1, MEK1/2, and ERK1/2. 70,71 However, in adipocytes EGCG suppressed the IGF-stimulated phosphorylation of IGF-signaling molecules, PKCζ/λ, but not that of AKT and ERK1/2 proteins. 75 It has been shown that EGCG suppresses IGF stimulation of 3T3-L1 adipocyte glucose uptake by inhibiting GLUT4 translocation without affecting the GLUT1 pathway. 75 Moreover, through the PI3K/Akt insulin signaling, EGCG also suppressed the clonal expansion of adipocytes by inactivating FoxO166 and inhibited adipogenesis of preadipocyte by downregulating PPARγ and FAS expression. 76
Adipocyte lipogenesis and thermogenesis
EGCG upregulates lipolysis and thermogenesis. For example, EGCG was found to reduce total TG accumulation in murine 3T3-L1 preadipocytes during their differentiation to adipocytes 66 and in maturing preadipocytes 69 by increasing glycerol release. 77 EGCG could affect adipocyte conversion and fat accumulation by decreasing the expression of C/EBPα, PPARγ, and SREBP-1c 63,64,66,74,76,78 –82 and by activating AMPK, a suppressor of PPARγ and C/EBPα expression. 64,80,83
Moreover, EGCG increased the mRNA levels of the UCP-2, a mitochondrial transporter present in the inner mitochondrial membrane, which induces heat production by uncoupling respiration from ATP synthesis. 84 Thus, UCP-2 is a crucial protein in thermogenesis that increases the electron flow through the mitochondrial electron transport chain, promoting the energy expenditure with a reduced metabolic efficiency. EGCG upregulates the expression of PPARγ coactivator (PGC-1α) and its promoter activity in differentiated 3T3-L1 adipocytes. PGC-1α is a crucial regulator of mitochondrial biogenesis that participates in the thermogenesis process. 85 EGCG also increases sirtuin 1 abundance in adipocytes, which appears to be involved in its ability to enhance glucose consumption and suppress p65 phosphorylation-mediated NF-κB activation, as the sirtuin 1 inhibitor nicotinamide attenuates those effects exerted by EGCG. 86
EGCG and caffeine, synergistically with norepinephrine stimulate the thermogenesis of brown adipose tissue, 87 by regulating various enzymes related to lipid anabolism and catabolism, such as ACC, FAS, pancreatic lipase, gastric lipase, HSL, and lipoxygenase. 77,88 –91
Recently, a novel mechanism underlying the effect of EGCG on the reduction of TG content in adipocytes has been proposed. It involves the induction of autophagic lipolysis by upregulating Beclin1 and ATG7 expression and promoting the association between RAB7 and lipid droplets, which suggest lipophagy activation. 92 Lipophagy is a form of autophagy specialized for the degradation of lipid droplets, and RAB7 is a key molecule that enables the specific recognition of lipid droplet proteins in this process. 93 In vitro effects of EGCG and green tea extracts (GTEs) are summarized in Table 3.
Antiobesity Effects of Epigallocatechin Gallate and Green Tea Extracts: In Vitro Studies
Akt, protein kinase B; Cdk 2, cyclin-dependent kinase 2; EGCG, epigallocatechin gallate; FOXO1, forkhead box protein O1; FOXO3, forkhead box protein O3; IR, insulin receptor; IRS, IR substrate; LXRα, liver X receptor alpha; MAPK, mitogen-activated protein kinase; PI3K, phosphoinositide-3-kinase; ROS, reactive oxygen species.
Impact of EGCG and GTEs in animal models
The antiobesity effects of EGCG and GTE have been widely confirmed in animal studies. EGCG or GTE have been reported to decrease body weight, adipose mass, 94 –111 total lipids, cholesterol, and triglycerides in liver and plasma, 79,94,96,101,104,105,108,112,113 and improve glucose homeostasis. 94,96,98,105,107 –109,112,114 Supporting the role of ECGC against obesity, the intraperitoneal injection of this flavanol, but not other structurally related catechins (epicatechin, epigallocatechin, or epicatechin gallate) reduces body weight, blood levels of testosterone, estradiol, leptin, insulin, IGF-I, glucose, cholesterol, and triglyceride in male and female Sprague-Dawley rats within 2–7 days of treatment. 88,115
In mice fed a HFD supplemented with EGCG (3.2 g/kg diet for 16 weeks), Bose et al. reported decreases of body weight gain, body fat percent, visceral fat weight, insulin resistance, plasma cholesterol, MCP and alanine aminotransferase concentrations, liver weight, and liver triglycerides. 96 Interestingly, short-term treatment with EGCG (4 weeks) reversed preexisting HFD-induced metabolic pathologies in obese mice, as the mesenteric fat weight and blood glucose were decreased. 96 Similar effects were observed with GTE (0%, 1%, or 2% (w/w) for 6 weeks) in obese male leptin receptor-deficient Zucker rats 115 and in male leptin-deficient (ob/ob) mice, 97 suggesting that the leptin pathway is not involved in the effect of EGCG.
Short-term supplementation (4–7 days) with EGCG (up to 1% [w/w]) in mice fed a HFD increased fecal energy excretion (increased fat and nitrogen excretion) and postprandial fat oxidation while macronutrient and energy intakes were not affected. 101 EGCG supplementation also decreased postprandial triglycerides, hepatic glycogen content, and the incorporation of dietary lipids into fat, liver, and skeletal muscle tissues. 101 The authors showed that EGCG dose dependently reversed the HFD-induced effects on the intestinal nutrient transporters CD36, FATP4, and SGLT1, and downregulated lipogenesis-related genes (ACC, FAS, and stearoyl-CoA desaturase-1 (SCD1)) in liver in the postprandial state. 101 This effect on nutrient digestibility was also observed with a highly purified extract from green tea leaves containing minimum 94% EGCG (TEAVIGO™; DSM Nutritional Products, Basel, Switzerland) administered for 4 weeks to HFD-fed mice. 102 The supplementation resulted in an attenuation of body fat accumulation; noteworthy food intake was not affected while fecal energy content was slightly increased by EGCG. This lower food digestibility was accompanied by the downregulation of SCD1 and upregulation of UCP2 gene expression in white fat and liver. 102
Recently, the role of EGCG in energy expenditure and bioenergetics has also been studied, and EGCG (50 mg/kg/day, 16 weeks) has been shown to prevent obesity in HFD-fed mice by stimulating the mitochondrial respiratory chain, increasing energy expenditure, particularly from fat oxidation, thereby contributing to the prevention of hepatic steatosis and improved insulin sensitivity. 116 A GT (0.5% w/w) extract (1000 mg GT powder containing 464 mg EGCG, for 4 months) prevented the decrease on energy expenditure induced by HFD, by preventing the decrease of Adcyap1r1 (adenylate cyclase-activating polypeptide 1 receptor 1) and Adrb1 (Adrenergic, beta-1-, receptor) gene expression. 110
On the other hand, EGCG administration (75 mg/kg, i.p., 3 times/week, 17 weeks) to HFD-fed mice decreased the levels of advanced glycation end products (AGEs) in both plasma and liver while inhibiting the expression of receptor for AGE, activating Nrf2, and enhancing GSH/GSSG ratio. These effects were also accompanied by a reduction in weight gain, plasma glucose, insulin level, and liver and kidney weight. 117
GT supplementation (4%, 22 weeks) reduced the body fat content and hepatic triacylglycerol and cholesterol accumulation in mice fed a HFD; this reduction was negatively correlated with the amount of Akkermansia muciniphila and the total amount of bacteria in the small intestine. The protective effect of GT may be mediated by the microbiota as GT in combination with a single strain of Lactobacillus plantarum was able to promote growth of Lactobacillus in the intestine, and attenuate HFD-induced inflammation. 95 Consistently, shorter supplementation with GT (400 mg/kg bw/day, 8 weeks) also attenuated low-grade inflammation in HFD-induced obese mice, which was evidenced by reduced TNFα levels as well as TLR4, MYD88, and TRAF6 proinflammatory signaling. 99
Green tea (up to 1%, 6 weeks) administered to obese ob/ob mice decreased inflammatory status by reducing hepatic TNFα protein and inhibited adipose TNFα mRNA level, and restored antioxidant defenses by normalizing total glutathione, malondialdehyde and Mn- and Cu/Zn superoxide dismutase activities and increasing catalase and glutathione peroxidase (GPX) activities in the liver. 104 This anti-inflammatory and antioxidant effect of GTE (0.5% for 4 weeks) was also found in HFD-fed obese female rats, in which serum IGF-1 and leptin as well as proinflammatory cytokines, and liver GPX protein level were decreased, compared with the unsupplemented animals.
In addition, GT benefited body composition (fat mass, fat-free mass, total body water, intracellular fluid and extracellular fluid) and bone properties (bone mineral density and strength) in obese rats, increasing percentage of fat-free mass and bone mineral density/strength. 106 In obese mice (ob/ob), GTE (1%, containing 30% total catechins (w/w), for 6 weeks) prevented the increase on hepatic lipid accumulation, inflammatory infiltrates and serum alanine aminotransferase activity. It has been proposed that GTE mitigates lipid peroxidation and protein nitration by suppressing the generation of reactive oxygen and nitrogen species. 118
The beneficial effect of GTE, regular exercise, and regular exercise plus GTE was compared in mice fed HFD. 107 After 15 weeks of treatment, GTE almost doubled the reduction in body weight gain and visceral fat accumulation compared with the regular exercise. Interestingly, when both treatments were combined, the reduction of these parameters was twice as important. GTE supplementation increased hepatic FA oxidation both in the exercised and nonexercised animals and, when combined with regular exercise, it also stimulated skeletal muscle FA oxidation. 107
On the other hand, the administration of GTE to lean rats was also shown to modulate glucose uptake by decreasing GLUT4 translocation in adipose tissue and skeletal muscle, and suppressing the expression and/or activation of the adipogenesis-related transcription factors, PPARγ and SREBP-1. 94
Total cholesterol in the liver, triglycerides in serum and liver, and nonesterified FAs in serum from mice, which were administered green tea diet (4% for 16 weeks), were lower than those in the controls. 111 Consistent with the improvement on the lipid profile, the administration of GTE (0.05%) to ob/ob mice fed a HFD for 12 weeks, increased plasma HDL cholesterol and reduced adipose tissue weights and hepatic triglyceride content. A suppression of the enzymes implicated in FA synthesis (glucose-6-phosphate dehydrogenase and malic enzyme) was also found in the GTE group, without changing those involved in FA oxidation in hepatic and adipose tissue. 100 In a similar study, the reduction in adipose tissue weight induced by GTE intake was also accompanied by an increase in the expression level of IGF-binding protein-1 (IGFBP1) in white adipose tissues. 109
In a comparative study conducted for 6 months with green tea (20 g Dilmah infusion) and EGCG (1 mg/kg/day) in HFD-fed rats, EGCG was shown to increase the expression of genes involved in thermogenesis and differentiation (UCP2 and PPARγ) in adipose tissue, but without effect on liver or muscle at this dose. 98 However, GT infusion suppressed adipocyte differentiation (Pref-1, C/EBP-beta, and PPARγ gene expression) and increased the liver expression of genes involved in FA synthesis (SREBP-1c, FAS, Malonyl-CoA decarboxylase (MCD), ACC) and oxidation (PPAR-α, Carnitine palmitoyltransferase (CPT-1), acetyl-CoA oxidase (ACO)), without inducing hepatic fat accumulation. 98 Conversely, when green tea was delivered as extract (up to 1%) for a shorter time (6 weeks) in genetically obese animals, a decrease in the hepatic steatosis markers, SREBP-1c and FAS, was observed, in addition to a decrease in SCD1 and HSL mRNA level and lower plasma concentrations of nonesterified FAs. 104
Recently, the neuroprotective property of EGCG has been studied in a model of obesity induced by high-fat and high-fructose diet in mice. 119 This compound (2 g/L for 16 weeks) attenuated neuronal damage, learning alterations, and memory loss induced by this diet, in addition to prevent body weight gain, fat accumulation and insulin resistance. It significantly ameliorated insulin resistance and cognitive disorders by upregulating the (IRS-1)/AKT and ERK/cAMP cAMP response element-binding protein/brain-derived neurotrophic factor signaling pathways. The neuroinflammation process was decreased by EGCG through the inhibition of MAPK and NF-κB pathways, and TNFα expression. Consistently, a key role has been suggested for EGCG in the regulation of feeding behavior and energy homeostasis, specifically by counteracting the daytime overeating induced by HFD in mice. 120 In vivo effects of EGCG and GTEs in animal models are summarized in Table 4.
Antiobesity Effects of Epigallocatechin Gallate and Green Tea Extracts: Animal Studies
ACO, acetyl-CoA oxidase; Adcyap1r1, adenylate cyclase activating polypeptide 1 receptor 1; Adrb1, adrenergic receptor, beta-1; Agrp, agouti-related protein; ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate transaminase; Cntfr, ciliary neurotrophic factor receptor; Ghr, growth hormone receptor; Ghrl, ghrelin/obestatin prepropeptide; GM-CSF, granulocyte/macrophage colony-stimulating factor; HDL, high-density lipoprotein; 4-HNE, 4-hydroxynonenal; iNOS, inducible nitric oxide synthase; Lepr, leptin receptor; LPO, lactoperoxidase; MCAD, medium-chain acyl coA dehydrogenase; MPO, myeloperoxidase; NADPH, nicotinamide adenine dinucleotide phosphate; NF-κB, nuclear factor-κB; Nr3c1, nuclear receptor subfamily 3, group C, member 1; NRF1, nuclear respiratory factor 1; SCD1, stearoyl-CoA desaturase-1; SOD, superoxide dismutase; Sort1, sortilin 1.
Impact of EGCG and GTEs in obese human subjects
Controversial results have been reported regarding the impact of GT, EGCG, or GTE, with or without caffeine, in overweight/obese subjects. Indeed, while several studies reported a beneficial impact of these products on body weight, BMI, fat mass, waist and hip circumference, abdominal fat, and intra-abdominal adipose tissue, 121 –125 others did not observe any effect. 126 –128
In a randomized, double-blind trial conducted in 115 women with BMI ≥27 kg/m2 and WC ≥80 cm, the administration of high dose of GTE (857 mg of EGCG thrice a day for 12 weeks) significantly decreased BMI and WC, in addition to lower ghrelin and increased adiponectin levels. 125 Consistently, the administration of GT (690 mg/day) for 12 months improved the nutritional status and reduced oxidative stress in normal and overweight males. 129 The administration of catechin (∼580 mg catechins/day) for 24 weeks to obese children 130 or 12 weeks to obese adult 131 promoted WC reduction and decrease in LDL levels, 130,131 but had no effect on inflammatory biomarkers and leptin levels. 130
Green tea beverage consumption (4 cups/day) for 8 weeks significantly decreased BMI and biomarkers of lipid peroxidation in subjects with obesity and metabolic syndrome. 122 The administration of GT (250 mg containing mostly caffeine and EGCG, three times per day) to obese subjects for 8 weeks reduced their body weight, WC, and respiratory quotient (RQ), and increased their energy expenditure, without effect in their satiety score, food intake, or physical activity. 121 A similar effect was found when green tea (containing 104 mg caffeine/day, 573 mg catechins/day) was administered to overweight and obese subjects for 17 weeks. 132 This effect on RQ was also reported after administering 300 mg of EGCG/day for only 2 days in overweight men. 123
Interestingly, the protection against body weight gain was also evidenced after administering decaffeinated GTE (∼400 mg catechins twice daily for 6 weeks). 124 Moreover, in healthy men the administration of caffeine (50 mg, three times per day) did not affect 24-h energy expenditure and RQ, whereas GTE (50 mg caffeine + 90 mg EGCG, twice daily) improved these parameters. 133 Taken together, these findings indicate that EGCG alone increase fat oxidation and might thereby contribute to the antiobesity effects of green tea, independently of the caffeine content.
On the other hand, although EGCG administration (300 mg/day for 12 weeks) to overweight/obese postmenopausal women with moderate physical activity did not further increase their losses in WC, total body fat, abdominal fat, and intra-abdominal adipose tissue, it significantly decreased resting heart rate and reduced glycemia in those with impaired glucose tolerance. 127 On the contrary, GTE administration to overweight/obese women (375 mg catechins containing 270 mg EGCG daily for 12 weeks 126 or 1125 mg tea catechins +225 mg caffeine/day for 87 days 128 ), did not affect their BMI, WC, or energy expenditure. 126,128 In overweight and obese adults the administration of GTC (625 mg GTC, 39 mg caffeine/day for 12 weeks) promoted body weight reduction and improved the lipid profile, but had no effect on oxidative stress and inflammation biomarkers and on glucose homeostasis. 134
Some studies have reported a protective effect of GTE on cardiovascular risk factors in obesity, by decreasing both systolic and diastolic blood pressure, 135,136 improving lipid profile (reducing total cholesterol, LDL, and TG and increasing HDL) 136 and reducing plasma serum amyloid-alpha, an independent cardiovascular disease risk factor. 137 However, other studies did not report any beneficial effects of GT on these factors in obese subjects. 122,124 While GTE was also found to reduce fasting serum glucose and insulin and insulin resistance, 136 other studies have shown that GT failed in improving these parameters. 135 Regarding the protective effect of GTE against chronic low-grade inflammation, its supplementation for 3 months decreased serum TNFα and CRP. 136 However, other studies did not support these results. GT beverage or GTE supplementation for 8 weeks did not significantly improve the features of metabolic syndrome or inflammation biomarkers, including adiponectin, CRP, IL-6, IL-1β, soluble vascular cell adhesion molecule-1, soluble intercellular adhesion molecule-1 (sICAM-1), and the leptin:adiponectin ratio. 137
These controversial findings about the healthy properties of dietary flavonoids could be due to genetic polymorphisms in the genes codifying for the key enzymes involved in the absorption and metabolism of flavonoids, resulting in high interindividual variability in these processes. O-methylation, which is an important pathway of flavonoid metabolism, especially for catechins, is catalyzed by the catechol-O-methyltransferase (COMT). For example, a genetic polymorphism (G → A substitution at codon 108/158 resulting in Val→Met change in the COMT protein) has been identified, which alters the function of this enzyme, reducing its activity by three to fourfold. 138,139 It is therefore possible that this polymorphism influences the rate of catechin metabolism and modulates the systemic exposure to methylated derivatives. In fact, in the study by Brown et al., COMT Val/Met genotype was found to influence the 24-h urinary accumulation of EGC and 4′-O-methyl EGC. Lower concentrations were noted for homozygous individuals with the high-activity H-allele compared with those carrying at least one copy of the low-activity L-allele. These authors suggest that this may reflect differences in metabolic fluxes, with the conversion of tea catechins to downstream metabolites occurring more rapidly in the individuals homozygous for the G-allele. 124
Asian populations appear to have a higher frequency of H-allele (Val/Val polymorphism) than the Caucasian populations, who have a higher frequency of L-allele. Half of the Caucasian population is homozygous for the COMT L-allele (25%) and the COMT H-allele (25%), being the remaining 50% heterozygous. This genotype might explain the difference in sensitivity to catechins and the failures in some studies conducted in Caucasian subjects, as these differences in COMT activity are thought to affect individual variation in metabolism of green tea catechins, thus potentially influencing the biological effects of green tea consumption on weight loss and weight control. For example, the administration of decaffeinated GTE (843 mg EGCG/day for 12 months) to overweight and obese postmenopausal women with high-COMT activity showed a higher postmeal insulin response than those with low-COMT activity. However, GTE had no effect on plasma leptin, ghrelin, and adiponectin, independently of the COMT genotype. 140 In vivo effects of EGCG and GTEs in humans are summarized in Table 5.
Antiobesity Effects of Epigallocatechin Gallate and Green Tea Extracts: Human Studies
BMI, body mass index; DBP, diastolic blood pressure; GTC, green tea catechin; GTE, green tea extract; HbA1C, glycated hemoglobin; HOMA-IR, homeostatic model assessment-insulin resistance; REF, reference; RQ, respiratory quotient; SBP, systolic blood pressures; sICAM-I, soluble intercellular adhesion molecule I; sVCAM-1, soluble vascular cell adhesion molecule-1; WC, waist circumference.
Taking into account the in vitro effects of EGCG on fat cell functions and adipocyte differentiation and its effects on body weight, body fat, thermogenesis, and fat oxidation, it may be concluded that the intake of EGCG, mainly under the form of GT, contribute to decrease the incidence of obesity and could be useful for its treatment. However, it is important to consider the subject's sensitivity to catechins in the design of further trial, given that the occurrence of polymorphism in key enzymes involved in flavonol metabolism seems to be crucial for treatment success.
Conclusion
Although, both the polyphenols reviewed in this study have shown antiobesity effects in in vitro and animal models, there are more studies on EGCG than on quercetin addressing their role in obesity. They have shown to reduce adipocyte mass by inhibiting the proliferation and/or by promoting the apoptosis of adipocytes. They have also demonstrated to decrease adipogenesis and lipid accumulation interfering with several enzymes and markers involved in lipid formation and breakdown in adipocytes. These antiobesity effects found in vitro may give insights of the mechanism underlying their antiobesity effects observed in animal models, in which quercetin and EGCG induced the reduction of body weight and fat mass. They have consistently, been shown to decrease the inflammatory state of obese animals and ultimately contribute to improve glucose homeostasis and lipid profile.
Their antiobesity effects in humans, however, are still unclear. There are several studies addressing the effect of EGCG in the nutritional status, and most of them have shown a decrease on BMI, WC, fat mass, and inflammatory status. Quercetin, on the other hand, has been less studied than EGCG in human subjects, and the evidence found in nutritional status improvement and inflammation reduction is weaker than that found with EGCG (Fig. 3). As quercetin is absorbed 10 times more efficiently than EGCG in humans 141 –143 and both polyphenols have a half life ∼3.5 h, 141,143 it is unlikely that their differences in antiobesity effects can be explained by their pharmacokinetic profiles.
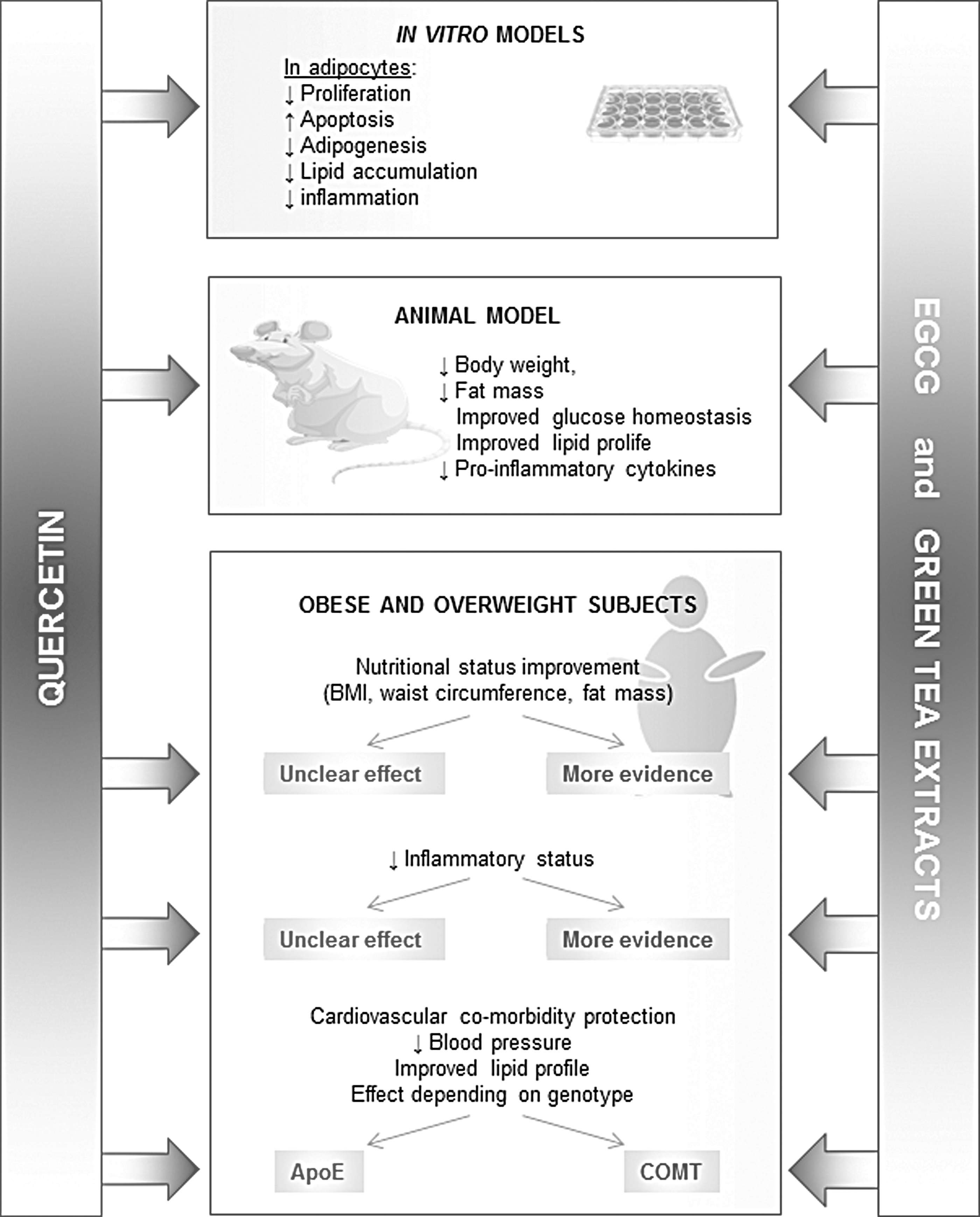
Comparison of the antiobesity effects of quercetin and epigallocatechin gallate. Most documented findings are shown in in vitro studies and animal and human models.
Compared with quercetin, a larger amount of unabsorbed EGCG can reach the colon and be metabolized by the microbiota. These metabolites may contribute to the antiobesity effects found with the original unmetabolized, absorbed compound. Microbial enzymes promotes the hydrolysis of the ester bonds of EGCG and the fission of the C-ring of catechins to produce gallic acid and ring-fission metabolites. 144 Although the antiobesity effects of gallic acid (or gallic acid-rich extracts) have been reported in in vitro, animals and human models, 145,146 more studies are needed to understand the contribution of the microbiota metabolites to the antiobesity effects exhibited by the original compounds. Obesity is associated with increased mortality due to cardiovascular disease. 147 Even though quercetin and EGCG do not have a relevant impact on body weight loss, they may contribute to save the life of obese and overweight subjects as they can reduce the cardiovascular risk by decreasing blood pressure and improving lipid profile.
Footnotes
Acknowledgments
The present study was supported by Proyecto de Investigación SOCHINUT/Nestlé 2016 and Fondecyt Initiation into Research Grant 11130232 to C.C.-P.
Author Disclosure Statement
No competing financial interests exist.