
Abstract
The mechanisms underlying the effect of Ganoderma lucidum (Reishi mushroom) polysaccharides (GLP) on obesity are not clear. In this study, GLP were found to attenuate the oleic acid–induced cell viability loss and apoptosis dose dependently in splenic lymphocytes in vitro. The effects of GLP on lipid metabolism, oxidative stress, and apoptosis in mice fed a high-fat diet (HD) were determined. GLP administration (200 and 400 mg/kg bw) significantly lowered the body-weight increases; liver, heart, and white adipose tissues indexes; serum lipid accumulation; and serum and small intestine oxidative stress in mice fed a HD. Moreover, GLP inhibited HD-induced apoptosis by decreasing the Bax/Bcl-2 ratio and suppressing caspase-3 activation in splenic lymphocytes. These findings indicate that GLP can exert hypolipidemic, antioxidant, and antiapoptotic effects in HD-induced obese mice.
Introduction
O
Pharmacotherapy, diet modification, intragastric balloon treatment, behavior modification, and bariatric surgery are essential for both the prevention and management of obesity. 7 When behavioral approaches are insufficient, pharmacological approaches may be recommended for treating obesity. However, the success rate of anti-obesity treatments is very low. Once pharmacological approaches are discontinued, weight is often regained. 8 Most of the anti-obesity drugs (e.g., amphetamine, orlistat, and sibutramine) were approved for use in Europe and the United States, but they have since been withdrawn from the market due to their adverse event profiles, which include insomnia, constipation, restlessness, increased blood pressure, hepatic damage, and mild to moderate gastrointestinal disease. 9,10
Many studies have proven that obesity increases oxidative stress, which is a potential pathogenic mechanism of obesity-associated metabolic syndrome. 11 Oxidative stress has been implicated in pancreatic β-cell dysfunction and vascular complications in diabetes. Remarkably, obese people and mice with or without diabetes develop intensified systemic oxidative stress in their adipose tissue. 12 Therefore, antioxidants are used as protective and preventive agents to relieve the body weight gain–associated oxidative inflammatory status and to treat various obesity-induced diseases. 13,14
Recently, due to their low adverse effects, biologically active ingredients extracted from functional food have attracted attention for their prevention and treatment of obesity and obesity-related disorders. 15,16 Reishi mushroom, Ganoderma lucidum, is a basidiomycete belonging to the Polyporaeae family that has been used as a panacea in Eastern countries for thousands of years. Polysaccharides derived from G. lucidum possess antioxidative, anticancer, anti-inflammatory, and immunopotentiating properties. 17,18 These polysaccharides could be remarkably effective for alleviating many diseases, with no side effects or toxicity. 19 –21 However, studies evaluating the potential of G. lucidum (Reishi mushroom) polysaccharides (GLP) to treat high-fat diet (HD)-induced obesity and oxidative stress remain scarce. Therefore, this study was designed to investigate the protective effects of GLP against lipid peroxidation, oxidative stress, and apoptosis in HD-fed mice.
Materials and Methods
Materials
G. lucidum (Leysser: Fr) Karst was obtained from Erli Biotechnology Co. Ltd. (Changsha, China; cat. no. 20160412). Cell counting kit-8 (CCK-8) was obtained from Dojindo (Tokyo, Japan). Percoll was purchased from GE Healthcare (Waukesha, WI, USA). Alpha-lipoic acid (LA) was obtained from Sigma–Aldrich (St. Louis, MO, USA). Low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), total cholesterol (TC), total triglycerides (TGs), malondialdehyde (MDA), catalase (CAT), and glutathione peroxidase (GSH-Px) levels were measured by kits from Nanjing Jiancheng Bioengineering Institute (Nanjing, China) according to the manufacturer's instructions. A Total Superoxide Dismutase Assay Kit was obtained from Beyotime Biotechnology (Nantong, China). An Annexin V-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit was obtained from Santa Cruz Biotechnology (Dallas, TX, USA).
Preparation of GLP
GLP was isolated from the fruit bodies of G. lucidum, as previously described. 22 The crude extract was composed of acidic polysaccharides containing 89% total carbohydrate and 11% uronic acid. The molecular weight composition of the polysaccharide was as follows: 10–30 kDa, 32.1%; 30–50 kDa, 21.8%; >50 kDa, 46.1%. The polysaccharides consisted of arabinose, galactose, cellulose, and glucose at a ratio of 11:3:1:3, and α-glycosidic linkages were present. GLP powder was dissolved in RPMI 1640 media with 2% fetal calf serum for cell viability assays or in a physiological saline solution for the in vivo experiment.
Splenic lymphocyte preparation
Spleens were dissected from mice fed normal diets (NDs). Splenic lymphocytes were prepared according to the method described by Smina et al. 23 Briefly, splenic lymphocytes were isolated by gently disrupting the spleen in cell culture medium with a brass wire cloth (200 mesh). After centrifugation at 1814 g for 5 min, the pellet was washed twice with phosphate-buffered saline (PBS), mixed, and separated by two-layer Percoll gradient centrifugation (40% and 70%). The viability of the splenic lymphocytes was determined by trypan blue dye exclusion.
Cell viability assay
Splenic lymphocytes were plated (1.0 × 104/well) on 96-well plates and incubated for 24 h. Then, the cells were exposed to OA and/or GLP (0–1000 μg/mL). After 48 h, 20 μL of CCK-8 reagent was added and incubated at 37°C for another 4 h. Cell viability was measured at 490 nm using an enzyme-linked immunosorbent assay reader (MK3; Thermo Fisher Scientific, Waltham, MA, USA). The inhibitory rate was calculated as follows: inhibitory rate (%) = (1 − the average OD value of the treatment group/the average OD value of the control group) × 100%.
Western blotting analysis
Lymphocytes were isolated from the spleens of mice fed a ND and incubated for 24 h. Subsequently, the cells were exposed to 500 μm OA and/or 0.6 mg/mL GLP for different times. The total protein was extracted, and the protein concentration of the supernatant was measured by bicinchoninic acid (BCA) assay. Then, the lysates were separated on 10% sodium dodecyl sulfate polyacrylamide gels and transferred onto polyvinylidene fluoride membranes. The blots were blocked in Tris-buffered saline (TBST; containing 0.1% Tween 20), supplemented with 5% bovine serum albumin, and then probed with the following primary rabbit polyclonal antibodies at 4°C overnight: Bcl-2 (1:1000, rabbit polyclonal antibody; Proteintech, Wuhan, China), Bax (1:2000, rabbit polyclonal antibody; Proteintech), caspase-3 (1:1000, rabbit polyclonal antibody; Cell Signaling Technology, Danvers, MA), and β-actin (1:4000, mouse monoclonal antibody; Proteintech). Following three washes with TBST, the membranes were probed with horseradish peroxidase (HRP)-conjugated secondary antibodies (1:6000, polyclonal goat anti-rabbit immunoglobulin G [IgG] HRP, and 1:5000, polyclonal goat anti-mouse IgG HRP; Proteintech) at room temperature for 1 h. The specific proteins were visualized using an enhanced chemiluminescence detection kit according to the manufacturer's instructions. Densitometry was performed using ImageJ software v1.8.
Animals and treatment
C57BL/6 mice (6–8 weeks old, male) and normal and HDs were purchased from Tianqin Biological Technology Co. Ltd. (Changsha, China). The mouse license for the laboratory (no. SCXK 2016-0002) was issued by the National Science and Technology Department of Hunan Province. Animal experiments were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals and the related ethical regulations of Hunan Agricultural University approved by the Ministry of Health, China. The mice were housed in a room with an alternating 12 h light/12 h dark cycle at 23°C ± 1°C. After acclimatization for 1 week and ND feeding (Table 1), 100 mice were randomly assigned to one the following five groups (four cages per each treatment): normal diet (ND, 1.37 kcal/g), high-fat diet (HD, 3.45 kcal/g), high-fat diet + a low dose of GLP (HD +200 mg/kg bw GLP, HDL), high-fat diet + a high dose of GLP (HD +400 mg/kg bw GLP, HDH), and high-fat diet +20 mg/kg bw LA (HLA, used as a positive control). GLP and LA were dissolved in physiological saline and administered once daily by oral gavage. All mice were allowed free access to the test diets for 12 weeks.
Composition of the Diets
Vitamins premix (mg/100 g diet): VB1 0.999, VB2 0.999, VB3 4.998, VB5 2.665, VB6 1.166, VB11 0.333, VB12 0.0017, VH 0.033, VA 0.22, VD3 0.0042, VE 8.329, VK3 0.333.
Minerals premix (mg/100 g diet): FeSO4·7H2O 26.754, CuSO4·5H2O 2.532, MnSO4·H2O 16.536, ZnSO4·7H2O 14.112, Na2SeO3 0.00234, KI 0.0426·FeSO4.
HD, high-fat diet; ND, normal diet.
Collection of organ, tissue, and serum samples
Mice were anesthetized with an intraperitoneal injection of 10% chloral hydrate (Sinopharm Chemical Reagent Co. Ltd., Shanghai, China). Then, blood was collected from the orbital sinus, stored at 4°C overnight, and centrifuged at 1000 g for 20 min. The supernatant was aspirated carefully and analyzed immediately. After decapitation, tissue samples were collected immediately, weighed on ice, and stored at −80°C for later use. The tissue index was calculated as follows: tissue index = (wet weight of tissue/body weight) × 100%. 24
Serum lipid analysis
Serum TC, TG, LDL-C, and HDL-C levels were measured by the colorimetric enzymatic methods using commercial kits, according to the manufacturer's instructions. Then, the samples were analyzed on a Multiskan Microplate Reader (MK3; Thermo Fisher Scientific).
Antioxidant enzymes activities
The total protein concentration of the small intestine was determined by BCA protein assay (Beyotime). The WST-8 colorimetric method was applied to detect superoxide dismutase (SOD) activity, as described by Ukeda et al. 25 The total SOD activity was evaluated using a Total Superoxide Dismutase Assay Kit (Beyotime) and a spectrophotometer (MK3; Thermo Fisher Scientific) at 560 nm. CAT activity was measured using a catalase assay kit (Beyotime), as described by Xia et al. 26 The antioxidant enzymes activity is expressed as units per milliliter for serum or milligram protein for the small intestine.
Measurement of MDA and GSH-Px levels
The small intestine was weighed and homogenized on ice in nine volumes of physiological saline. The MDA content was detected using thiobarbituric acid and presented as mmol per milliliter or milligram protein. GSH-Px level was measured using a 5,5′-Dithiobis-(2-nitrobenzoic acid) reaction test, and the results are expressed as units per milliliter or milligram protein for serum and small intestine samples, respectively. All steps were performed in accordance with the manufacturer's protocol.
Fluorescence observation of splenic lymphocytes
The morphology of splenic lymphocytes was assessed using Hoechst 33258 staining. After splenic lymphocytes were extracted from each group, the cells were fixed in 100% ice-cold methyl alcohol overnight at 4°C, permeabilized, and stained with 2 μg/mL of Hoechst 33258 solution for 5 min in the dark. Cell morphology was observed under a fluorescence microscope (CKX41; Olympus, Tokyo, Japan).
Determination of splenic lymphocyte apoptosis
Apoptosis was investigated using an Annexin V-FITC Apoptosis Detection Kit. Splenic lymphocytes (1 × 106/mL) were prepared from each in vivo group. Splenic lymphocytes were extracted from normal mice and then exposed to GLP (0.4 and 0.8 mg/mL) and 500 μM OA for 24 h in vitro experiment. After the cells were centrifuged at 1000 g for 10 min at 4°C, the precipitate was washed twice with 1 mL of precooled PBS, centrifuged, and suspended in 200 μL of binding buffer. Subsequently, the cells were stained with 10 μL of Annexin V-FITC and 10 μL of propidium iodide (PI) for 30 min at 4°C in the dark. Cell apoptosis was analyzed by flow cytometry (Beckman Coulter, Inc., Miami, FL, USA) within 30 min of staining. Annexin-V and PI signals were monitored at wavelengths of 525 and 630 nm, respectively.
Statistical analysis
All statistical analyses were performed using PASW Statistics for Windows v18.0 (SPSS, Inc., Chicago, IL, USA). The mean and standard deviation for each group are from replicate experiments. The statistical significance of differences between groups was examined by one-way analysis of variance followed by Tukey's post hoc test. The results were considered statistically significant at P < .05 and extremely significant at P < .01.
Results
GLP suppressed OA-induced cell viability loss in splenic lymphocytes
To evaluate the effect of GLP on the OA-induced loss of cell viability, splenic lymphocytes were treated with various concentrations of OA and/or GLP in vitro. As shown in Table 2, the exposure of splenic lymphocytes to OA dose-dependently inhibited cell growth, while this inhibition was markedly attenuated by GLP (50–1000 μg/mL) in a dose-dependent manner. Compared to control treatment, high concentration GLP treatment promoted cell growth, even in the presence of OA. Additionally, 50–1000 μg/mL of GLP showed no cytotoxicity to normal splenic lymphocytes.
Effect of Ganoderma lucidum (Reishi Mushroom) Polysaccharides on Oleic Acid-Induced Splenic Lymphocytes Viability In Vitro
The data are presented as mean ± SD (n = 6). Values with different letters within a column are significantly different by Tukey's post hoc test (P < .05).
GLP, Ganoderma lucidum (Reishi mushroom) polysaccharides; OA, oleic acid; SD, standard deviation.
GLP alleviated OA-induced apoptosis in splenic lymphocytes
As shown in Figure 1, compared to control treatment, 500 μM of OA stimulated obvious increases in cell apoptosis (Fig. 1A) and Bax and cleaved caspase-3 (17 and 19 kDa) protein levels at 48 h. The Bax/Bcl-2 ratio was significantly higher after OA treatment than control treatment at 48 h. In contrast, GLP increased Bcl-2 and caspase-3 expression levels in OA-treated lymphocytes, while simultaneously decreasing Bax and cleaved caspase-3 (17 and 19 kDa) expression levels and the Bcl-2/Bax ratio at 6–48 h (Fig. 1B, C). GLP also reduced the apoptotic rate mediated by OA in splenic lymphocytes (Fig. 1A).

GLP regulated the expression of apoptotic-related proteins of splenic lymphocytes induced by OA. Cell apoptotic rate
GLP induced changes in body weight and relative organ weight
As shown in Table 3, the HD group showed higher body weight gains than the ND group, but the group fed the HD while receiving a high dose of GLP had lower body weights than the HD group. The group fed the HD and receiving a low dose of GLP did not have significant changes in HD-induced body weight gain. Compared to the ND group, the high-fat group had increased relative weights for the liver, heart, perirenal fat and epididymal fat as well as reduced relative weights for the spleen and kidney (P < .05). GLP were shown to decrease the relative weights for the liver, heart, fat tissues, spleen, and kidneys after HD feeding, suggesting that GLP substantially influence body weight and relative organ weight in mice fed a HD.
The Weight and Index of Organs in High-Fat Diet Fed C57BL/6 Mice
The data are presented as mean ± SD (n = 6). Values with different letters within a column are significantly different by Tukey's post hoc test (P < .05).
HDL, HD group plus 200 mg/kg GLP group; HDH, HD group plus 400 mg/kg GLP group; HLA, HD group plus 20 mg/kg LA.
HDL, high-density lipoprotein; LA, lipoic acid.
GLP regulated serum lipid levels
To determine the anti-obesity effect of GLP further, serum lipid levels were examined. As shown in Figure 2, compared to those in the ND group, the serum lipid levels, including TC, TG, LDL-C, and HDL-C, were increased significantly by 145%, 222%, 164%, and 139% in the HD group, respectively. Notably, GLP effectively increased HDL-C levels and decreased TG and LDL-C levels in the serum (P < .01 and P < .05) after HD feeding, whereas GLP had no effect on the increase in serum TC levels.
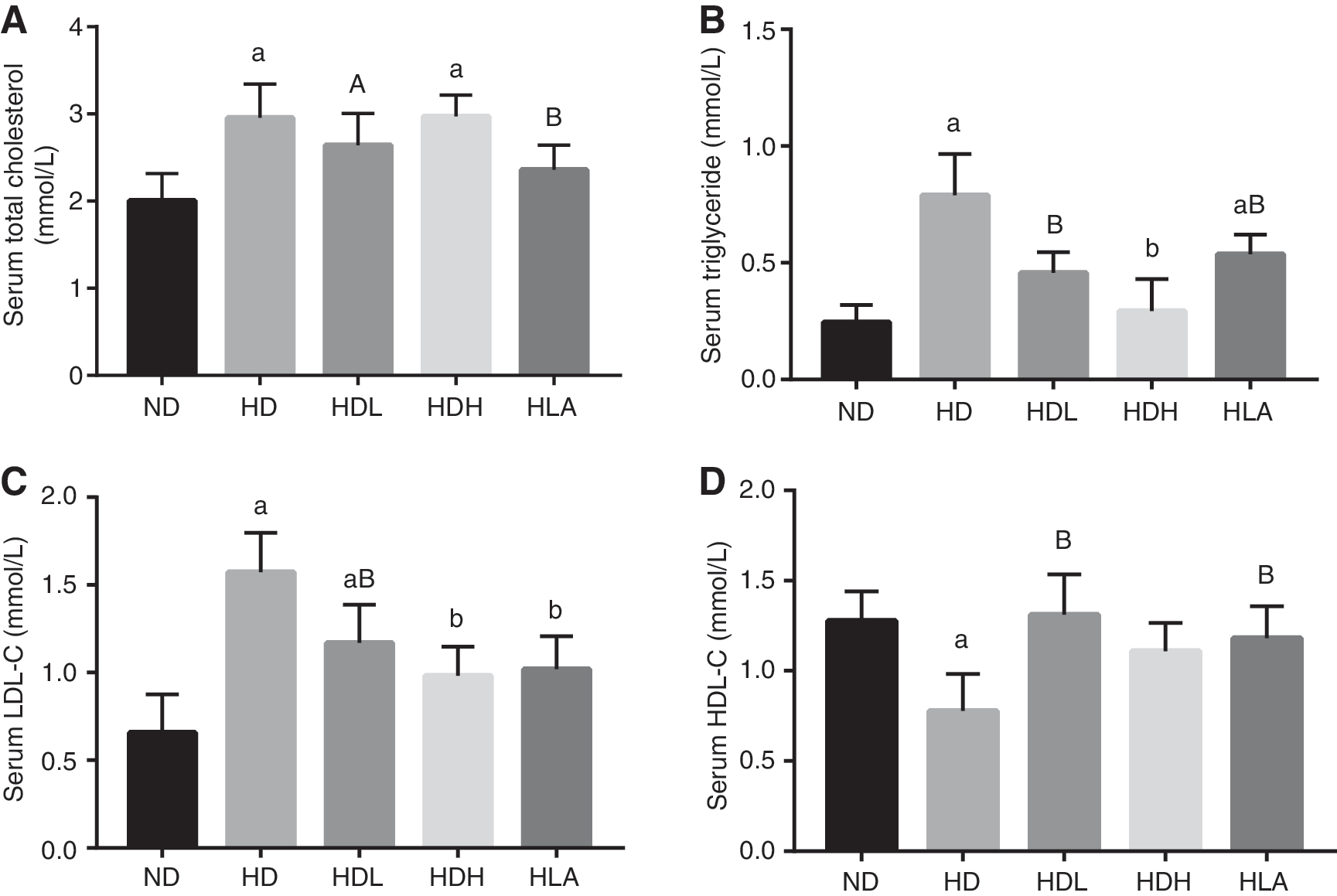
GLP decreased serum lipid levels in HD fed C57BL/6 mice. At the end of the experimental period,
GLP reduced oxidative stress in the serum and small intestine
The activities of serum CAT (Fig. 3A), SOD (Fig. 3C), and GSH-Px (Fig. 4C) were obviously lower in the HD group than in the ND and HD + GLP groups (P < .01 and P < .05). The serum MDA level was higher in the HD group than in the ND and HD + GLP groups (Fig. 4A). Furthermore, there was no significant difference in MDA, SOD, and GSH-Px levels between the ND and HD + GLP groups. A similar trend for CAT, SOD, GSH-Px, and MDA levels in the small intestine tissues was observed (P < .01 and P < .05).

GLP increased SOD and CAT activities in serum and small intestine of C57BL/6 mice. At the end of the experimental period,

GLP decreased MDA and GSH-Px levels in serum and small intestine of C57BL/6 mice. At the end of the experimental period, the levels of
GLP prevent splenic lymphocyte apoptosis in vivo
As shown in Figure 5A, almost all splenic lymphocytes in the ND group showed cytoplasmic diffusion and dark blue fluorescence. The cellular fluorescence intensity was significantly higher in the HD group than in the ND group. Moreover, some characteristic apoptotic morphologies, including chromatin condensation and nuclei fragmentation, were observed in splenic lymphocytes from mice fed the HD. GLP significantly decreased the cellular fluorescence intensity and number of fragmented nuclei in splenic lymphocytes induced by the HD in the HD + GLP groups compared to the HD group. The effect of GLP treatment on splenic lymphocyte apoptosis was also determined by flow cytometry. As shown in Figure 5B and C, the HD caused a markedly higher induction of late apoptosis (18.00% ± 2.6%) and early apoptosis (27.79% ± 3.2%) in splenic lymphocytes compared to the ND group (P < .01), whereas the rate of cell apoptosis was significantly suppressed by GLP treatment compared to the HD group (P < .01). These findings suggest that the protective action of GLP against typical nuclear fragmentation and conspicuous nuclear staining are associated with the cell apoptosis induced by HD as shown in Figure 5A. Splenic lymphocytes in the HLA group also showed decreases in nuclei fragmentation and cell apoptosis (data not shown).

Effects of GLP on apoptosis of splenic lymphocytes in HD fed C57BL/6 mice.
Discussion
HD-induced obesity is characterized by the significant accumulation of excess adipose tissue in animals and humans, and body weight reduction is the recommended first-line treatment. 27 GLP have been proven to be potent antioxidants capable of scavenging/neutralizing an array of reactive oxygen species, including hydroxyl anions, superoxide anion, hydrogen peroxide, and 1,1-diphenyl-2-picrylhydrazyl radicals. Previous work reported that GLP mediate apoptosis in the human colorectal cancer cell line HCT-116, activating the mitogen-activated protein kinase and mitochondrial pathways. 22 However, the effect of GLP on HD-induced oxidative stress and the mechanisms remain elusive. In this study, various concentrations of GLP (>10 kDa) alleviated the inhibition of proliferation in splenic lymphocytes incubated with OA, and a high concentration of GLP could promote splenic lymphocyte proliferation in vitro, even in the presence of OA (Table 2). Moreover, GLP reduced the ratio of Bax/Bcl-2 and increased the level of the pro-apoptotic protein caspase-3 at different times in OA-treated splenic lymphocytes (Fig. 1). These findings suggest a protective effect of GLP against OA-mediated lymphocyte apoptosis though decreasing the Bax/Bcl-2 ratio and suppressing caspase-3 activation in splenic lymphocytes.
These results prompted an investigation of the protective effect of GLP in vivo in C57BL/6 mice fed a HD. The data showed that oral GLP treatment for 12 weeks reduced the increases in body weight and relative weights of the liver, heart, and white adipose tissues induced by a HD in C57BL/6 mice. These results indicate that GLP could prevent the risk of obesity, fatty liver, and fatty heart induced by HD consumption. A previous study reported that a water extract of G. lucidum mycelium and its polysaccharides (>300 kDa) decreased body weight gain and liver, epididymal fat, and subcutaneous fat weights in mice fed a HD. 28 Interestingly, the change in body weight and adipose tissue weights induced by different molecular weight of GLP showed the same trends, indicating that GLP are the main component of G. lucidum and contribute to its anti-obesity effect. Cholesterol constitutes a well-known component of the body's cellular membrane, and it is involved in controlling the liquidity of biofilm. 29 Like TG, cholesterol circulates in the blood but is deposited in body fat and used when the body needs extra energy. 30 LDL particles contain the majority of cholesterol and can accumulate in the walls of blood vessels in the body. 31 HDL absorbs cholesterol and carries it back to the liver. 32 A high LDL-C level raises the risk for health problems, while a high HDL-C level might be attributed to a lower risk. The results of the Lipid Research Clinics Primary Prevention Trial indicate that elevated serum cholesterol and TG levels, particularly LDL-C induced by a HD, are intimately related to a high risk of cardiovascular disease (e.g., coronary artery disease), fatty liver disease (which can develop into nonalcoholic fatty liver disease), and type 2 diabetes mellitus. 33,34
In this study, the 200 and 400 mg/kg GLP doses chosen were used in vivo, as reported by Yang et al. 35 Compared to that in the HD group, serum lipid accumulation, including TC, TG, and LDL-C levels, was significantly suppressed, and the HDL-C levels were higher in mice fed the HD plus GLP, indicating that GLP improved the serum dyslipidemia resulting from the HD. This observation is in agreement with Seto et al., who showed an inverse correlation between G. lucidum consumption and serum TG and TC levels in +db/+db mice. 36 Similar results were also obtained after treatment with LA. In this experiment, LA was used as a positive control because it has been shown to decrease body weight considerably in overweight or obese mice via inhibiting oxidative stress, lipogenesis, and inflammatory reactions. 37,38 It could be inferred that GLP are potent hypocholesterolemic agents. Although the beneficial effects of dietary GLP intake on serum lipid concentrations had not been reported in mice fed a HD, greater reductions in lipid concentrations may still be obtained with a higher dosage of GLP.
The small intestine is extremely sensitive to reactive oxygen species and is a primary site for the action of dietary antioxidants. 39 Previous studies have shown that diabetes-induced oxidative stress in the small intestine is possibly associated with concomitant functional alterations, which could be suppressed by antioxidants. 40,41 The inhibition of diabetes-induced oxidative damage to the small intestine may be effective in overcoming oxidative stress. Excess dietary cholesterol consumption has been considered as a contributory factor to oxidative stress and body organ malfunction in vivo. 42 Thus, in this study, the levels of enzymatic and non-enzymatic antioxidants were selected to determine the antioxidant effects of GLP in mice, with special emphasis on the serum and small intestine. The results showed that a HD reduced antioxidant defense mechanisms (CAT, SOD, and GSH-Px) and increased MDA production. The observed increases in lipid peroxidation levels in the small intestine are in agreement with similar findings in the serum. 39 It is noteworthy that GLP efficiently suppressed elevated MDA levels and decreased CAT, SOD, and GSH-Px activities in the serum and small intestine, suggesting that GLP play an antioxidant role in the prevention of oxidative stress in mice fed a HD. Such a protective effect of GLP might be associated with their ability to scavenge hydroperoxyl radical (OOH), singlet oxygen ( 1 O2), peroxynitrite (ONOO−), peroxyl radicals (ROO•), hydroxyl radicals (•OH), and hypobrpmpus acid (HOBr). 43 The increased levels of lipid peroxidation cause the loss of reactive end products and alter membrane properties, followed by reactions with other molecules leading to DNA damage. 44 DNA damage triggers the apoptotic pathways leading to cell apoptosis, which is indicated by a series of characteristics, such as oligonucleosomal DNA fragmentation, chromatin condensation, apoptotic bodies, membrane surface blebbing, and phosphatidylserine externalization. 45 The current data revealed that GLP effectively ameliorated HD-induced early and late apoptosis in splenic lymphocytes in vivo and in vitro. Some investigations have proven that obesity-induced alterations in nutrition, endocrine function, and metabolism may contribute to changed immunocompetence. The spleens of obese (C57BL/6J ob/ob) mice weigh less and contain fewer Thy 1.2-positive lymphocytes and IgG-producing lymphocytes than those of lean controls. 46 Some approaches could alleviate the negative health impacts caused by obesity. For example, voluntary wheel exercise reduced H2O2-induced apoptosis in total splenocytes and CD4 + cells, as well as interferon-γ production, compared to no voluntary wheel exercise. 47 This study observed that a similar effect could be produced by GLP. The results indicate that GLP have hypolipidemic, antioxidant, and anti-apoptotic effects in HD fed-mice. It is inferred that GLP contributes to a decrease in MDA production in the serum and small intestine, resulting in the enhancement of the antioxidant enzymatic systems, followed by the inhibition of lipid peroxidation reactions and cell apoptosis. This work will benefit the development of future GLP applications for improving dyslipidemia and oxidative stress and opens a new line of research on the anti-apoptosis potential of GLP in immunocytes. The anti-obesity effect of GLP might involve the promotion of intestinal peristalsis though the inhibition of gut dysbiosis and regulation of bile acid and cholesterol metabolism through the modulation of lipogenic gene expression in obese animals. 28,48,49 Nonetheless, the hypothesis that GLP may also directly decrease food intake by slowing the gastric emptying rate, intestinal digestion, and absorption of a HD by effectively blocking the interaction, and contact between digestive enzymes and food to cause weight loss cannot be ruled out. 50
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 31501484), Hunan Provincial Natural Science Foundation of China (No. 2016JJ3081), Agricultural Science and Technology Innovation of Hunan Province (2017GP17), and Hunan Agricultural Science and Technology Innovation Team Project (2017GC03).
Author Disclosure Statement
No competing financial interests exist.