
Abstract
Dioscorea batatas Decne (Chinese yam), a perennial trailing plant, has been used as a traditional medicine due to its beneficial biological effects. Recent studies reported that yam bark powder increased antioxidant enzyme activity and exerted an anti-inflammatory effect in drug-induced liver injury or colorectal cancer in rats. In this study, we examined whether ethanol extracts of yam bark (peel) could ameliorate the macrophage-induced inflammatory damage of colonic epithelial cells in culture and in an acute colitis mouse model. Human HCT116 colorectal epithelial cells and/or lipopolysaccharide (LPS)-activated murine Raw 264.7 macrophages were treated with the yam bark extract, followed by biochemical examination of cellular cytokine and antioxidant enzyme expression levels. Our results demonstrated that the yam bark extract decreased nitric oxide production and proinflammatory protein expression in LPS-activated Raw 264.7 cells, and reduced the level of reactive oxygen species in hydrogen peroxide-challenged HCT116 cells. Moreover, yam bark extract attenuated dextran sulfate sodium (DSS)-induced acute colitis in BALB/c mice. These findings suggest that oral supplementation with yam bark extract inhibited DSS-induced colon inflammation through upregulation of antioxidant enzymes and subsequent downregulation of inflammatory proteins.
Introduction
I
Chronic inflammation increases the risk of developing colorectal cancer, in which oxidative stress plays an important role. 7 –9 Accumulation of reactive oxygen species (ROS) and nitrogen species is generally concurrent with inflammatory responses, which are preceded by upregulation of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), inducible nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2). In addition, nuclear factor (erythroid-derived 2)-like 2 (Nrf2), a key transcription factor, induces phase 2 antioxidant enzymes, such as heme oxygenase-1 (HO-1), NAD(P)H quinone dehydrogenase 1 (NQO1), glutathione reductase, and gamma-glutamylcysteine synthetase, to suppress NF-κB-mediated inflammation. 10 –12 Nrf2 activators could thus counteract the NF-κB signaling pathway, and thereby suppress the inflammatory response.
Dioscorea batatas Decne (a synonym of Dioscorea polystachya Turcz.) is widely distributed in East Asian countries, including China, Japan, and Korea, and is commonly known as Chinese yam. The plant has been considered as a medicinal herb due to its functional ingredients reportedly exerting antioxidative and anti-inflammatory effects. 13 –17 Multiple studies have demonstrated that yam bark can inhibit iNOS and COX-2 expression in mouse macrophages, decrease prostaglandin D2 and leukotriene C4 production in mouse bone marrow-derived mast cells, and protect hepatocytes from carbon tetrachloride-induced toxicity in rats. 18,19 In particular, diosgenin, which is reported to be present in wild yam, has been shown to inhibit colon carcinogenesis. We thus hypothesized that ethanol extracts of yam bark could prevent chronic colitis. As such, this study investigated the preventive effect of yam bark ethanol extracts against chronic colitis using both human colorectal epithelial cells and a dextran sulfate sodium (DSS)-induced acute colitis mouse model.
Materials and Methods
Preparation of D. batatas Decne ethanol extracts
Dry powdered D. batatas Decne bark, which had been air-dried and finely ground, was obtained from the Forest Resources Development Institute of Gyeongsangbuk-do (Andong, S. Korea). The ethanol extract of D. batatas Decne bark (DBE) was prepared as follows: 50 g of D. batatas bark powder was suspended in 250 mL 40%, 60%, 80%, or 95% (v/v) ethanol in distilled water (DW) and shaken at 250 rpm for 1 h at 50°C using a shaking incubator (Daihan Labtech Co., Ltd., Namyangju, S. Korea). After filtering through filter paper (8 μm pore size; Whatman, Little Chalfont, United Kingdom), filtrates were vacuum-evaporated (EYELA N-1000, Tokyo, Japan) and subsequently freeze-dried. Lyophilized DBE was dissolved in 80% (v/v) dimethyl sulfoxide in DW for further examination.
Cell culture
A murine macrophage cell line (Raw 264.7) and human colon cancer cell line (HCT116) were obtained from the Korean Cell Line Bank (Seoul, S. Korea). Cells were cultured in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Welgene, Gyeongsangbuk-do, S. Korea) and 1% penicillin-streptomycin (Invitrogen/Thermo Fisher Scientific, Carlsbad, CA, USA), and maintained at 37°C in an atmosphere of 5% CO2/95% air under saturating humidity.
Cell viability assay
To test the cytotoxicity of DBE, a cell counting kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) assay was used as previously described. 20 Cells were treated with various concentrations of DBE followed by incubation for 24 h. The absorbance, which is proportional to the number of living cells in each well, was measured at 450 nm using a microplate reader (Sunrise™; Tecan Group Ltd., Männedorf, Switzerland).
Antioxidant response element-luciferase activity assay
To measure the transcriptional activity of the antioxidant response element (ARE), luciferase reporter gene assay was conducted on HepG2-ARE and HCT116-ARE cells as previously described. 21 Cells were treated with various concentrations of DBE in 0.5% FBS-containing culture medium for 24 h. ARE-luciferase activity was measured using a luciferase assay system (Promega, Madison, WI, USA) according to the manufacturer's instructions. Sulforaphane (Sigma-Aldrich, St. Louis, MO, USA) or SFSA 22 (Sigma-Aldrich) was used as the positive control. Luminescence was detected using a TD-20/20 luminometer (Turner Designs, Sunnyvale, CA, USA), and calibrated with the amount of total protein. Final values were normalized against the untreated control.
Detection of intracellular ROS
Intracellular oxidative stress was assessed using reduced 2′,7′-dichlorofluorescein diacetate (H2DCFDA; Invitrogen), which forms fluorescent 2′,7′-dichlorofluorescein when oxidized by ROS, as previously described with minor modifications. 19 Cells were plated into a black-bottom 96-well plate (Nunc, Rochester, NY, USA) at a density of 2 × 104 Raw 264.7 cells/well or 2.5 × 104 HCT116 cells/well. After a 24 h preincubation period, cells were incubated with varying concentrations of the extracts for another 24 h. Cells were subsequently challenged with hydrogen peroxide (H2O2, 500 μM in 1% FBS-containing phosphate-buffered saline [PBS]) for 30 min, and thereafter treated with H2DCFDA (20 μM) for 30 min. After 1 h-incubation in H2O2-containing PBS, fluorescence was measured at excitation and emission wavelengths of 485 and 535 nm, respectively, using a fluorescence microplate reader (Infinite 200; Tecan, Grodig, Austria).
Determination of nitric oxide production
Raw 264.7 cells were incubated with various DBE concentrations (0, 25, 50, and 100 μg/mL) and/or lipopolysaccharide (LPS, 1 μg/mL; Sigma-Aldrich). After 24 h, culture medium was collected and nitric oxide (NO) concentration determined by measuring nitrite content using the Griess reagent system (Promega).
Measurement of inflammatory cytokine levels
The levels of cytokines present in the culture medium were measured using commercial ELISA kits for interleukin (IL)-1β, IL-6, IL-12, tumor necrosis factor (TNF)-α, and granulocyte-macrophage colony-stimulating factor (GM-CSF) (all from BD Biosciences, San Jose, CA, USA).
Animals
Male BALB/c mice (6-week old, 20–22 g body weight [b.w.]) were purchased from Daehan BioLink (Eumseong, S. Korea). Mice were acclimatized for 1 week under standard conditions (room temperature of 20–25°C, a relative humidity of 45–55%, and a standard 12 h light-dark cycle), and fed a regular diet (Daehan BioLink) and water ad libitum.
DSS-induced colitis mouse model establishment
All animal studies were approved and conducted according to the guidelines of the Institutional Animal Care and Use Committee of the Kyungpook National University (approval number: KNU 2017-85). After 1 week of animal acclimatization, colitis was induced in 7-week-old BALB/c mice by oral ingestion of 3% (w/w) DSS-containing drinking water for 10 days. A total of 48 mice were randomly assigned to six experimental groups (eight mice per group; Table 1); group 1 received the solvent only, group 2 received 3% DSS only, while group 3, 4, and 5 received 3% DSS and 50, 100, or 200 mg DBE/kg b.w., respectively. Group 6, a positive control, received 3% DSS and SFSA (200 mg/kg b.w.) that is used as a medication to treat UC and Crohn's disease. Samples (DBE or SFSA) were orally administered four times (day 0, 4, 7, and 11) in the entire experimental period, followed by sacrifice at day 12. The solvent was composed of 30% (v/v) polyethylene glycol, 10% (v/v) Tween-20, and 10% (v/v) ethanol in saline. DBE and SFSA were dissolved in solvent and administered using oral gavage. During the entire experimental period, mouse b.w. was regularly monitored. The disease activity index (DAI) value was evaluated as the sum of the scores for weight loss, stool consistency, and blood in feces. 23 All animals were sacrificed 24 h after final treatment with the sample extract. Blood, liver, and colon samples were collected for further histological and biochemical analysis.
Experimental Animal Groups
A total of 48 mice were randomly assigned to six groups (eight mice per group).
The solvent was composed of 50% (v/v) polyethylene glycol, 10% (v/v) Tween-20, and 10% (v/v) ethanol in saline.
DBE, 95% ethanolic extract of Dioscorea batatas Decne peel.
SFSA, sulfasalazine used as the positive control.
b.w., body weight; DSS, dextran sulfate sodium.
Histological analysis
Collected colon tissues were fixed in a 10% (v/v) formalin solution and then paraffin-embedded as previously described. 24 Paraffin blocks were sectioned with a 5-μm thickness using a microtome (RM-2125 RT; Leica, Nussloch, Germany). Tissue sections were subsequently stained with hematoxylin and eosin dyes. Tissue damage was observed under a microscope (Eclipse 80i; Nikon, Tokyo, Japan).
Measurement of inflammatory cytokine levels in mouse serum and colon tissue
The levels of cytokines present in plasma were measured using commercial ELISA kits for IL-1β, IL-6, IL-12, TNF-α, and GM-CSF (all from BD Biosciences). Colon tissues were collected from sacrificed mice and homogenized by ultra-sonication in 0.1 M phosphate buffer for 15 sec at 40 V. The homogenate was centrifuged at 15,000 g at 4°C for 30 min, and the supernatant assessed for each cytokine using the respective ELISA kits.
Western blot analysis
Whole cell or tissue lysates were prepared by homogenization in precooled lysis buffer (20 mM Tris-HCl, 145 mM NaCl, 10% glycerol, 5 mM EDTA, 1% Triton-X, and 0.5% Nonidet P-40). After centrifugation at 15,000 g at 4°C for 10 min, the supernatants were quantified and denatured in sample buffer at 95°C for 10 min. Alternatively, nuclear and cytoplasmic proteins were fractionated using the NE-PER™ nuclear and cytoplasmic protein extraction kit (Thermo Fisher Scientific, Rockford, IL, USA), quantified, and denatured. Isolated proteins were then electrophoretically separated on sodium dodecyl sulfate polyacrylamide gels and subsequently transferred to a polyvinylidene fluoride (PVDF) membrane (Merck Millipore Corp., Billerica, MA, USA). PVDF membranes were blocked in 1% bovine serum albumin (BSA) in Tris-buffered saline including 0.1% (v/v) Tween 20. The primary antibodies used in this study were as follows; COX-2 (Cell Signaling, Danvers, MA, USA), iNOS (Cell Signaling), HO-1 (Santa Cruz Biotechnology, Dallas, TX, USA), NQO1 (Santa Cruz Biotechnology), phosphorylated-p65 (p-p65) (Santa Cruz Biotechnology), p65 (Santa Cruz Biotechnology), Nrf2 (Abcam), lamin B, and β-actin (Santa Cruz Biotechnology). Appropriate secondary antibodies conjugated to horse radish peroxidase (Santa Cruz Biotechnology) were used to target each respective primary antibody. Protein bands were visualized using SuperSignal™ West Pico Chemiluminescent Substrate (Pierce, Cheshire, United Kingdom), digitally imaged using ImageQuant LAS 4000 mini (GE Health care Life Sciences, Little Chalfont, United Kingdom), and densitometrically analyzed using Image Studio Lite version 5.2 (LI-COR Biotechnology, Lincoln, NE, USA).
Statistical analysis
All statistical analyses were performed using SPSS software version 23.0 (SPSS, Inc., Chicago, IL, USA). Comparisons were conducted via one-way analysis of variance followed by Duncan's multiple range test. P-values <.05 were considered to be statistically significant. Significant differences are indicated using different alphabetical letters.
Results
DBE increases ARE-luciferase activity in HCT116-ARE cells
The ARE-mediated transcriptional activation potential of DBE was increased in human epithelial cell lines carrying the ARE-luciferase construct (Supplementary Fig. S1; Supplementary Data are available online at

Cellular antioxidant activity of DBE ethanol extracts.
DBE reduces intracellular ROS levels in H2O2-challenged HCT116 and Raw 264.7 cells
To examine whether DBE lowers intracellular ROS levels, HCT116 and Raw 264.7 cells were treated with varying extract concentrations after treatment with H2O2 (Fig. 1B, C). Intracellular ROS levels were increased by H2O2 treatment, but decreased with DBE co-treatment in a concentration-dependent manner.
DBE decreases inflammation-related factors in LPS-stimulated Raw 264.7 cells
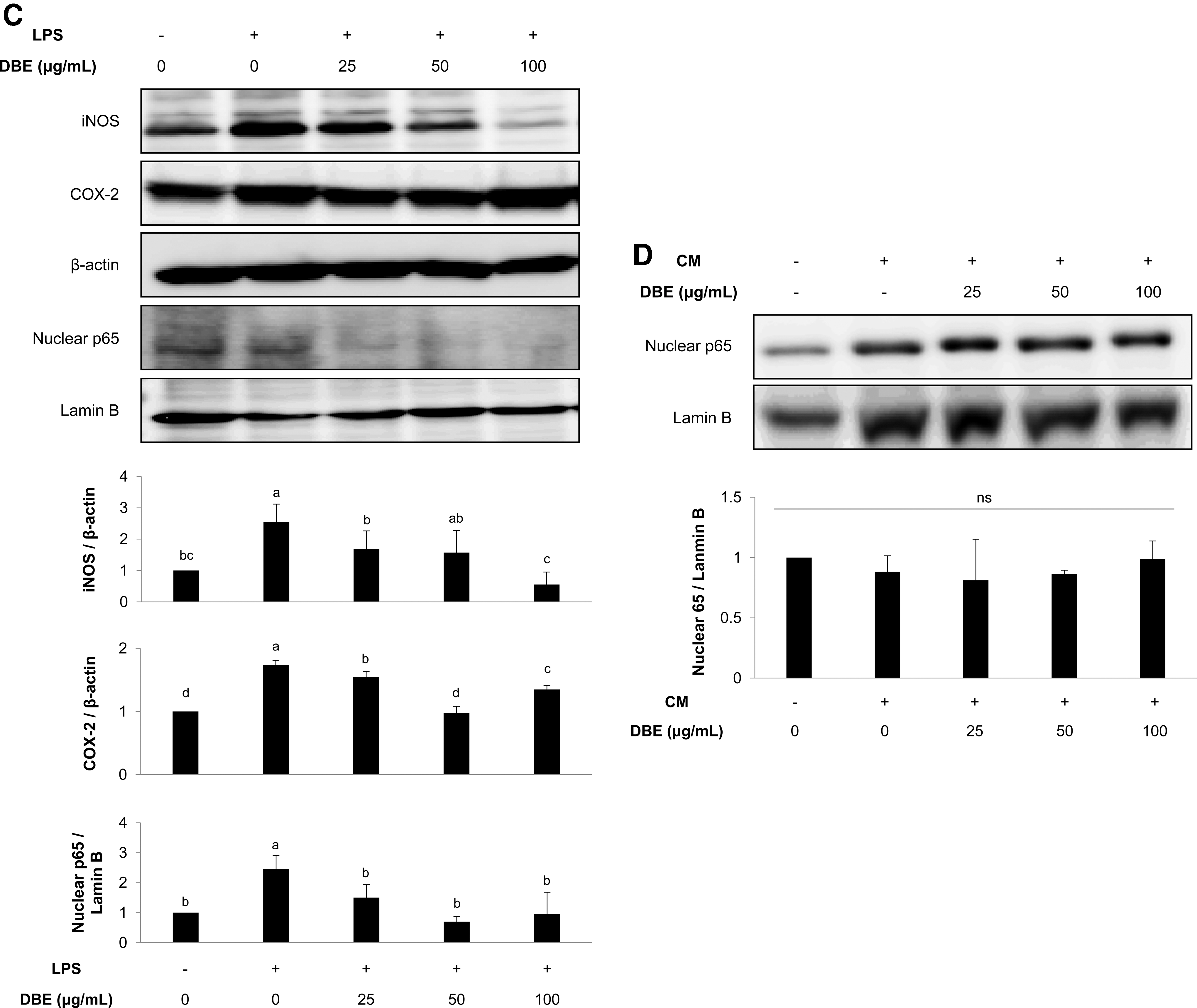
To examine the anti-inflammatory activity of DBE, Raw 264.7 cells were stimulated with LPS and then treated with varying extract concentrations. Thereafter extracellular NO levels (Fig. 2A) and proinflammatory cytokine levels, including IL-6, TNF-α, and GM-CSF (Fig. 2B), were measured. DBE decreased NO and cytokine (IL-6, TNF-α, and GM-CSF) levels in the culture medium of LPS-stimulated Raw 264.7 cells in a dose-dependent manner. Moreover, cytoplasmic iNOS and COX-2, and nuclear NF-κB p65 levels, which increased after LPS stimulation, were reduced by DBE (Fig. 2C). Interestingly, DBE did not affect nuclear p65 levels in HCT116 cells incubated in conditioned medium harvested from LPS-stimulated Raw 264.7 cell culture (Fig. 2D).

DBE reduces inflammatory factors in LPS-activated Raw 264.7 macrophage cultures.
DBE upregulates Nrf2 and its downstream antioxidant enzymes
We further examined whether DBE promotes Nrf2-mediated antioxidant enzyme expression in HCT116 cells. When treated with extract, protein expression levels of the Nrf2 downstream antioxidant enzymes HO-1 and NQO1 were significantly increased in HCT116 cells compared with the control (Fig. 3A). In addition, HCT116 cells treated with extract, and subsequently exposed to conditioned medium from LPS-stimulated Raw 264.7 cell culture, displayed increased nuclear Nrf2 levels with extract at ≥50 μg/mL, while the conditioned medium had no discernible influence (Fig. 3B).

DBE upregulates Nrf2 and its downstream antioxidant enzymes in HCT116 cells.
Oral supplementation of DBE mitigates tissue damage in a DSS-induced colitis mouse model
To examine the in vivo anti-inflammatory effect of DBE, an acute colitis mouse model was employed. DBE was orally administered at 50, 100, or 200 mg/kg b.w. every 3 days during the entire experimental period of 12 days, while DSS was administered in drinking water 2 days after initiating DBE treatment and continued until the experiment was completed, as illustrated in Figure 4A. Average b.w. was significantly reduced with DSS treatment (Fig. 4B). DBE treatment at a dose of 100 mg/kg b.w. alleviated DSS-caused b.w. loss, which was comparable to the SFSA-treated positive control. In addition, DBE at 100 mg/kg b.w. decreased the average DAI value, as determined by the summation of weight loss scores, stool consistency, and blood in feces 23 (Fig. 4C). DSS treatment also shortened the length of the large intestine and disrupted the mucosal epithelium; however, DBE administered at 100 mg/kg b.w. lowered the average DAI, although colon length was not recovered (Fig. 4D, E).

Oral supplementation with DBE alleviates histological damage in a DSS-induced colitis mouse model.

Oral supplementation of DBE reduces inflammatory cytokines and induces Nrf2-mediated antioxidant proteins
The levels of plasma inflammatory cytokines including IL-1β and IL-6 increased by DSS treatment was significantly suppressed in mice orally administered with 50, 100, and 200 μg DBE/kg b.w. (Fig. 5A). In addition, the plasma IL-12 level also showed similar trend except that the level was further increased in mice treated with 200 μg DBE/kg b.w. (Fig. 5A). The concentrations of IL-12 and GM-CSF, which initially increased in colonic tissues from DSS-treated mice, subsequently decreased after extract treatment (Fig. 5B). However, the colonic tissue levels of IL-6 and TNF-α were not affected by treatment with DSS or DBE (Fig. 5B).
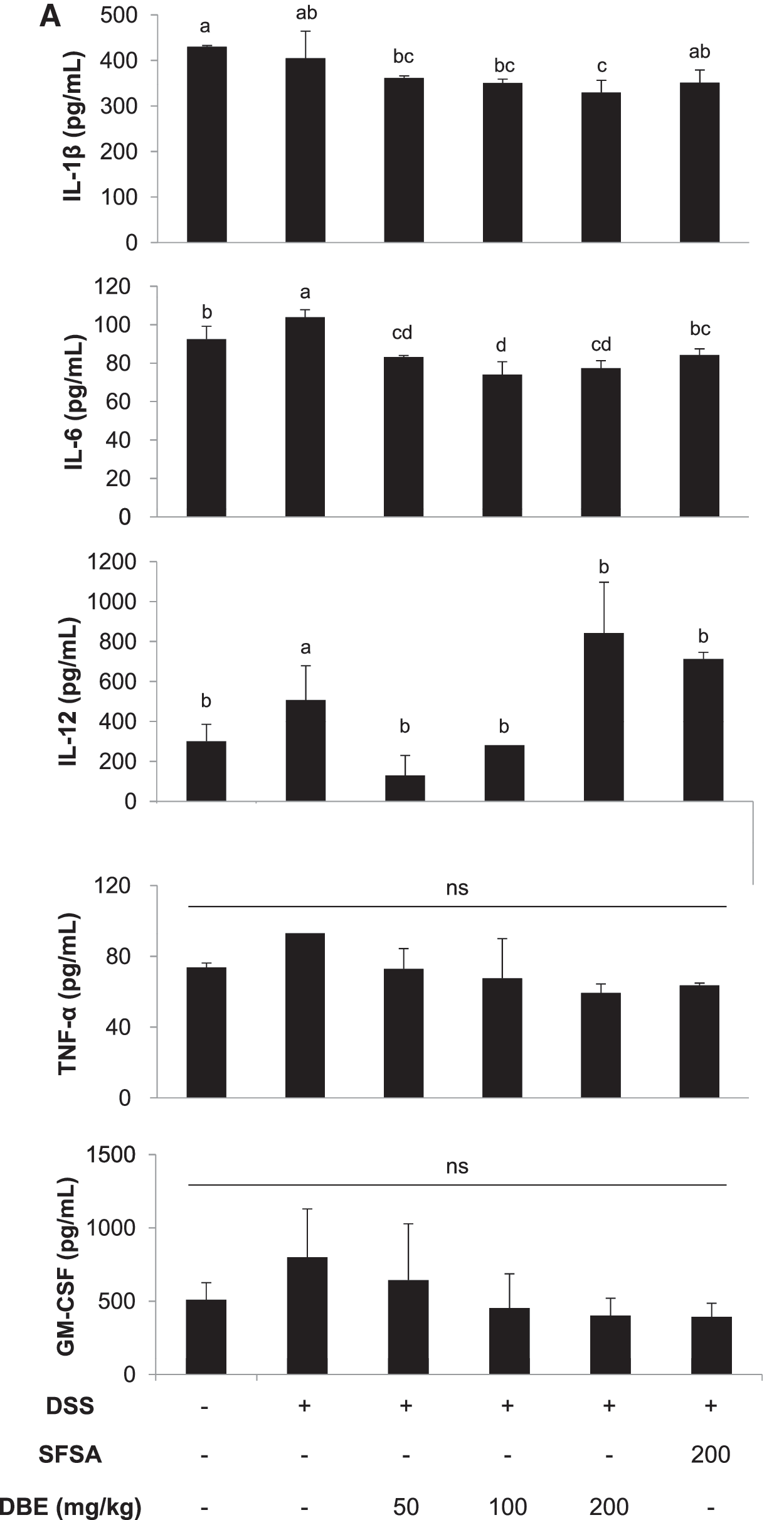
Oral supplementation with DBE reduces inflammatory cytokines and increases Nrf2-mediated antioxidant proteins in a DSS-induced colitis mouse model.


In addition, orally administering DBE decreased the expression levels of nuclear NF-κB p65, cytoplasmic iNOS, and COX-2 in the colonic tissue of DSS-treated mice (Fig. 5C). Moreover, the HO-1 protein level was enhanced in colon from mice treated with 50 and 100 μg DBE/kg b.w. whereas NQO1 level was significantly higher in colon from mice orally administered with 50 μg DBE/kg b.w. but was not different in the groups treated 100 μg DBE/kg b.w. and higher. Nuclear Nrf2 level in colon tended to increase in the groups administered with DBE, with no statistically significant difference (Fig. 5C). Meanwhile, DSS treatment itself did not affect the expression or levels of Nrf2 and its downstream enzymes including HO-1 and NQO1 although it significantly increased nuclear translocation of inflammatory mediator p65 and the expression of iNOS and COX-2 (Fig. 5C).
Discussion
Dioscorea species, belonging to the family Dioscoreaceae, are mostly grown for their starchy tubers or medicinal properties. 25,26 D. batatas is known to have biologically beneficial effects including antioxidative, antidiabetic, antiatherosclerotic, and anti-inflammatory activities. 13,14,17,27,28 Despite the diverse health benefits of D. batatas, little is known about the positive potential of the peel enriched with phytochemicals against colonic inflammation. It was recently reported that dietary supplementation with D. batatas root mitigated azoxymethane-induced colonic mucosal abnormalities, which are suggestive of colon cancer, and activated antioxidative enzymes in red blood cells and colonic mucosa in rats. 29 In this study, the anti-inflammatory effect of DBE bark was examined using a DSS-induced colitis mouse model along with Raw 264.7 and HCT116 cell lines.
DSS-induced colitis in mice mimics the clinical symptoms of human UC, including weight loss, colon shortening, diarrhea, and occult bleeding. 30,31 The DAI value and histology data showed that DSS treatment caused UC-like symptoms in BALB/c mice and was accompanied by inflammatory cytokines in the blood and colon, which was consistent with a previous report. 32 More importantly, orally administered D. batatas bark extract reduced DSS-induced colonic tissue damage and inflammatory cytokine production. Considering our in vitro results that D. batatas bark extract suppressed the induction of inflammatory cytokine expression in LPS-stimulated Raw 264.7 cells but not HCT116 cells, it can be reasonably inferred that D. batatas bark likely reduces the NF-κB-mediated inflammatory response in macrophages involved in host defense in mice with DSS-induced colitis.
In addition, we found that D. batatas bark extract decreased intracellular ROS in H2O2-challenged Raw 264.7 and HCT116 cells, and induced an Nrf2-mediated antioxidant response in HCT116 cells and colonic tissues. Multiple studies have demonstrated that ROS is one of the crucial factors that induces IBD, 33 while activation of the Nrf2 signaling pathway can reduce inflammatory damage in mice with DSS-induced colitis. 34 –37 Therefore, it is most likely that D. batatas bark extract could, in part, protect the colon from DSS-induced oxidative damage by activating the Nrf2-mediated antioxidant machinery. In particular, HO-1, which is a typical downstream antioxidant enzyme regulated by Nrf2, has been reported to exert potent anti-inflammatory effect by generating carbon monoxide (CO). 38 CO was reported to significantly suppress the NO generation and TNF or IL-6 production in LPS-stimulated macrophages and also increases production of anti-inflammatory cytokine, IL-10, in macrophages. 39
Several substances obtained from D. batatas or its derivatives were identified as pharmacologically active components. 26 For example, bark's ethanol extracts inhibit NO and prostaglandin E2 production in LPS-induced Raw 264.7 cells via NF-κB and ERK1/2 inhibition, 17 which was consistent with our in vitro results. Considering previous studies that have addressed active components in the whole tuber of D. batatas, such as glycoproteins in water extracts 16 or 6-hydroxy-2,7-dimethoxy-1,4-phenanthraquinone in methanol extracts, 40 the anti-inflammatory activities of D. batatas bark ethanol extracts observed herein could possibly be attributed to those components.
Diosgenin, an aglycone form of dioscin found in the Dioscorea family, attracted significant attention as it was known to have antioxidant, cholesterol-lowering, antiosteoporotic, and anticarcinogenic activities. D. batatas has been reported to contain 13.6 mg dioscin per gram dry powder. Dioscin, which is likely to be less bioactive than its aglycone diosgenin, is suspected to undergo extensive conversion into diosgenin in the large intestine by microbial enzymes, and thereby exert a variety of biological activities. Another candidate compound that may be responsible for improving DSS-mediated colitis includes phenanthraquinone and its derivatives. The compound is also a potential anti-inflammatory agent in D. batatas, since 1,4-phenanthraquinone has been reported to suppress LPS-induced NO production and inflammatory cytokine expression. 41 However, further studies are required to accurately identify the specific anti-inflammatory compounds present in the DBE bark used in our study.
Taken together, our findings show that oral supplementation with DBE bark could enhance the antioxidant activities in colonic epithelial cells and attenuate the inflammatory response in activated macrophages, thereby alleviating acute colitis in DSS-treated mice. As such, D. batatas bark can be considered as a natural supplement for preventing inflammatory diseases of the intestine.
Footnotes
Acknowledgment
This study was funded by Forest Resources Development Institute of Gyeongsanbuk-do, Andong, South Korea (2018).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.