
Abstract
Our study aimed to investigate the effects of the polysaccharide-rich extract of Phragmites rhizoma (PEP) against water immersion restraint (WIR) stress and forced swimming-induced fatigue. Exposure to WIR stress significantly increased the ulcer index, bleeding score, the weight of the adrenal gland, blood glucose concentrations, total cholesterol, cortisol, and creatine kinase (CK). The weight of the spleen decreased significantly. In addition, myeloperoxidase (MPO) and thiobarbituric acid-reactive substance (TBARS) were significantly upregulated by WIR stress. The antioxidative factors such as glutathione (GSH) and superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) in the stomach were decreased by WIR stress. Alterations induced by WIR stress were effectively reversed by pretreatment with PEP. The swimming endurance capacity of mice was significantly prolonged by the oral administration of PEP. Swimming-induced fatigue significantly reduced the body weight; however, the injection of PEP inhibited the decrease of body weight. The PEP-treated group had significantly lower CK levels in plasma, an indicator of muscle damage. These results indicated that PEP has anti-stress and anti-fatigue effects, which are mediated by suppressing the hyperactivation of the hypothalamus–pituitary–adrenal axis, and antagonism of the oxidative damages induced by WIR stress and prolonged swimming times.
Introduction
Stress is a response of the body to change in environmental conditions that requires an adjustment or response. Stress reactions include physical, mental, and emotional response. Although stress is an experience from environment, body, and thoughts in a normal part of life, excessive stress causes disease in various body organs such as the cardiovascular, 1 gastrointestinal, 2 immune, 3,4 metabolic, 5 reproductive, 6 and central nervous system. 7 Moreover, the incidences of stress-derived disorders have increased in recent years. 8
Fatigue is derived from stress and is a kind of physiological warning that alerts an individual about the mental and physical status, providing vital signals either before or after the body has been subjected to harm. Previous studies on numerous excessive and prolonged stress-derived fatigue demonstrated an increase in blood followed by rapid exhaustion, and the lactate remained in the muscles or the blood, resulting in fatigue. 9,10 Strenuous and stressful stimuli cause oxidative injury to the body and tissue damage. 11 Earlier studies have demonstrated that increased oxidative damage in the tissues following physical stress is also associated with lipid peroxidation. 12 –14
Plant-derived components are widely used and have been investigated for the treatment of a number of disorders, especially for their anti-stress and anti-fatigue activities. 15 The initial studies on Eleutherococcus senticosus and Panax ginseng made way for a vast area of research, and a substantial body of research has evaluated the anti-stress properties of plants such as Withania somnifera, 16 Ocimum sanctum, 17 and Bacopa monniera. 18 Similarly, numerous studies have reported the anti-fatigue properties of various natural agents, such as P. ginseng, 19 Trichopus zeylanicus, 20 Morinda officinalis, 21 and garlic, 22 which is a result of their adaptogenic and antioxidative properties.
Polysaccharide-rich extract of Phragmites rhizoma (PEP) possesses antioxidative and free radical scavenging properties, 23 whereas the polysaccharides present in Phragmites communis enhance the activities of antioxidant enzymes in the blood of aged mice. 24 Our previous study revealed that treatment with PEP attenuates atopic dermatitis-like skin lesions, suggesting that PEP might suppress the immunological and inflammatory responses in atopic dermatitis. 25 A number of studies have demonstrated that excessive and prolonged stress directly contribute to pathophysiology of inflammatory diseases. 26 In this study, we therefore explored whether the suppressive effect on inflammation of PEP might play a role in attenuating stress and fatigue in animal models.
Materials and Methods
Experimental animals
Male ICR mice (6 weeks old) and Sprague-Dawley rats (8 weeks old) were purchased from Samtako Bio Korea, Inc., (Kyung gi-do, Korea). Three to four animals were housed per cage at a temperature of 22°C ± 0.5°C and 50% relative humidity, in a 12-h light/12-h dark cycle (7:00 a.m. to 7:00 p.m.), in a controlled environment. The animals were randomly assigned to one of the three groups: (1) the normal group, which was not subjected to water immersion restraint (WIR) stress or forced swimming fatigue; (2) the vehicle group, in which the animals were subjected to WIR stress or forced swimming fatigue; and (3) the PEP-treated group in which the animals were subjected to WIR stress or forced swimming fatigue and were treated with PEP at each experimental dose. They were provided standard laboratory food and were allowed ad libitum access to water. The animals were kept in the laboratory for a week of acclimation. All the experimental procedures and methods of animal care were reviewed and approved by the IACUC (Institutional Animal Care and Use Committee) at Chung-Ang University (2016-00007).
Preparation of PEP
P. rhizoma was purchased from Woori Oriental Medicine Materials Co. (Seoul, Korea). The polysaccharides, which are the main bioactive components, were extracted by hot water decoction. 24,25,27 Briefly, the sliced and dried raw materials of P. rhizoma were blanched at 100°C for 4 h (43.3 g/L of water) and allowed to cool down to room temperature. After cooling, the supernatant was applied to a Sephadex G-150 (2.7 × 72 cm) column and the bound materials were eluted with a linear gradient of NaCl (0.1 M NaCl). The fractions containing carbohydrate were pooled and precipitated thrice with ethanol. The resulting polysaccharide extract was freeze-dried for preparing the PEP powder. The concentration of carbohydrates in the fraction was determined using the phenol–sulfuric acid method. 28 The carbohydrate content of the final product was 85.6%, and contamination by proteins and nucleic acids was negligible. The approximate molecular weight of PEP was found to be 267,000.
Experimental procedure for WIR stress and/or swimming fatigue
We used WIR stress model in rats and/or forced swimming fatigue model in mice to evaluate the effect of PEP on restriction stress and/or exercise fatigue in respective murine model. The WIR stress in rats and the forced swimming test in mice are well-established models according to the methods proposed by Takagi and Okabe 29 and by Matsumoto et al., 30 respectively. In the WIR stress experiment, the rats were more appropriate than mice for further study on tissue assay.
In WIR stress model, the rats were fasted for 12 h, during which they were allowed free access to water, and were subsequently subjected to WIR stress. Briefly, the rats were individually immobilized in a restraint cage and were immersed in water for 4 h in a temperature-controlled water tank (23°C). In the swimming fatigue model, to accustom the mice to swimming, they were trained for a week during the preliminary period, during which they were made to swim for 30 min twice a week, at which time the water was circulated at a flow rate of 6 L/min. The swimming time was measured every other day, for 10 days.
At the end of each experimental period, the mice were anesthetized with ether, and blood was collected by cardiac puncture. The blood was centrifuged at 1200 g for 20 min at 4°C and the plasma was separated. The plasma was used to estimate the levels of glucose, triglycerides, cholesterol, creatine kinase (CK), lactate dehydrogenase (LDH), and cortisol.
Thiobarbituric acid-reactive substances assay
Lipid peroxidation was determined according to the method of Buege and Aust, by measuring the formation of thiobarbituric acid-reactive substances (TBARS) with a spectrophotometer. 31 Stomach tissue samples (100 mg) were added to 10-fold its volume of saline and subsequently homogenized on ice for 30 sec (TME-20DN, TAEMIN, Korea). The homogenates were sonicated four times at 20 kHz for 20 sec on ice. After centrifugation at 12,000 g for 15 min at 4°C, 0.9 mL of trichloroacetic acid (TCA) (8%) was added to 0.3 mL of the supernatant. Following centrifugation at 10,000 g for 5 min at 4°C, 0.25 mL of TBA (1%) was added to 1 mL of the supernatant and the resulting solution was heated at 100°C for 20 min. The tubes were cooled down to room temperature and 2 mL of n-butanol was added to each tube and vortexed for 90 sec. After centrifugation at 3000 g for 5 min at 4°C, 1 mL of the butanol phase was utilized for the TBARS assay, wherein the formation of TBARS was measured by a spectrophotometer at 532 nm (UV-160A, Shimadzu, Japan), using malonaldehyde bis (dimethyl acetal) as the standard. The protein assay was performed according to the Bradford method.
Myeloperoxidase activity assay
The myeloperoxidase (MPO) assay was performed as previously described. 32 The samples of stomach tissue were homogenized in 10-fold of their volume of 0.5% hexadecyltrimethylammonium bromide dissolved in 50 mM potassium phosphate buffer (pH 6.0) in a homogenizer. The homogenates were then centrifuged at 40,000 g for 15 min at 25°C for extraction. Duplicate samples (30 μL) of the resulting supernatant were poured into 96-well microtiter plates. For the assay, 200 μL of a mixture containing 100 μL phosphate-buffered saline, 85 μL of 0.22 M sodium phosphate buffer (pH 5.4), and 15 μL of 0.017% hydrogen peroxide was added to the wells. The reaction was initiated by the addition of 20 μL of 18.4 mM 3,3′,5,5′-tetramethylbenzidine·HCl in 8% aqueous dimethylformamide. The plates were stirred and incubated at 37°C for 3 min and then placed on ice, at which time the reaction was ceased by the addition of 30 μL of 1.46 M sodium acetate (pH 3.0) to each well. The level of MPO activity was evaluated by measuring the absorbance of the samples at 620 nm (OD value), using MPO as the standard.
Glutathione assay
The levels of reduced glutathione (GSH) were determined by using fluorescence-proved o-phthalaldehyde (OPA). 33 The samples of stomach tissue were homogenized in 20 mM HCl, 5 mM diethylenetriaminepentaacetic acid (DTPA), 10 mM ascorbic acid, and 5% TCA. The 666 suspension was centrifuged at 14,000 g, and the supernatant was centrifuged through a 0.45 mm microcentrifuge filter (Millipore Corp., Bedford, MA), which yielded a 5% deproteinized homogenate that was stable at 270°C for at least 4 weeks. The assays consisted of paired samples in 10 × 75 mm borosilicate tubes, labeled A and B. Sample A represented the background, consisting of non-glutathione-dependent fluorescence that was subtracted from the paired sample B. The OPA-derived fluorescence was measured at 365 nm excitation (slit width 5 nm) and 430 nm emission (slit width 20 nm). The following solutions were used for the OPA assay: Redox quenching buffer (RQB) (20 mM HCl, 5 mM DTPA, and 10 mM ascorbic acid); 5% TCA in RQB (TCA–RQB); 7.5 mM N-ethylmaleimide (NEM) in RQB; 1.0 M KPi buffer (pH 7.0); 0.1 M KPi buffer (pH 6.9); and 5.0 mg/mL OPA in methanol. The OPA solution was prepared immediately before use.
Superoxide dismutase activity assay
The superoxide dismutase (SOD) activity was measured by a spectrophotometer with an SOD assay kit using a microwell plate (Fluka, Japan). The samples of stomach tissue were homogenized on ice with 1 mL of 50 mM potassium phosphate buffer (pH 7.4), containing 1 mM ethylenediaminetetraacetic acid (EDTA) per 100 mg of tissue. After centrifugation at 10,000 g for 15 min at 4°C, the supernatants were collected for the assay as sample solution. Briefly, a solution of the water-soluble tetrazolium salt and xanthine was added to each sample. The reaction was subsequently initiated by adding a solution of xanthine oxidase. After incubation at 37°C for 20 min, the absorbance was measured at 450 nm.
Glutathione peroxidase activity assay
The activity of glutathione peroxidase (GPx) was measured according to the method of Lawrence and Burk. 34 Briefly, 1 mL of 50 mM phosphate-buffered saline (PBS) solution (pH 7.4) containing 5 mM EDTA, 2 μM reduced NADPH, 20 μM GPx, 10 μM sodium azide, and 23 mU glutathione reductase was incubated at 37°C for 5 min. Then, 20 μL of 0.25 mM H2O2 solution and 110 μL of the supernatant from the specimen were added to the assay mixture. The change in the absorbance at 340 nm was monitored for 1 min. The change in the absorbance of a blank sample containing all the ingredients, except the supernatant, was also monitored for the same period of time.
Catalase activity assay
The catalase (CAT) activity in the stomach was measured using the method of Aebi. 35 Briefly, H2O2 was used as the substrate and the decrease in the concentration of H2O2 at 20°C in phosphate buffer was observed by measuring the absorbance at 240 nm. One unit of CAT activity was defined as the amount of enzyme that degrades 1 μmol of H2O2/min.
Anti-fatigue determination in mice
Anti-fatigue assay was only done on ICR mice. To determine the anti-fatigue property of the extracts, the swimming capacity of male ICR mice was studied using an adjustable-current swimming pool. The details of this apparatus have been described by Matsumoto et al. 30 We used an acrylic plastic pool (90 × 45 × 45 cm) filled with water to a depth of 38 cm. The surface of the tank was clear and smooth, which prevented the animal from supporting itself while swimming. The current in the pool was generated by a pump, which circulated the water. We devised a spout and suction apparatus to generate a uniform current. Vertical holes of appropriate diameter were bored in the nozzle in a straight line, with precision to the nearest 0.1–0.2 mm. Water was returned to the pump through a narrow slit in the plastic pipe fixed to the bottom of the pool. The strength of the current was adjusted by changing the water flow, which was regulated by opening and closing a valve, and was monitored by a water flowmeter. The temperature of the water was maintained at 34°C, with a water heater and a thermostat.
To accustom the mice to swimming, they were trained for a week during the preliminary period, during which they were made to swim for 30 min twice a week, at which time the water was circulated at a flow rate of 6 L/min. During the preliminary period, the mice were fed on a commercial diet. On the last day of the preliminary period, they were then made to swim until they were fatigued, which was defined as the failure to rise to the surface of the water for breathing within a 7-sec period, when the strength of the current was 8 L/min.
The total period during which the mice could swim before reaching fatigue was measured as the index of swimming capacity. The mice were then divided into three groups, each consisting of 6 mice, and having similar average swimming times: (1) the normal group; (2) the vehicle group; and (3) the PEP-treated group (1 g/kg of body weight, p.o., administered daily until the end of the experiment). The normal and control groups were fed with the vehicle for the same number of treatment days. The swimming time was measured every other day, for 10 days. At the end of the experimental period, the mice were anesthetized with ether and blood was collected by cardiac puncture. The blood was centrifuged at 1200 g for 20 min at 4°C, and the plasma was separated. The plasma was used to estimate the levels of glucose, triglycerides, cholesterol, CK, LDH, and cortisol.
Data analyses
All the analyses were performed using the SPSS/PC statistical program (version 12.0 for Windows; SPSS, Chicago, IL, USA). All the values are expressed as the mean ± SEM. The Student's t-test, ANOVA, and Fischer's exact test were used to determine the statistical significance of data. Differences were considered statistically significant at P < .05.
Results
Determining the optimal exposure time for WIR stress and dose of PEP in rats
To determine the optimal duration for WIR stress exposure to the rats, we evaluated the effects of WIR stress over varying periods of time by scoring the incidences of bleeding and ulceration in the stomach. The bleeding index and ulcer index after WIR stress varied depending on the time of exposure (Fig. 1A, B). Ulcer formation and severe bleeding were observed when the WIR stress was induced for 4 h. Therefore, a 4-h WIR stress model was used for subsequent experimentations.
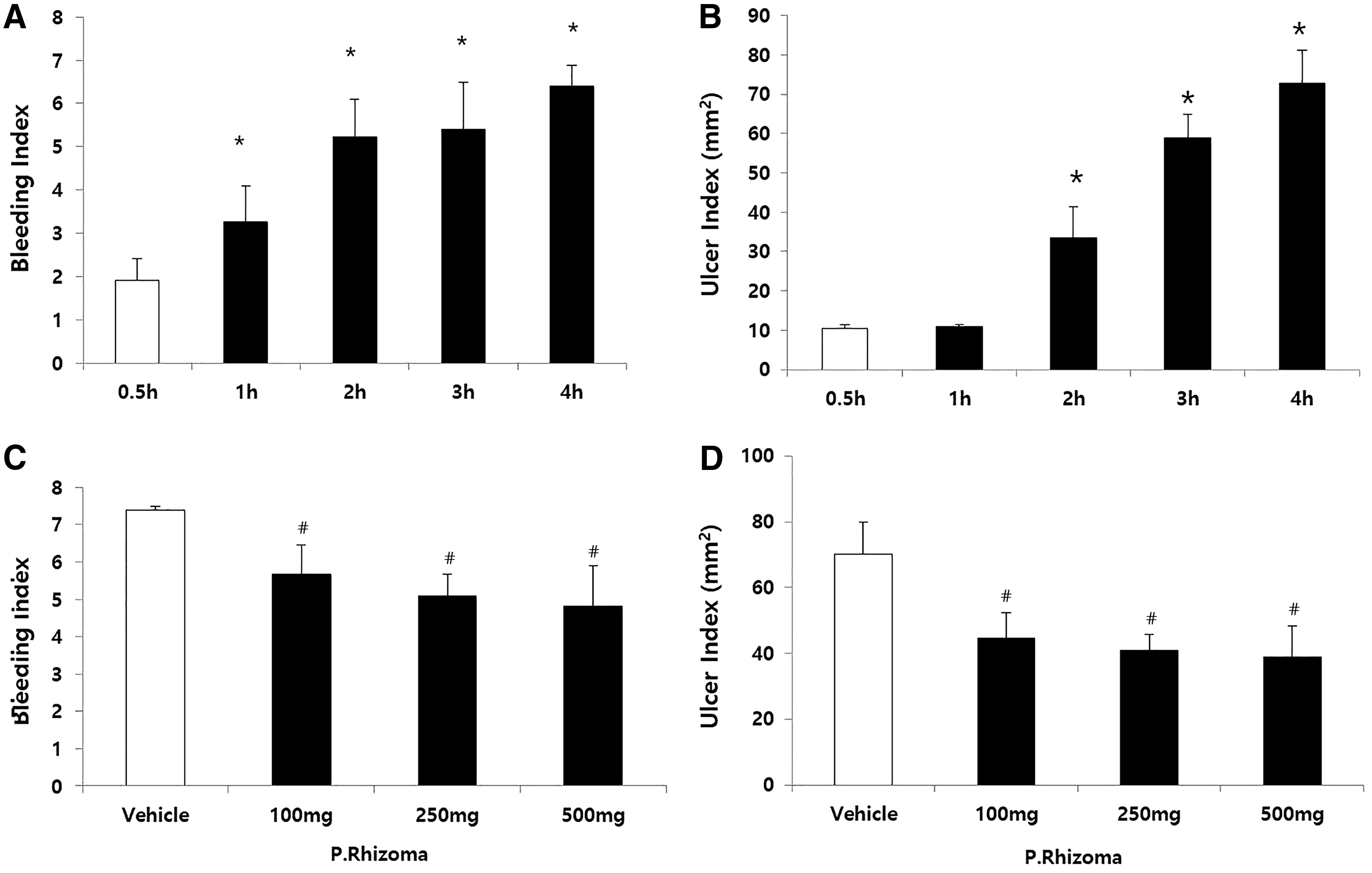
Determination of WIR stress exposure time and PEP dose in rats.
To determine the optimal dose of PEP for the rats, three different doses were tested for each extract. Both bleeding and ulcer formation were significantly inhibited by all the three doses of the extract (Fig. 1C, D). Ulcer formation was predominantly inhibited at a dose of 250 mg/kg in the PEP group. Therefore, 250 mg/kg of PEP was injected by oral gavage to the rats in the subsequent experiments.
Effect of PEP on WIR stress-induced alterations on body and organ weights in rats
Immediately after exposure to WIR stress for 4 h, the body weight (Fig. 2A) and the weight of the dissected organs were measured. The weight of the spleen was significantly reduced by WIR stress (Fig. 2B), in comparison to that of the control. The reduction in the weight of the spleen did not completely recover following treatment with PEP. Exposure to WIR stress significantly increased the weight of the adrenal gland, in comparison to that of the normal group. However, the increase in the weight of the adrenal gland was significantly inhibited following the administration of PEP (Fig. 2C).
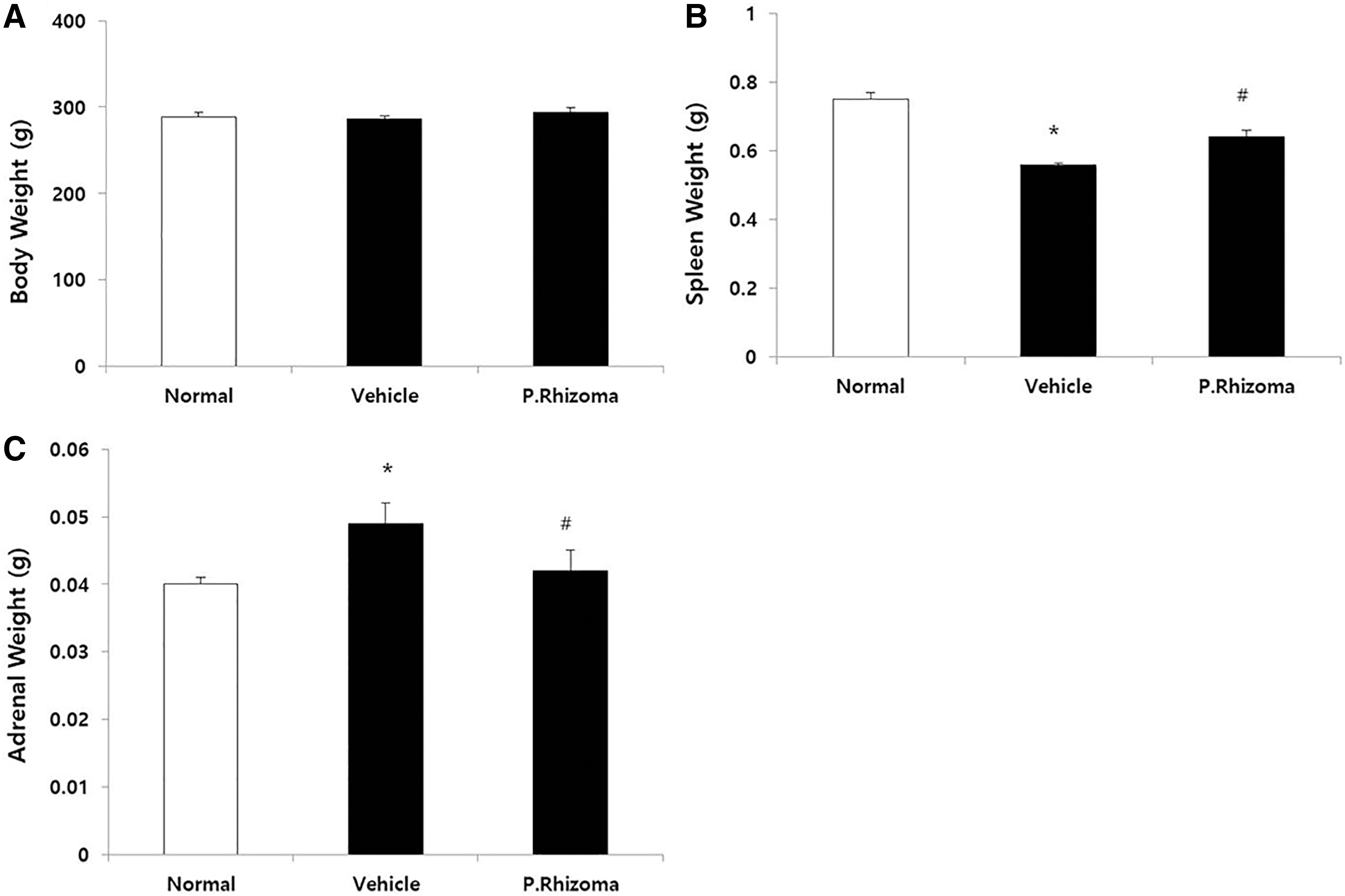
Effects of PEP pretreatment on WIR stress-induced alterations in body and organ weights of rats. Except normal group, all rats were exposed to WIR stress for 4 h.
Effects of PEP on WIR stress-induced biochemical alterations in rats
WIR stress significantly increased the levels of plasma glucose, CK, total cholesterol (T. chol), and cortisol (Table 1). However, their levels after pretreatment with PEP showed no significant change.
Effects of Polysaccharide-Rich Extract of Phragmites rhizoma on Plasma Biochemical Parameters of Water Immersion Restraint Stress
Except normal group, all rats were exposed to WIR stress for 4 h. Data are mean ± SEM (n = 10).
Indicate values that are significant from normal group (* P < .05, ** P < .01).
LDH, lactate dehydrogenase; CK, creatine kinase; TG, triglyceride; T. Chol, total cholesterol; WIR, water immersion restraint; PEP, polysaccharide-rich extract of Phragmites rhizoma.
Effect of PEP on MPO activity and TBARS levels in rats
An increase in the MPO activity and the levels of TBARS was observed in the stomach in the vehicle group, in comparison to those of the normal group (Fig. 3A, B). The increase in the MPO activity and level of TBARS in the gastric mucosa was significantly attenuated in the PEP-treated group.
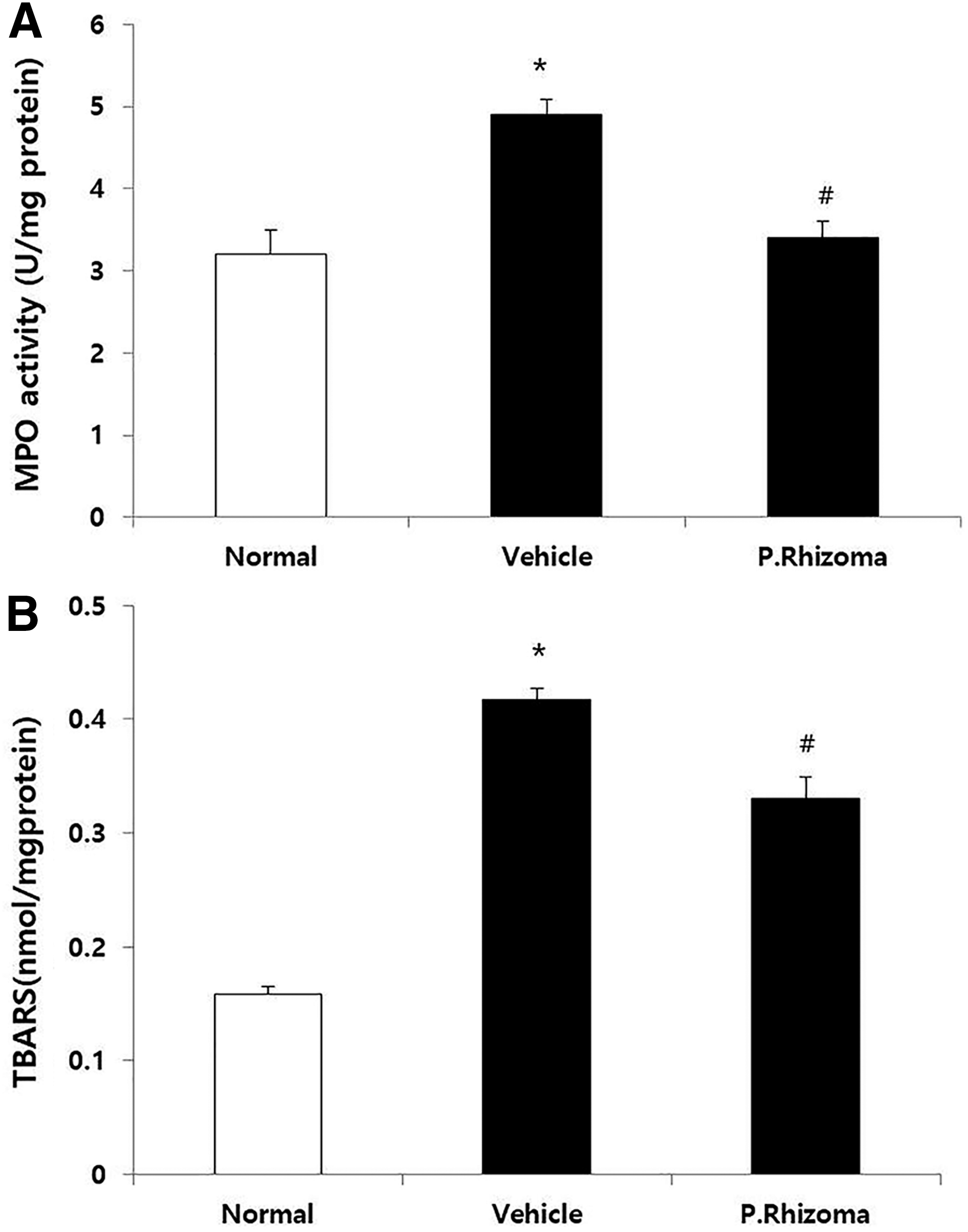
Effects of PEP on MPO activity
Effect of PEP on GSH, SOD, CAT, and GPx activity in rats
The decrease of the activities of SOD and CAT in the stomach was observed in the vehicle group in comparison to those of the normal group (Fig. 4A, B). Conversely, their activities were significantly enhanced in the PEP-treated group. The levels of GSH and GPx activity also followed the same pattern (Fig. 4C, D).
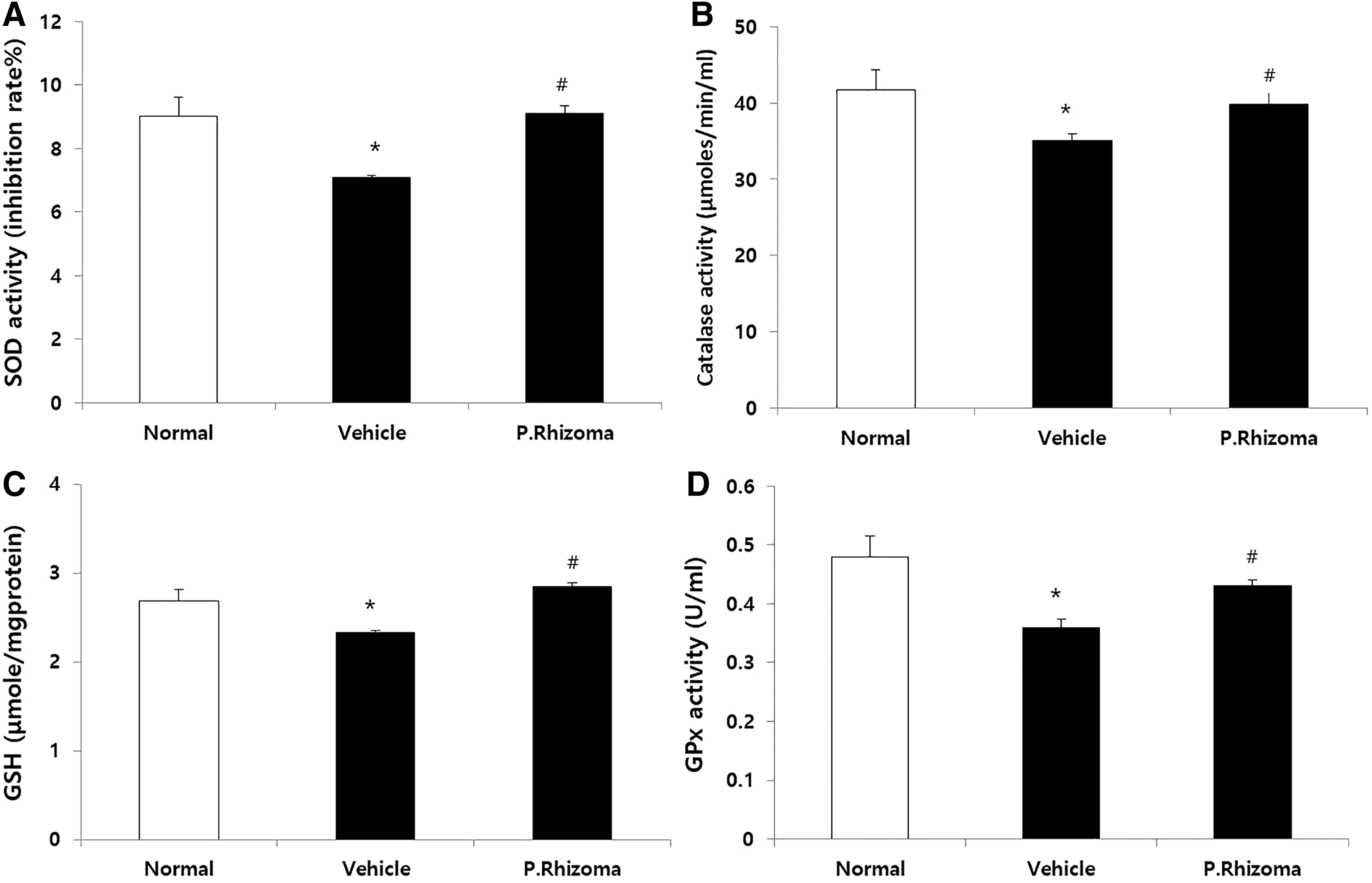
Effects of PEP on SOD
Effects of PEP on the endurance capacity during prolonged swimming and alterations in body weight induced by forced swimming tests in mice
The trained mice were made to swim every alternate day for 10 days, at which time the water flow was kept at 8 L/min, until they were fatigued. The time taken for the mice to reach fatigue was markedly increased in the PEP-treated group (Fig. 5A), in comparison to that of the normal group. The body weight was not changed (Fig. 5B).
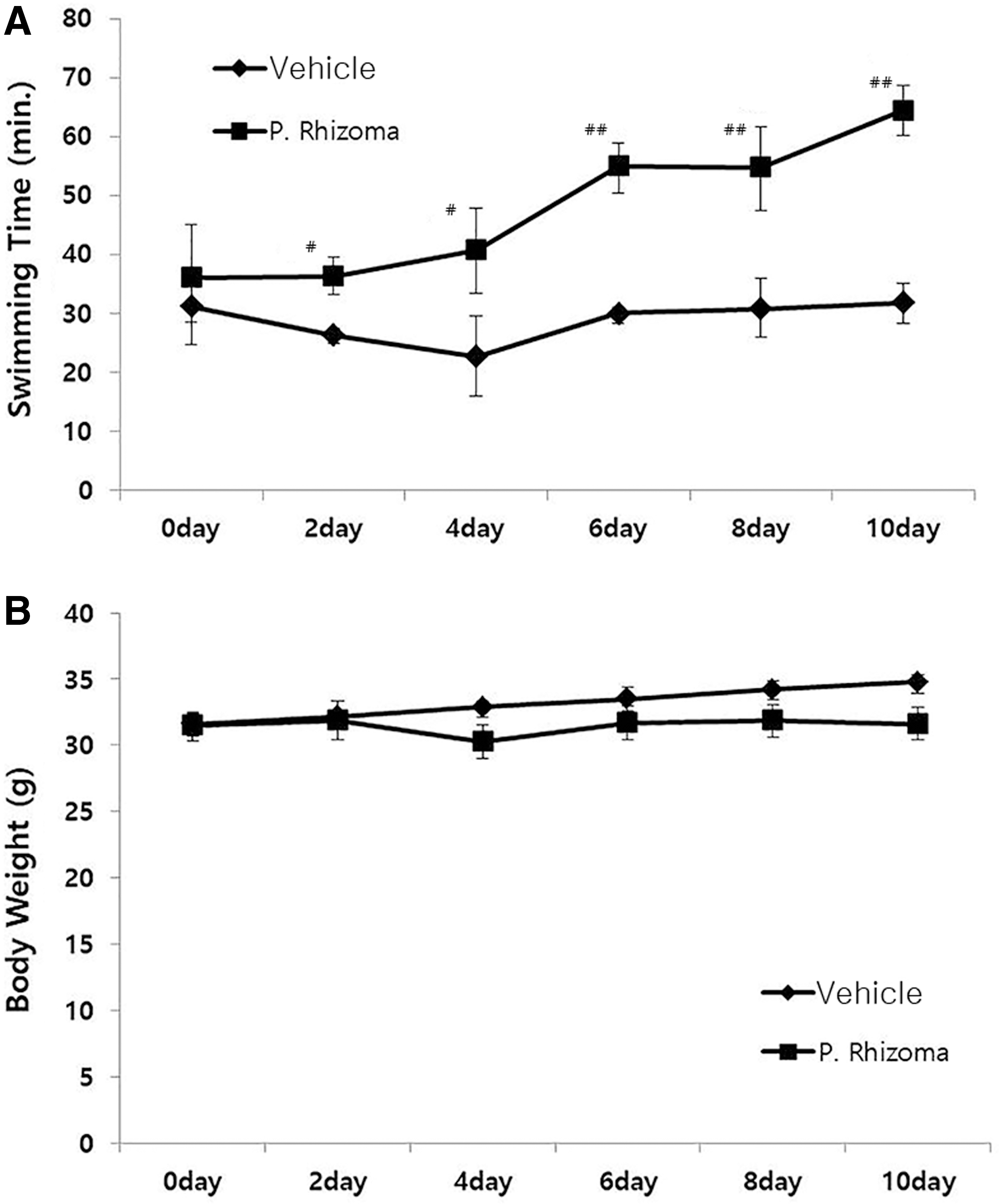
Effects of PEP on swimming time
Effect of PEP on the biochemical alterations induced by forced swimming tests in mice
For each group, the glucose, LDH, and
Effects of Polysaccharide-Rich Extract of Phragmites rhizoma on Plasma Biochemical Parameters of Forced Swimming Test
Normal group did not undergo a forced swimming test. Data are mean ± SEM (n = 6).
and # indicate values that are significant from normal group and vehicle group, respectively (* or # P < .05, ** P < .01).
Discussion
The stress response is a complex system that requires the subsequent involvement of the hypothalamus–pituitary–adrenal (HPA) axis. 36,37 The HPA axis is important for regulating the effects of stressors and for organizing the endocrine, autonomic, and behavioral responses to stress. 38 The metabolisms of glucose, lipids, and protein are affected by the HPA axis. There is considerable scientific evidence to support that stress causes gastric ulcers and release of corticosterone. 29,30,39 Stress results in the formation of ulcers through the hyperactivation of the paraventricular nucleus. Because reduced mucosal blood flow increases gastric contractility, it induces hyperactivation of the paraventricular nucleus. The pathogenesis of ulcers is linked to the imbalance between the destructive and defensive factors. 40
Adrenal hypertrophy induced by stress was observed after WIR stress. The HPA axis highly responding to stress is known to cause adrenal hypertrophy, 41,42 and activates the sympathetic nervous system. Corticosterone and epinephrine release are induced by the activation of the sympathetic nervous system. 43 These hormones were considered necessary responses to counter stress response in the body and aids combating stress. When stressed, the nerve terminals trigger the recruitment of lymphocytes from the spleen to the blood, and is associated with the reduction in the weight observed after exposure to WIR stress. There were a number of plant extracts studied for their rejuvenating as well as adaptogenic or anti-stress properties. 44 In this study, pretreatment with PEP reversed the stress-induced elevation of the ulcer index, adrenal hypertrophy, and spleen atrophy, suggesting that PEP possesses potential anti-stress activities.
The activity of the HPA axis was increased by physical and psychological stressors. It also resulted in oxidative damages by inducing an imbalance in the in vivo status of pro-oxidants and antioxidants. Stress stimulated numerous cell signaling pathways that increased free radicals. Previous studies have shown that oxidative stress production in the brain contributed to stress. Numerous studies have also shown that restraint stress alters the status of the antioxidant, leading to an increase in oxidative stress, which resulted in oxidative damage. 45 –47 It has been described that PEP has antioxidant properties in terms of scavenging reactive oxygen species (ROS) caused by ultraviolet (UV) light. 48 The oxidative stress was induced by excess of ROS. 49 One of the important mechanisms for cellular damage in oxidative stress was lipid peroxidation. This study demonstrated that pretreatment with PEP effectively prevented lipid peroxidation in the gastric mucosa.
The antioxidant defense systems were induced by the generation of ROS, which were increased by a severe stressor such as restraint. SOD and CAT, which are free radical scavenging enzymes, were decreased by WIR stress because of the excessive production of ROS. This weakens the first step of enzymatic defense against superoxide anions and hydrogen peroxide. Clinical studies on patients with affective disorders have also demonstrated lower levels of SOD 50 and CAT. 51 Moreover, GSH and GPx were also observed to have been significantly depleted from the stomach of WIR-stressed rats in this study, which indicated the impairment of the second step of antioxidant defense. This probably further aggravated oxidative damage by negatively affecting the critical GSH-related repair pathways, including free radical scavenging, detoxification of electrophilic compounds, modulation of cellular redox status, and the regulation of cell signaling. The results of this study demonstrated that stress-induced deficiency of antioxidant activities in the stomach was relieved by PEP.
The effects of PEP in fatigue were investigated in mice using a flow rate-adjustable swimming pool. For mice administered the extracts, duration of the swimming gradually increased compared with the control group. On day 10, there was significant improvement in the duration of swimming. In the control group, the swimming time of the mice did not show significant changes. It was demonstrated that the extracts affected the recovery from fatigue induced by intensive exercise.
Numerous studies have demonstrated that oxidative stress plays a crucial role in the etiology of chronic fatigue syndrome and antioxidants might prove useful in reversing oxidative stress. 52 –54 The increase in oxidative stress due to an imbalance between the attack by oxidants, arising from the production of free radicals, and defense by antioxidants, has been well documented in studies involving patients with chronic fatigue syndrome. 52,54,55 In humans, generation of ROS in the muscle was induced by muscular exercise. Previous studies demonstrated that oxidation of proteins promoted by exercise resulted from ROS and ROS was a major contributor to muscle fatigue. 56 Therefore, the mechanisms of the treatment of muscle fatigue might be scavenging ROS. Endogenous cellular defense mechanisms for eliminating ROS existed in the muscle cells. In disease states, such as chronic fatigue, defense mechanisms against oxidative damage became weakened. The exogenous antioxidants from the extracts studied herein may have acted directly. A cooperative network of cellular and exogenous antioxidants was created for protecting against fatigue. These defense mechanisms against oxidative stress became weakened in chronic fatigue and other disease condition.
In this study, the plasma concentration of CK was only affected by treatment with PEP following the forced swimming test. The level of CK in the blood represented the muscle damage following endurance exercise. This enzyme was widely used to indicate muscular damage in previous studies. 57 –59 In this study, PEP significantly reduced the levels of CK in the plasma. It may be considered that the suppression of muscle damage improved the physical stamina and endurance of mice. In addition, the concentrations of TG were decreased by the forced swimming test. It was well known that TG concentrations are affected by numerous factors, including physical activity. The major factors altering TG concentrations during physical activities were the intensity, duration, and frequency of physical activity. Physical activities such as forced swimming might affect the metabolic rate, increasing the resulting need for energy by several folds of the resting rate, and simultaneously trigger the metabolic pathways for oxidizing fats and carbohydrates. 58
The results of this study indicate that the antioxidant potential of PEP might contribute to its therapeutic activity. The observation that pretreatment with PEP ameliorated stress-induced oxidative damages suggested that the protective actions of PEP, as observed during the treatment of numerous stress-related diseases in the clinics, could be mediated by the increase in the in vivo antioxidant status, which could be one of the important mechanisms underlying the activities of PEP.
Footnotes
Acknowledgment
This research was supported by the Chung-Ang University Research Scholarship Grants in 2018 and by the Basic Science Research Program through the National Research Foundation of Korea (NRF) grant funded by the Ministry of Education, Science and Technology [Grant No.: 2016R1D1A1A09918019].
Author Disclosure Statement
The authors declare no conflicts of interest.