
Abstract
The effects of dietary supplementation with aqueous Hovenia dulcis Thunb. extract (HDE) (20 weeks) on high-fat diet (HFD)-induced nonalcoholic fatty liver disease and dyslipidemia were evaluated in mice. Supplementation with 200 and 800 mg/kg feed HDE (HDE200 and HDE800, respectively) resulted in no significant difference in growth in the HFD-fed groups. The triglyceride (TG) levels and free fatty acids were significantly decreased, whereas high-density lipoprotein cholesterol was increased in the HDE800 group (P < .05). The hepatic intracellular TGs were significantly decreased in the HDE-fed groups and lipogenic enzymes (acetyl CoA carboxylase, fatty acid synthase, stearoyl CoA desaturase, and diacylglycerol transferase) in the liver were significantly downregulated by HDE supplementation (P < .05). The diminished serum antioxidant enzyme activities in the HFD group were effectively restored by HDE supplementation, which also contributed to the attenuation of hyperlipidemia.
Introduction
Nonalcoholic fatty liver disease (NAFLD) is defined as elevated hepatic triglyceride (TG) accumulation (>5% fat by weight) and it is associated with hypertension, atherosclerosis, diabetes, and dyslipidemia. 1,2 NAFLD can progress to serious liver alterations, which is known as nonalcoholic steatohepatitis, by continuous oxidative stress and inflammation. 3 Steatosis is now increasing worldwide because of the increased prevalence of obesity. 4 Although abnormal liver functions are the primary causes of NAFLD, the precise NAFLD mechanisms remain unknown. 5
Dyslipidemia is characterized by abnormal TG levels, reduced high-density lipoprotein-cholesterol (HDL-C), and excessive low-density lipoprotein-cholesterol (LDL-C) in the blood. 1 The occurrence of dyslipidemia has increased with cardiovascular diseases worldwide. 6,7 Severe hypertriglycedemia is associated with the onset of atherosclerosis exclusive of hypercholesterolemia. 8 The prevalence of hyper-LDL-cholesterolemia is cardiovascular disease, 9 and that reduced HDL-C is also a high-risk factor of cardiovascular diseases. 10 Furthermore, chronically elevated free fatty acid (FFA) is associated with impaired insulin signaling pathways. 11 Taken together, dyslipidemia is recognized as an imperative risk factor for cardiovascular disease.
Diet, exercise, and lipid-lowering agents can improve NAFLD and dyslipidemia. 12 The fruit of Hovenia dulcis Thunb. (HD, Oriental Raisin Tree) is a widely used traditional medicine in Asian countries, including Korea, Japan, and China. The aqueous extract of HD has been used as a functional ingredient for hepatoprotective health in Korea based on the evidence of improved alcohol detoxification 13 ; however, the effect of HD aqueous extract (HDE) on NAFLD has not been explored. This study was conducted to evaluate the effects of HDE supplementation on high-fat diet (HFD)-induced NAFLD and dyslipidemia in mice.
Materials and Methods
HDE preparation
Concentrated aqueous HDE was kindly provided by Kwang-Dong Pharmaceutical Co. Ltd. (Seoul, Korea). The powdered sample was obtained by freeze drying the concentrated aqueous HDE. Sugars in the sample were partially removed using Diaion HP20 resin (Mitsubishi Chemical, Tokyo, Japan) column chromatography (2.5 × 300 mm) and washing with distilled water. Adsorbed compounds in the resin were recovered by eluting with acetone. The powdered sample was prepared by removing solvent. The total phenolic and flavonoid contents in the samples were 87 mg gallic acid equivalent/g powder, and 12 mg catechin equivalent/g powder, respectively. The phenolic content in the recovered sample was increased by 2.5 times compared with that in the initial aqueous HDE, and the partially desugared HDE was used for the animal study.
Animals and diets
C57BL/6 male mice (4 weeks old) were obtained from Dae-han Bio Link (Chung-Buk, Korea) and were housed at 22°C ± 1°C and 50% ± 10% relative humidity with a 12-h light:dark cycle (8:00 a.m. to 8:00 p.m.). All animal experiments abided by the protocols for animal studies as permitted by IACUC at Kookmin University (KMU-2016-8). After adaptation to a normal chow diet for 1 week, mice were randomly assigned into five groups (12 mice in each group) as follows: (1) normal diet group (ND, 18% kcal from fat), (2) control group (CTL, HFD, 60% kcal from fat) (Research Diets, Inc., New Brunswick, NJ, USA), (3) HDE200 group (HDE200, HFD +200 mg/kg HDE feed), (4) HDE800 group (HDE800, HFD +800 mg/kg HDE feed), and (5) fenofibrate group as a positive control (FF, HFD +500 mg/kg FF feed) (Sigma, Ronkonkoma, USA). Mice had free access to food and water for 20 weeks. Body weight and feed intake were measured weekly. The concentration of the experimental dosage was determined based on a previous study that the oral administration of HDE (200–1000 mg/kg bw, mice) was able to rescue alcohol-induced liver injury, hangover, and acute hyperlipidemia. 14
Body composition measurement
Mice were fasted for 4 h and then anesthetized by intraperitoneal injection of ketamine (100 mg/kg bw) and xylazine (10 mg/kg bw) before measuring body composition. Lean body mass and body fat mass were measured at 4, 8, 12, 16, and 20 weeks by dual energy X-ray absorptiometry (DEXA) (Medikors, Gyeong-Gi, Korea).
Blood biochemical analysis
Blood was collected from the retro-orbital cavity after anesthesia. Serum was separated by centrifugation at 2000 g for 15 min at 4°C and was stored at −80°C until further analysis. Serum TG, FFA, total cholesterol (TC), LDL-C, and HDL-C levels were measured using the appropriate assay kits (Wako Chemicals, Tokyo, Japan). Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels in the serum were also determined using assay kits (Asan Pharm, Seoul, Korea).
Antioxidant enzyme activities
Superoxide dismutase (SOD), glutathione reductase (GR), and glutathione peroxidase (GSH-Px) levels in the serum were determined according to the manufacturer's protocol (Cayman, Ann Arbor, MI, USA) after appropriate dilution with distilled water. Catalase activity was determined using the method of Peng et al. 15
Hepatic histology and liver lipid content
Liver tissues were frozen in optimal cutting temperature compound (Sakura Finetek, Torrance, USA) at −80°C. Cryostat 4-μm sections were examined using a light microscope at 100 × scale for Oil Red O staining. Total liver lipids were extracted using 5% NP-40 and quantified using a commercial kit (Wako Chemicals). Liver proteins were determined using the bicinchoninic acid method. 16
Immunoblotting analysis
Liver tissue proteins were extracted using RIPA buffer (CellNest, Tokyo, Japan), containing a phosphatase inhibitor (Cell Signaling, Danvers, MA, USA) and a protease inhibitor (Cell Signaling) for 50 min at 4°C. After centrifugation at 12,000 g for 10 min at 4°C, the protein concentration in the supernatants was determined using the Bradford assay. Equal quantities of proteins were separated by SDS-PAGE (20%) and the separated proteins were then transferred to a polyvinylidene difluoride membrane. The membranes were blocked in 5% bovine serum albumin (GenDEPOT, Houston, TX, USA) for 1 h at room temperature and immuno blotted using primary antibodies. The primary antibodies were as follows: acetyl CoA carboxylase (ACC), fatty acid synthase (FAS), stearoyl CoA desaturase 1 (SCD1), β-actin (Cell signaling), diacylglycerol acyltransferase (DGAT) (BioVision, Milpitas, USA), and sterol regulatory element-binding protein 1 (SREBP1) (SantaCruz, Dallas, TX, USA). Suitable horseradish peroxidase-conjugated antirabbit IgG (Bio-Rad) secondary antibodies were applied at a dilution of 1:3000. Protein bands were observed by enhanced chemiluminescence (Bio-Rad) using a molecular imager chemotic XRS+ (Bio-Rad). Band intensity was quantified by Image Lab (Bio-Rad). Expressions of de novo lipogenesis proteins including ACC, FAS, SCD1, DGAT, and SREBP1 were normalized as ratios to β-actin.
Statistical analysis
Data are presented as mean ± standard error of the mean. Statistical analyses were performed using Prism 6 GraghPad (La Jolla, CA, USA). The results were compared by one-way ANOVA followed by the Dunnett multiple comparisons test. P values ≤.05 were considered statistically significant.
Results and Discussion
Effect of HDE supplementation on growth performance
No adverse effects were found in any of the mice during the experimental period. The effects of HDE on growth performance comprising body weight, feed intake, fat mass, and lean mass are shown in Figure 1. HFD-fed groups including CTL, HDEs, and FF had higher body weight than the ND group. Body weight in the FF group began to diverge after 8 weeks and remained significantly lower until 20 weeks (P < .05) (Fig. 1A). There was no difference in body weight between the CTL and HDEs groups. Feed intake in the FF group was greater than that of the CTL and HDE groups (P < .05); however, no difference was found in feed intake between the CTL and HDEs groups (P < .05) (Fig. 1B). Other animal studies with HDE supplementation showed similar results for body weight and food intake. 17,18

Effect of HDE supplementation on
There were no previous reports regarding the effects of HDE supplementation on body composition using DEXA. Body fat mass in the FF group significantly decreased compared with that in the CTL group after 8 weeks, and the effect remained significant until 20 weeks (P < .001). No difference was shown between the HDEs and CTL group during a 20-week period (Fig. 1C). Lean mass analysis showed lower mean mass in the FF group than in the CTL group; however, there was no difference between the CTL and HDEs groups (Fig. 1D). Taken together, dietary HDE supplementation did not affect body weight, food intake, fat mass, or lean mass.
Effect of HDE supplementation on serum biochemical markers
The serum levels of TC, LDL-C, and HDL-C after HDE supplementation are presented in Table 1. The levels of TC and LDL-C were not significantly different between the CTL and HDEs groups. Serum HDL-C levels in HDE800 group were significantly increased compared with those in the CTL group (P < .01). The increased HDL-C had a positive effect on cardiovascular diseases. 10 Serum TG levels are an important predictor of the risk of cardiovascular diseases. 19 Increased FFA levels induce mitochondrial dysfunction, development of lipotoxicity, and an impaired insulin signaling pathway. 11,20 Serum TG levels in the HDE200 and HDE800 groups were significantly reduced by 36% compared with those in the CTL group (P < .001), and serum FFA levels in the HDE800 group were decreased by 18% compared with those in the CTL group (P < .05).
Effect of Hovenia dulcis Thunb. Extract Supplementation on Serum Biochemical Markers
Data are presented as mean ± standard error of the mean (n = 12 per each group).
ND, normal diet; CTL, high-fat diet (HFD); HDE200, HFD +200 mg/kg HDE feed; HDE800, HFD +800 mg/kg HDE feed; FF, HFD +500 mg/kg fenofibrate feed.
P < .05; ** P < .01; *** P < .001, versus CTL.
TC, total cholesterol; LDL-C, low-density lipoprotein cholesterol; HDL-C, high-density lipoprotein cholesterol; TG, triglyceride; FFA, free fatty acid; ALT, alanine aminotransferase; AST, aspartate aminotransferase.
Biologically active compounds in HDE including saponin derivatives (hodulosides I–V), phenolic acid (vanilic acid, ferulic acid, etc.), and flavonoids such as myricetin, dihydromyricetin (ampleopsin), quercetin, and taxifolin were identified. 21,22 Gallic acid is one of the major phenolic acids found in vegetable and fruits, including HDE. Chao et al. demonstrated that supplementation with 50 and 100 mg/kg per day gallic acid for 16 weeks significantly reduced HFD-induced hepatosteatosis, and this effect is partly ascribed due to the restoration of HFD-induced disturbance of lipid and glucose metabolism based on a metabolite profile analysis. 23 Guo et al. also reported that supplementation with myricetin extracted from HD significantly decreased serum TG levels and increased HDL-C levels. 24 Based on these results, dietary supplementation with HDE prevented dyslipidemia by improving HDL-C, TG, and FFA.
Increased levels of AST and ALT are the important biomarkers for liver damage and inflammation. 25 The levels of AST and ALT in serum are given in Table 1. Dietary supplementation with HDE revealed that there was no significant difference in AST and ALT levels among the groups. The mild increase in AST and ALT levels in the HDE supplemented groups also did not exceed the reference range in mice (∼67.95 and ∼51.05 U/L, respectively). 26 Based on these results, dietary supplementation with HDE did not alter the enzyme levels tested for liver function.
Effect of HDE supplementation on hepatic lipid accumulation
Liver fat content can be changed by FFA flux through lipolysis, fatty acid oxidation, de novo lipogenesis, and very low-density lipoprotein secretion. 20 Hepatic fat accumulation is associated with insulin resistance without obesity. 27 Accumulation of lipid droplets in the liver tissue was evaluated by Oil Red O staining and quantification of intracellular TG content (Fig. 2). HFD-induced hepatic lipid accumulation, an indicator of steatosis, was increased in the CTL group. The hepatic intracellular TGs were significantly decreased in the HDE200 and HDE800 groups in a dose-dependent manner (P < .05 and P < .001, respectively) (Fig. 2B). It was reported that dihydromyricetin, a major flavonoid found in HDE, significantly reduced oleic acid-induced hepatic steatosis in HepG2 cells by reducing cellular TGs, cholesterol, and malonaldehyde. 28 The administration of 400 and 800 mg/kg bw myricetin derived from HD significantly improved liver damage in high choline-fed mice. Taken together, dietary supplementation with HDE showed health benefit without toxicity by decreasing hepatic lipid accumulation.
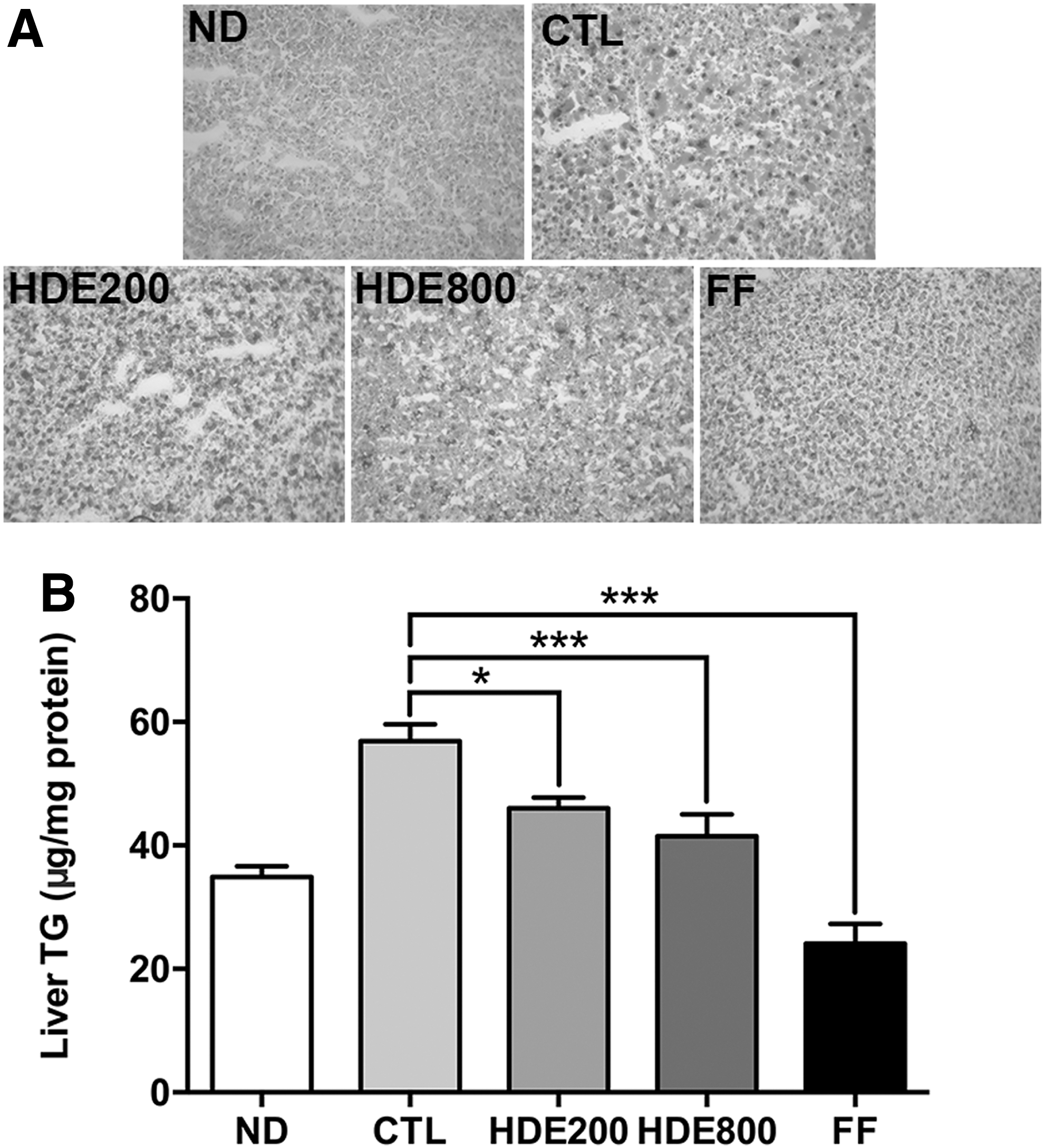
Effect of HDE supplementation on
Effect of HDE supplementation on hepatic de novo lipogenesis enzymes expression
To explore the mechanisms underlying reduced TGs and FFAs in serum, the protein expression levels of hepatic de novo lipogenesis enzymes in the CTL and HDEs groups were compared. As shown in Figure 3, the levels of ACC, FAS, SCD1, and DGAT were consistently lower in the HDE groups in a dose-dependent manner than in the CTL group (P < .05). These proteins are regulated by SREBP1. 29,30 Accordingly, the expression levels of the mature SREBP1 were significantly diminished by 57% in the HDE800 group compared with those in the CTL group (P < .05).
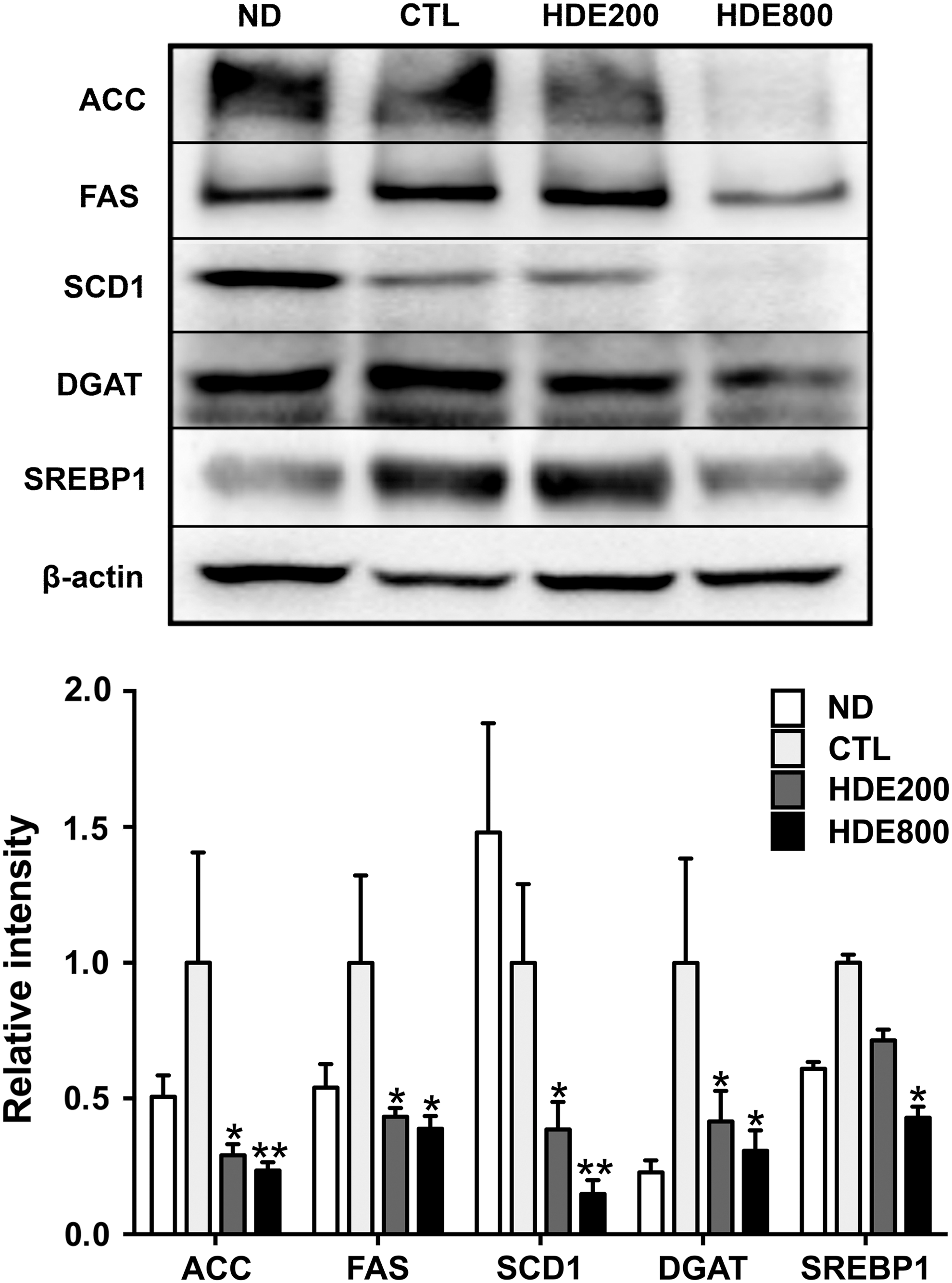
Effect of HDE supplementation on the expression of hepatic de novo lipogenesis enzymes. ND, normal diet; CTL, high-fat diet (HFD); HDE200, HFD +200 mg/kg HDE feed; HDE800, HFD +800 mg/kg HDE feed; FF, HFD +500 mg/kg fenofibrate feed. ACC, acetyl CoA carboxylase; FAS, fatty acid synthase; SCD1, stearoyl CoA desaturase 1; DGAT, diacylglycerol transferase; SREBP1, sterol regulatory element binding protein 1. *P < .05; **P < .01, versus CTL. Data are presented as mean ± SEM (n = 6 per each group).
Dihydromyricetin, a component of HDE, reduced lipid accumulation in two different hepatoma cells such as HepG2 (carcinoma) and LO2 (normal), and this effect was associated with peroxisome proliferator-activated receptor gamma (PPARγ), AMP kinase (AMPK), and phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) signaling pathways. 28 Kim et al. also reported that HDE effectively decreased adipogenic genes, including aP2, resistin, and adiponectin in 3T3-L1 adipocytes, and this antiadipogenic effect was modulated by the activation of AMPK, a key regulator of energy homeostasis. 31
Effect HDE supplementation on antioxidant activity
It has been reported that HFD increases oxidative stress in the liver and adipose tissues and triggers the onset of insulin resistance. 32 The increased oxidative stress is most likely associated with an increase in the mitochondrial oxidation rate, causing an increase in the generation of reactive oxygen species (ROS). Increased ROS production, in turn, elevates FFA in the plasma and liver and results in mitochondrial dysfunction. 33
The activities of antioxidant enzymes including SOD, catalase, GR, and GSH-Px in the HDE800 group were significantly higher than those in the CTL group (Fig. 4). All antioxidant enzyme activities were decreased in the CTL group compared with those in the ND group, but the activities of SOD and GSH-Px were more severely decreased. This result indicates that supplementation with HDE effectively reduces oxidative damage by restoring normal antioxidant enzyme activities in the serum from the HFD-induced increased oxidative stress. Increased superoxide radicals were effectively reduced to H2O2 by increased serum SOD activity and finally detoxified to water by the action of GSH-Px and catalase in rats fed a HFD. 34 Although GR does not interact with ROS directly, it does regenerate oxidized glutathione using NADPH. 35 The decreased activities of serum antioxidant enzymes led to increased malonaldehyde production, reflecting lipid peroxidation in rats fed a HFD. 36 Maieves et al. examined the effect of maturation stage on the antioxidant activity of HDE. 37 They suggested that the immature pseudofruit was a better antioxidant source than the mature fruit based on total phenolic content and antioxidant activity.
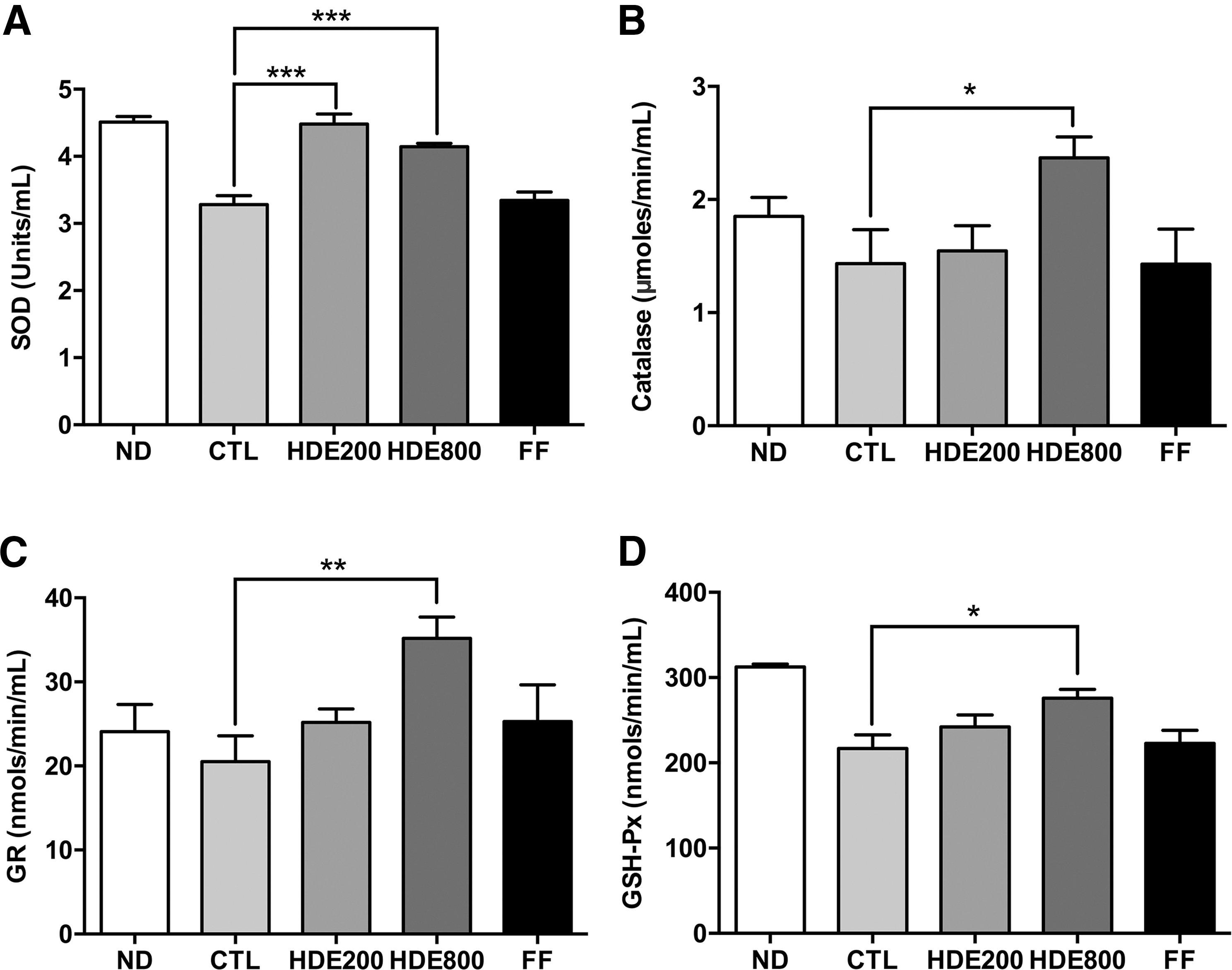
Effect of HDE supplementation on serum antioxidant enzyme activities.
This study demonstrated that dietary supplementation with HDE prevented the development of NAFLD and dyslipidemia induced by HFD in C57BL/6 mice. The diminished activity of antioxidant enzymes in the HFD group was effectively restored by HDE supplementation and possibly contributed to the attenuation of hyperlipidemia. However, whether HDE supplementation could improve the detailed progression of NAFLD-related liver fibrosis must be further investigated.
Footnotes
Acknowledgment
This study was supported by funding from Kwang-Dong Pharmaceutical Co. Ltd.
Author Disclosure Statement
This research was sponsored by Kwang-Dong Pharmaceutical Co. Ltd. and may lead to the development of products.