
Abstract
Perilla oil has been shown to be beneficial for ameliorating metabolic disorders, but its protective effect is still controversial. We investigated the effect of perilla oil on obesity-induced hepatic and vascular changes in high-fat diet (HFD)-fed mice and provided underlying mechanisms for potential therapeutic applications. Tomato and paprika extract was added to prevent the oxidation during storage of perilla oil. HFD-fed mice were orally administered palm or perilla oil for 90 days. Food intake, body and liver weight, and serum cholesterol levels were measured. Arterial and hepatic lipid accumulation was determined by histological staining. Hepatic triglyceride levels and the expression of proteins regulating lipid metabolism were analyzed. Food intake and body weight were not different between palm oil-treated and perilla oil-treated mice. Serum cholesterol level was significantly lower in perilla oil-treated mice compared with palm oil-treated mice. HFD-induced lipid accumulation was also lower in thoracic aorta and liver by perilla oil compared with palm oil. Perilla oil also decreased hepatic triglyceride level without changing the liver weight. Perilla oil treatment increased the AMP-activated protein kinase and acetyl-CoA carboxylase phosphorylation and the lipolytic protein levels, whereas it decreased the lipogenic protein levels in the liver. In conclusion, perilla oil reduced serum cholesterol and arterial and hepatic lipid accumulation in HFD-fed mice. The data suggest that perilla oil improves the balance of lipogenic and lipolytic protein expression, and ameliorates obesity-induced metabolic disorders and cardiovascular diseases.
Introduction
Obesity is a major cause of metabolic disorders, especially type 2 diabetes, cardiovascular disease, hypertension, and nonalcoholic fatty liver disease (NAFLD). 1 NAFLD is a condition of excessive fat accumulation in the liver, known as hepatic steatosis. The rate of hepatic fatty acid uptake and de novo synthesis exceeds the rate of fatty acid oxidation and export, therefore, the excessive fat accumulates. Interestingly, abnormal lipid metabolism and inflammatory responses are also known to cause insulin resistance, dyslipidemia, and other cardiovascular disorders associated with NAFLD. 2 The prevalence of NAFLD is 80–90% in obese adults, 30–50% in patients with diabetes, and up to 90% in patients with hyperlipidemia. 3 Obesity-induced NAFLD patients have abnormal levels of cholesterol and triglycerides from diet or the lipogenic pathway, and exhibit a chronic build-up of inflammatory and stress proteins in circulating blood and endothelial cells. It is known to induce atherosclerotic plaque formation and increase the risk of cardiovascular disorders. 4 In fact, individuals with NAFLD are at a significantly higher risk for cardiovascular disease. 5
Perilla oil, also known as deulgireum, is derived from the seeds of perilla frutescens. It has a unique nutty taste and is used as an antisteatotic herbal medicinal oil in Korea and many Asian countries. 6 It contains both saturated and unsaturated fatty acids, particularly abundant in α-linolenic acid (ALA). 7 ALA is an omega-3 polyunsaturated fatty acid found in many plant seeds, and must be acquired from the diet as an essential fatty acid. Mammals use ALA as a precursor for eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which are also available from fish oil, and are important for proper fetal development, cardiovascular and cognitive function, and lipid metabolic processes. 8,9 However, the intake of omega-3 fatty acids from fish oil should be cautioned for its potential adverse effects from heavy metals, particularly in pregnant women and children. 10 Thus, perilla oil that contains a high proportion of ALA (∼60%) may be a healthier alternative to other sources of omega-3 fatty acids as a dietary supplement. Perilla oil showed similar protective effects as fish oil on high-fat diet (HFD)-induced NAFLD, 11 and was also reported to reduce hypercholesterolemia, and facilitate cholesterol secretion in high fat (HF) and/or high-cholesterol (HC) diet in rats. 12 However, it induced a significant insulin resistance despite lowering serum lipids and increasing hepatic fatty lipid oxidation in rats. 13 In addition, perilla oil inhibited platelet aggregation and thrombus formation to improve blood flow and showed antiatherosclerotic effects in HC diets in rabbits. 6,14 Although it has beneficial effects, perilla oil may have an adverse effect at a specific dose range for a therapeutic purpose but not manifest at the dose of a dietary supplement.
We investigated the effect of perilla oil at a human equivalent dose of 1000 or 5000 mg/day/60 kg on reducing arterial and hepatic lipid accumulation and associated changes in lipid metabolic processes, compared with palm oil, a vegetable oil that has a high concentration of saturated fatty acid, especially palmitic acid (∼45%). 15 Understanding the mechanisms of action and dose responses, perilla oil in the pathology of lipid metabolism will provide important insights for attenuating disease symptoms associated with obesity and excessive fat consumption.
Materials and Methods
Determination of the acid and peroxide value of oil
The acid value (AV) was determined using the official method of AOAC Cd-3a-63. It is the number of milligrams of potassium hydroxide necessary to neutralize the free acids in 1 g of test sample. Five gram was weighed with an accuracy of (±0.01 g) into a 150-mL Erlenmeyer flask. Then, a neutralized solvent mixture of absolute ethanol and ether with the volume ratio of 1:2 was used as the solvents, and it was delivered into a 150-mL Erlenmeyer flask. Then two to three drops of phenolphthalein indicator solution in 1% of isopropyl alcohol were added to the content in the 150-mL Erlenmeyer flask. Finally, 0.1 mol/L of standard potassium hydroxide solution was used to titrate the sample while vigorously shaking to attain a pink color, which should persist for 30 sec; a control test was also performed. Then, the AV was calculated by the following equation:
where V 1 and V 0 are the volumes in ml of standard alkali used in titrating the sample and the blank respectively, and C is the normality of standard alkali, m is the grams of the sample. Each sample was measured three times and then averaged as the final chemical value.
Peroxide values (PV) were determined according to the official method of AOAC Cd-3a-63 with some modifications. Two grams of each oil sample was placed in a flask and dissolved with 10 mL of chloroform. Fifteen milliliters of glacial acetic acid and 1 mL of saturated solution of potassium iodide (KI) were added into the flask. After shaking for 1 min, the flask was closed and placed in a dark place for 5 min. Fifteen milliliters of distilled water was added and the mixture was titrated against 0.002 M sodium thiosulfate solution with starch solution as the indicator. A blank was also titrated under the same conditions. PV was determined using the following equation:
where V represented the volume of sodium thiosulfate used by the sample, and V
0 was the volume (mL) of sodium thiosulfate used in the blank,
Animals, diets, and treatments
Male C57BL/6 mice (4-week-old) were purchased from KOATECH Co. (Pyeongtaek, South Korea) and maintained in the animal facility at Gyeongsang National University. All animal experiments were approved by the Institutional Board of Research at Gyeongsang National University and were performed in accordance with the National Institutes of Health guidelines for laboratory animal care. Mice were housed with an alternating 12-h light/12-h dark cycle and were provided with water and standard chow ad libitum.
Mice were habituated for 1 week and then randomly divided into five groups (n = 8 per group). All mice were fed with a HFD (45 kcal% fat, Research Diets, Inc., New Brunswick, NJ) and administered orally with palm oil (200 or 1000 mg/kg/day), perilla oil (200 or 1000 mg/kg/day), or none of them for 90 days. Perilla oil was extracted by an expeller press machine (National Eng Co., Gyeonggi, Korea) at room temperature, and added with 0.5% of tomato and paprika extract. Then the mixture was treated by supercritical carbon dioxide (SC-CO2) at the pressure of 40 MPa at 50°C for 3 h with flow rate of 20 kg CO2/h. The SC-CO2 extracts of perilla oil with tomato and paprika extracts retarded lipid oxidation. Then, the SC-CO2 extracts were transferred into an air-tight glass container, wrapped with aluminum foil, and refrigerated before feeding the mice. The whole extraction process was performed by S&T FOODS Co. Ltd. (Jinju, Korea). The palm oil was purchased from a local market (Jinju, Korea).
Oil red O staining and hematoxylin and eosin staining
Aorta tissue was fixed in 10% formalin for 24 h and frozen. The frozen tissue was sectioned at 9 μm, and fixed in 10% formalin for 10 min. The sections were washed in distilled water and air dried. Then the sections were incubated in 0.5% Oil Red O working solution (Sigma-Aldrich, St. Louis, MO) for 5 min, rinsed with distilled water, and counterstained with Mayer's Hematoxylin (Sigma) for 1 min. The stained sections were mounted with glycerol gelatin (Sigma) and analyzed using a CKX41 light microscopy (Olympus, Tokyo, Japan).
Liver tissue was fixed in 10% formalin for 24 h. The tissue was sectioned at 5 μm and stained with Hematoxylin and Eosin (Sigma) and analyzed using a CKX41 light microscopy (Olympus).
Serum cholesterol and hepatic triglyceride measurement
The blood samples were collected transcardially or from mouse retroorbital sinus. After centrifugation, the serum cholesterol levels were determined by enzymatic colorimetric assays using a ChemiLab T-CHO Assay Kit (IVCLab, Uiwang, South Korea). Hepatic triglyceride levels were measured using a Triglyceride Colorimetric Assay Kit (Cayman, Ann Arbor, MI).
Western blot analysis
Liver tissue was homogenized in ice-cold RIPA buffer with protease inhibitors (Sigma), sonicated, and incubated for 20 min on ice. After centrifugation, the supernatant was transferred to a clean tube and protein concentration was determined using the Bio-Rad Protein Assay Kit (Bio-Rad, Hercules, CA, USA). The protein lysates were separated through sodium dodecyl sulfate–polyacrylamide gel electrophoresis followed by electrophoretic transfer onto a polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). The membranes were incubated with primary antibodies (1:1000 dilution) against phosphorylation of AMPK (p-AMPK), AMP-activated protein kinase (AMPK), fatty acid synthase (FAS), phosphorylation of ACC (p-ACC), acetyl-CoA carboxylase (ACC), stearoyl-CoA desaturase 1 (SCD1) (Cell Signaling Technology, Beverly, MA), adipose triglyceride lipase (ATGL), CD36 (Abcam, Cambridge, MA), and horseradish peroxidase-conjugated secondary antibody, and subjected to ECL substrates (Bio-Rad Laboratories, Hercules, CA). The ChemiDoc XRS+ System (Bio-Rad Laboratories) was used to analyze the densitometry of protein bands.
Statistical analysis
Statistical differences among the groups were determined with one-way analysis of variance, followed by Bonferroni post hoc analysis. Student's t-test was used when only two groups were compared. Values were expressed as the mean ± standard error of the mean. A P value < .05 was considered statistically significant.
Results
Effect of tomato and paprika extract on perilla oil oxidation
To examine the effect of perilla oil on obesity, perilla oil was prepared and characterized. The fatty acid compositions of palm oil and perilla oil were compared (Table 1). Saturated fatty acids, C16:0 (palmitic acid) and C18:0 (stearic acid) were 36.7% and 6.6% in palm oil versus 5.8% and 0% in perilla oil, respectively. Monounsaturated fatty acid, C18:1 (oleic acid), was 46.1 in palm oil versus 18.7% in perilla oil. Polyunsaturated fatty acid C18:2 (linoleic acid) and C18:3 n-3 (ALA) were 8.6% and 0.3% in palm oil versus 13.1% and 62.3% in perilla oil, respectively. Then, the acid and PVs were compared with perilla oil with and without 0.5% tomato and paprika extract that was treated by SC-CO2. The perilla oil alone showed dramatic increases in acid and PVs during heat treatment, but these increases were inhibited by adding 0.5% tomato and paprika extract. This result indicates that the addition of tomato and paprika extract has an antioxidant effect and prevents lipid oxidation (Supplementary Fig. S1; Supplementary Data are available online at
Fatty Acid Compositions of Palm Oil and Perilla Oil Used in This Study (% Weight)
Effect of perilla oil on food intake and body weight gain in HFD-fed mice
Male C57BL/6 mice were fed with HFD and treated with palm oil (200 or 1000 mg/kg/day) or perilla oil (200 or 1000 mg/kg/day) for 90 days. The amount of 1000 mg/kg/day is equivalent to 5 g/day consumption in 60 kg human. Food intake and body weight gain were not different between palm oil-treated and perilla oil-treated mice (Fig. 1A, B). The HFD-fed mice that were not treated with oil exhibited more body weight gain than palm oil- or perilla oil-treated mice, possibly due to the increased food intake.
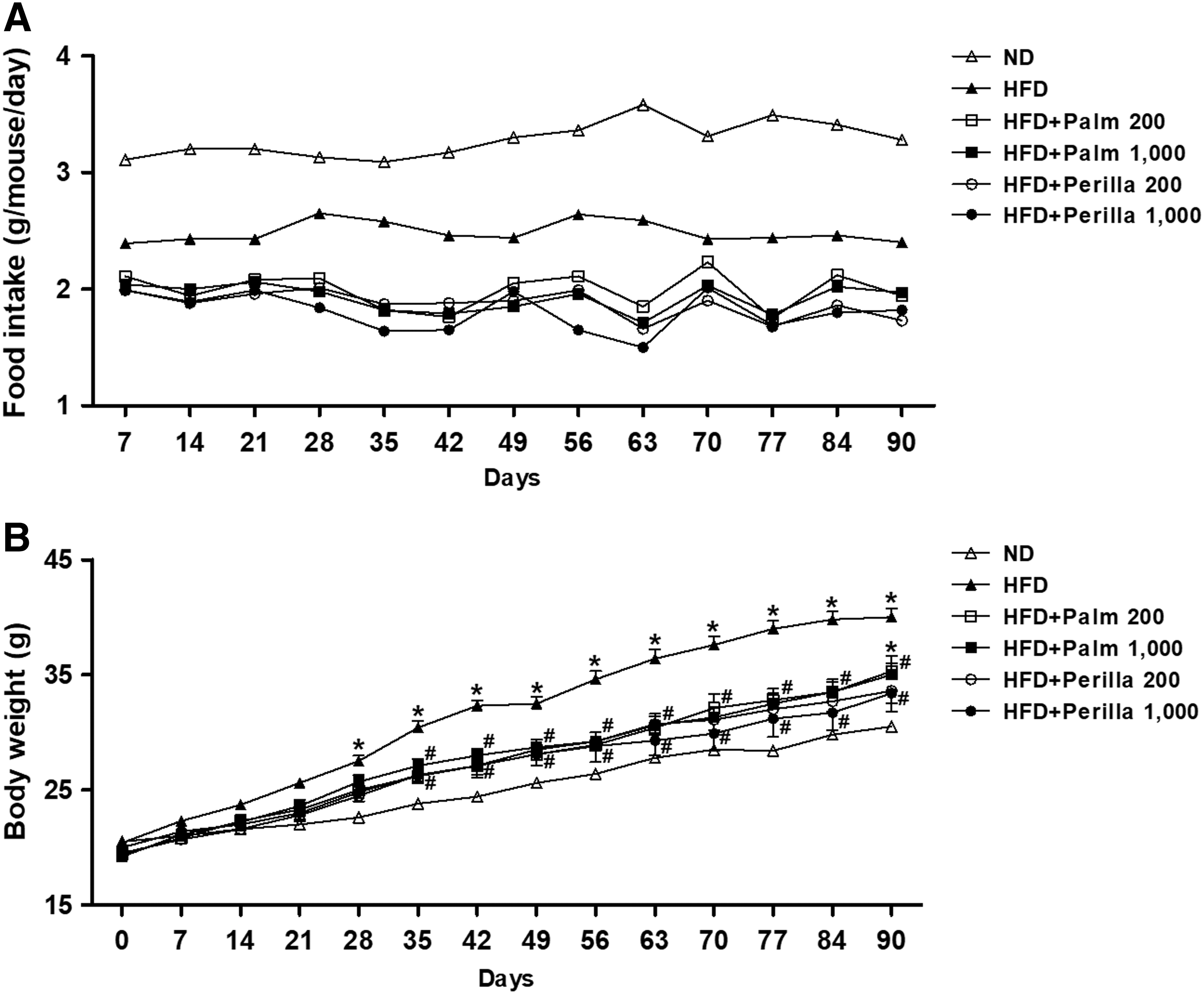
Effect of perilla oil on food intake
Perilla oil reduces serum cholesterol and arterial lipid accumulation in HFD-fed mice
Since perilla oil-treated mice showed a similar increase in body weight gain compared with palm oil-treated mice, we analyzed serum cholesterol levels at 30, 60, and 90 days of HFD feeding. Interestingly, perilla oil-treated mice exhibited a significant decrease in serum cholesterol levels compared with untreated or palm oil-treated mice (Fig. 2A), suggesting that perilla oil supplements may have a beneficial effect on lowering cholesterol levels in obese patients. Then, we examined the effect of perilla oil on thoracic aorta lipid accumulation by Oil Red O staining (Fig. 2B). The perilla oil (200 or 1000 mg/kg/day)-treated mice showed a significant reduction in arterial lipid accumulation compared with palm oil (200 or 1000 mg/kg/day)-treated mice. The results suggest that perilla oil attenuates HFD-induced arterial lipid plaque accumulation and may reduce a risk of atherosclerosis in obese patients with cardiovascular diseases.

Perilla oil reduces hepatic lipid accumulation in HFD-fed mice
Obesity increases the risk of NAFLD because excessive accumulation of triglycerides is a hallmark of diseases. We examined the effect of perilla oil on hepatic lipid accumulation and triglyceride levels in HFD-fed mice. Histological staining indicated that perilla oil treatment (1000 mg/kg/day) decreased HFD-induced hepatic lipid vacuole accumulation compared with palm oil treatment (1000 mg/kg/day), although a dose of 200 mg/kg/day of perilla oil did not show a significant reduction (Fig. 3A). Hepatic triglyceride levels were also reduced significantly in perilla oil-treated mice compared with palm oil-treated mice (Fig. 3B). However, there was no change in liver weight or the ratio of liver to body weight between perilla oil- and palm oil-treated mice (Supplementary Fig. S2).
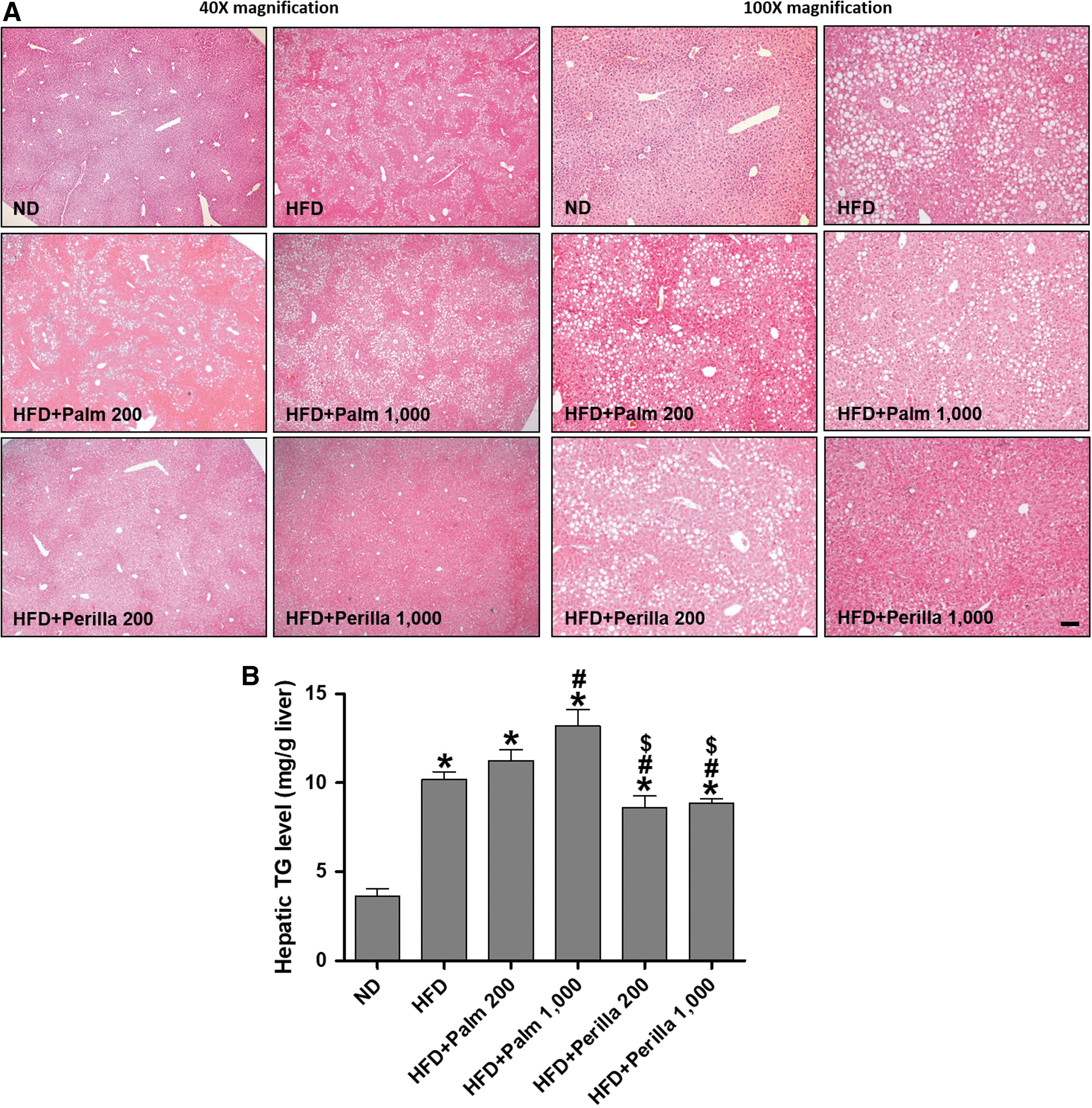
The effect of perilla oil on hepatic lipid accumulation in HFD-fed mice.
Perilla oil reverses HFD-induced alteration in lipid metabolic protein expression
The balance of lipid synthesis, oxidation, and transport has often been altered in metabolic disorders; thus, we examined whether perilla oil could affect the changes of proteins regulating lipogenesis and lipolysis in the liver from the HFD-fed mice (Fig. 4). AMPK activation inhibits cholesterol and fatty acid synthesis, and particularly increases fatty acid oxidation through ACC, a major substrate of AMPK. Palm oil treatment increased the p-AMPK in HFD-fed mice, and perilla oil further increased the level. Interestingly, perilla oil, not palm oil increased the p-ACC in HFD-fed mice. This result indicates that perilla oil has an effect on increasing fatty acid oxidation on HFD-induced metabolic stress. Then, we analyzed the expression of fatty acid transporter CD36 and ATGL in regulation of lipolysis. Palm oil treatment increased CD36 levels, but not ATGL, whereas perilla oil treatment increased both CD36 and ATGL levels. The lipogenic proteins of SCD-1 and FAS were also analyzed because their expression is highly correlated with hepatic steatosis. Palm oil treatment increased both SCD-1 and FAS protein levels, which were significantly reduced by perilla oil in HFD-fed mice. These results suggest that perilla oil improves the balance of lipolysis and lipogenesis, without changing HFD-induced body weight gain.
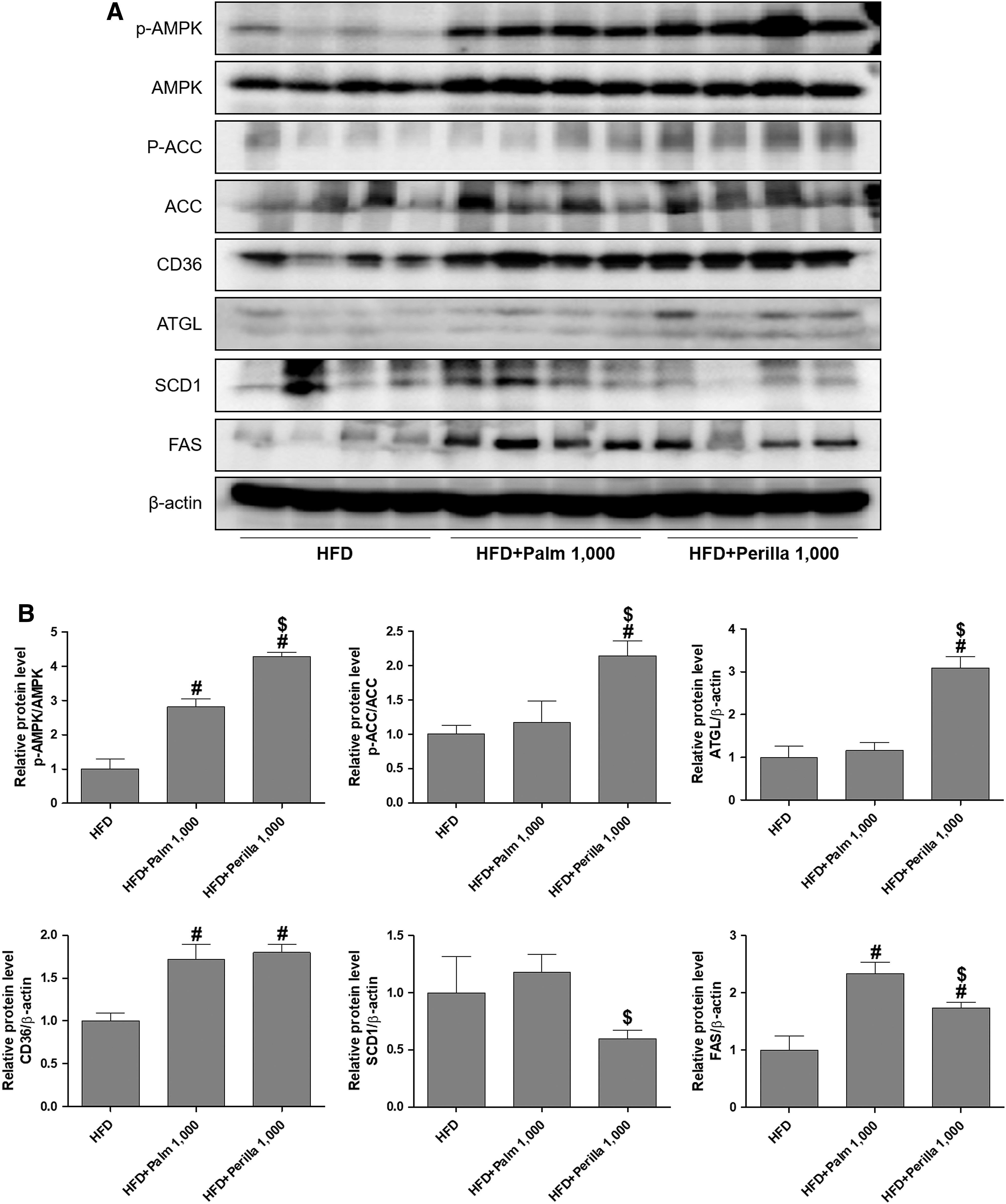
The effect of perilla oil on the hepatic expression of proteins regulating lipid metabolism in HFD-fed mice. Western blots
Discussion
Perilla oil is susceptible to oxidation, and oxidized linoleic acid, an abundant unsaturated fatty acid in perilla oil, is considered carcinogenic. 16 In this study, perilla oil was extracted under a cold-pressed method to prevent thermal degradation, and mixed with 0.5% tomato and paprika extract that was treated by SC-CO2. As shown in Supplementary Figure S1, the addition of 0.5% tomato and paprika extract markedly attenuated the increases in acid and PVs during heat treatment, suggesting the perilla oil prepared for our study has been protected from unwanted lipid oxidation. 17
The present study showed that perilla oil treatment reduced serum cholesterol and hepatic triglyceride levels compared with palm oil treatment in HFD-fed mice. However, there was no difference in the food intake and body weight gain between palm oil-treated and perilla oil-treated mice. Perilla oil treatment also attenuated lipid accumulation in the thoracic aorta and liver compared with palm oil treatment (1000 mg/kg/day). Therefore, we suggest that perilla oil may affect expression of lipid metabolic proteins, and showed that perilla oil increased p-AMPK, p-ACC, and lipolytic proteins, but decreased lipogenic proteins
Polyunsaturated fats are omega-3 and omega-6 fatty acids. Omega-3 is high in the oil from tuna, mackerel, flaxseed, and perilla seed, whereas omega-6 is high in the oil from corn, soybean, and sunflower seed. The balanced intake of omega-3 and -6 fatty acids was seriously disrupted after the increased consumption of soybean and corn that are genetically modified organism (GMO) crops, as well as the meat grown in GMO feeds in Western diets. The imbalance may be a reason for the increased incidence of pathogenesis of many diseases, including metabolic and cardiovascular disorders, inflammatory diseases, or cancer. 18
Perilla oil is particularly abundant in ALA, an omega-3 fatty acid, a precursor for EPA and DHA, which are important for proper body development and function. In addition, perilla oil is a rich source of phenolic compounds, flavonoids, and anthocyanins, which are known for their antioxidant and antimicrobial activities. 7 The previous studies have shown that omega-3 reduced hepatic and adipose tissue inflammation 8 and inhibited lipid mediators of inflammation (e.g., prostaglandins). 19 Omega-3 also improved cardiovascular function with an anticoagulant effect 9 and prevented neurodegeneration. 20 Perilla oil is a good alternative to fish oil as a source of omega-3 and its effect and underlying mechanisms remain under active investigation.
In this study, perilla oil-treated mice showed a significant increase in p-AMPK, an upstream kinase of ACC, and regulates cholesterol and fatty acid synthesis. Inactive p-ACC was increased only in perilla oil-treated mice, not in palm oil-treated mice; which indicates that perilla oil has directly activated β-oxidation of fatty acids in HFD-fed mice. CD36 and ATGL, the proteins for fatty acid uptake and hydrolysis were increased by perilla oil treatment, although the CD36 levels were also increased in palm oil-treated mice. CD36 facilitates fatty acid uptake and transmembrane transport, similar to the action of glucose transporter 4; fatty acids increased shortly the CD36 intracellular recycling and latter increased the CD36 gene transcription through peroxisome proliferator-activated receptors. 21 In this study, we demonstrated that long-term oil treatment may induce CD36 gene expression regardless of palm or perilla oil. Perilla oil treatment also reduced the expression of lipogenic proteins, SCD1 and FAS, in HFD-fed mice compared with palm oil. However, the reduced FAS levels were still higher than those of HFD-fed mice. We speculate that the long-term oil treatment in HFD-fed mice highly elevated lipogenic signaling, and that the lipid-lowering effect of perilla oil was not able to reduce to those of HFD-fed mice. We therefore suggest that perilla oil as a substituent for fat, which is recommended as 20–35% of total calorie intake per day. Perilla oil is also a better source of omega-3 than other oils. In the patients with dyslipidemia, a proper amount of perilla oil intake may improve the balance of lipid metabolism and be more beneficial than an absolute restriction of fat intake.
In conclusion, perilla oil reduced serum cholesterol and arterial and hepatic lipid accumulation by modulating lipogenesis and lipolysis in HFD-fed mice. Therefore, perilla oil is suggested as an omega-3 supplement that attenuates obesity-induced metabolic and cardiovascular disorders. Our mechanistic study and further understanding of perilla oil in the pathology of lipid metabolism is critical for developing its use as a therapeutic treatment of disease.
Footnotes
Acknowledgments
This study was supported by the Basic Science Research Program through the National Research Foundation (NRF) of Korea funded by the Ministry of Science, ICT, and Future Planning (NRF-2015R1A5A2008833).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.