
Abstract
We investigated the antitumor effect of Cordyceps militaris extract (CME) on A549 cisplatin-resistant (CR) lung cancer cells. The proliferation of A549/CR cells was suppressed by CME. Apoptosis of the cells was induced by CME. The cell cycle arrest was observed in the sub-G1 phase in the cells treated with CME. Proteomic profile analysis showed that H-Ras was downregulated in CME-treated cells and it was confirmed by western blot analysis. Collectively, these data demonstrated that CME is an alternative treatment for the anticancer effect.
Introduction
Lung cancer is one of the most deadly cancers in the world. Also in Korea, lung cancer has the highest mortality rate in both men and women. Nonsmall-cell lung cancer (NSCLC) accounts for 87% of all lung cancers. 1,2 Surgery is currently the most effective treatment for early NSCLC. 3 But most NSCLC patients are diagnosed after significant disease progression, with a 5-year survival rate of 17.4%, lower than many other area of cancers. 1,4
The chemotherapy based on cisplatin is usually the first-line chemotherapeutic administration in patients with advanced NSCLC. 5,6 Although, platinum-based chemotherapy can increase the life expectancy by 30–40%, the median overall survival is still not >10 months. 7 As a well-known DNA-damaging agent, cisplatin generally affects all rapidly dividing cells, and many normal cells throughout the body undergoing active growth and cell division are damaged. 8 The combination of radiation and chemotherapy promotes acute damage to normal cells. 9 Above all, cisplatin resistance has become a major clinical problem, because it causes the high tumor recurrence rate and subsequent treatment failure. Moreover, the cross-resistance with other platinum compounds severely elevates the difficulty of chemotherapy. 10 Therefore, it is imperative to find novel anticancer agents with fewer side effects and drug resistance.
Cordyceps militaris (CM), an edible Ascomycete, is an entomopathogenic fungus widely used in traditional medicine. Cordyceps genus has been used extensively as a tonic and health supplement for subhealth patients, especially seniors, in Korea and other Asian countries. In recent years, many pharmacological activities of this mushroom have been identified, including immunomodulatory, anti-inflammatory, antioxidant, antimicrobial, and antitumor effects. Well-known active ingredients within CM include cordycepin, cordycepic acid, sterols (ergosterol), nucleosides, and polysaccharides. Especially, cordycepin, a 3-deoxyadenosine, is the main active component of CM. 11 –13 Many studies have revealed that cordycepin has significant antitumor activities, such as inhibition of cell proliferation, migration, and induction of apoptosis. 14 –22 Also, cordycepin is one of the 18 new anticancer drugs currently being investigated and developed by the National Cancer Institute (NCI). 23 It is well established that CM or cordycepin had antitumor effects on various cancer cell models. In lung cancer cell models, it has been reported that antitumor activity in NSCLC cell line (NCI-H460), A549 lung cancer cell lines, human lung cancer cell line H1975. 16,17,24 But there was no study about antitumor effect of CM or cordycepin on cisplatin-resistant (CR) cancer cell models. Therefore, we showed that the potent anticancer effect of CM extract (CME) in CR cancer cell models.
Materials and Methods
Preparation of CME
CM (50 g) was provided from Yeonje Co., Ltd (Seoul, Korea). Fresh bodies or mycelia of CM were extracted with distilled water (D.W.) at 90–100°C for 3 h. The CME was filtered by using 200 meshes and evaporated under condition in vacuum. After drying at 60°C, 11.1 g of dried powder of CME was obtained.
Quantification and qualification of CME
Cordycepin and adenosine, the major compounds of CME, which structurally belong to nucleoside family, were obtained from Sigma-Aldrich Co. (St. Louis, MO, USA). high performance liquid chromatography (HPLC) analysis was performed using HP 1050 HPLC system (Agilent Technol., Santa Clara, CA, USA) with Symmetry Shield RP 18 column (Waters Co., Milford, MA, USA) at ambient temperature. The mobile phase consisted of water–methanol 85:15 (V/V) was run at a flow rate of 0.4 mL/min and the sample injection volume was 10 μL. Elution of adenosine and cordycepin was detected at 214 nm with the retention times of 15.1 and 18.8 min, respectively. Concentrations of adenosine and cordycepin in the sample were estimated using commercial adenosine and cordycepin as standard materials.
Ten milligrams of CME was dissolved in 50% methanol/water solution (5 mL). The extract was centrifuged and then the supernatant was collected for further analysis. The liquid chromatography (LC) separation was performed using a Acquity I-Class UPLC (Waters Corp., Baltimore, ML, USA) with an Acquity UPLC BEH C18 column (1.7 μm, 2.1 × 100 mm). The temperature of column was at 40°C. Mobile phase A and B were D.W and acetonitrile, respectively. The flow rate was 0.4 mL/min after the 2 μL injection. The MS detection was carried out on SYNAPT G2-Si system (Waters Corp.) with mean squared error of the data acquisition mode. The ionization used was electrospray ionization (ESI)-positive and negative. The source and the desolation temperature were 120 and 300°C, respectively. Leucine enkephaline was used for the lock mass compound. Positive and negative voltage ESI parameter were 3 and 2.5 kV, respectively. The collision energies were set as 6 eV (trap) for low energy and 20–45 eV ramp (trap) for high energy scan. The scan mass range was 100–1500 m/z. The liquid chromatography-mass spectrometry (LC-MS) data acquisition was controlled by MassLynx 4.1 (Waters Corp.). The acquisition data processing was performed by UNIFI1.8 with traditional medicine library.
Cell culture and cell viability assay
A549/CR human lung cancer cells were provided by a cell bank at ASAN Medical Center. The cells were cultured in RPMI1640 containing 10% fetal bovine serum (FBS) and 1 × antibiotics (Welgene, Dajeon, Korea). A549/CR cells were maintained routinely at 37°C in a humidified atmosphere of 5% CO2. A549/CR lung cancer cells (2 × 103 cells/well) were added to 96-well tissue culture plates. The cells were treated with CME or Afatinib as a negative control. After 72 h incubation, MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide] solution (5 mg/mL) was put into each well. After 2 h incubation at 37°C, the supernatants were removed, residual formazan crystals were solubilized in 100 μL of dimethyl sulfoxide (DMSO). Optical density was detected at 595 nm using an ELISA reader (Emax; Molecular Devices, USA).
Cell apoptosis assay
A549/CR cell apoptosis assay was determined using a Muse™ Annexin V and Dead Cell kit (EMD Millipore, Billerica, MA, USA) in accordance with the manufacturer's protocol. A549/CR cells were treated with CME or Afatinib for 24, 48, or 72 h, respectively. Total cells from each group were harvested and washed with phosphate-buffered saline (PBS). The cell pellets were suspended in RPMI1640 media containing 10% FBS mixed with the Muse Annexin V and Dead Cell reagent. After 20 min incubation at room temperature (RT) in the dark condition, optical density was detected using the Muse Cell Analyzer (Merck Millipore, Bedford, MA, USA).
Analysis of cell cycle
A549/CR Cell cycle was analyzed using a Muse Cell cycle kit (Merck Millipore). A549/CR cells were treated with CME or Afatinib for 24, 48, or 72 h, respectively. Total cells were harvested by centrifugation at 1000 rpm for 5 min. RPMI1640 media containing 10% FBS was used for suspension of the cell pellets, and then 70% ethanol cooled in ice was added to the cells. After 3 h incubation at −20°C, the cells were rinsed with PBS and treated with 200 μL of Muse Cell Cycle reagent (Millipore Corp., Bedford, MA, USA) according to the manufacturer's protocol. After 30 min incubation at RT in the dark condition, the cell suspension was analyzed using the Muse Cell Analyzer.
Caspase activity assay
A549/CR cells were harvested by using Typsin-EDTA after incubation with 6 μM of Afatinib and 1.5 mg/mL of CME for 72 h. The cell lysates were prepared and their protein concentrations were quantified. After 50 μL (100 μg) of protein extract was put in each well of 96-well plate, 1 M dithiothreitol (DTT) was added to reach the final concentration 0.1 M at each well. And then, 5 μL of caspase -3, -8, and -9 were added to each well and the plate was incubated at 37°C for 2 h. The optical density of each well was detected at 405 nm using ELISA plate reader.
Protein expression profiling using a protein chip-based antibody microarray
Each cell lysate (100 μg) was labeled with either Cy3 or Cy5 (GE Healthcare, UK) and free dyes were discarded by Spin column (S5059; Sigma, USA). To make an antibody microarray, glass slide coated with super epoxy group (SME2; Arrayit® Corporation, USA) was used. The antibody microarray using the glass chip was prepared by immobilizing 42 antibodies against cell cycle proteins. The cell lysates labeled with fluorescence dyes were applied on the antibody microarray. After 1 h incubated at 37°C in the dark condition, the antibody microarray chips were rinsed with phosphate buffered saline containing tween 20 (PBST) and dried using N2 gas. The chips were scanned using a GenePix 4100 A microarray scanner (Axon Instruments, Union City, CA, USA). The internally normalized ratio of all spots were calculated.
Western blot analysis
Protein extract (50 μg) was added into each well of 4–20% gradient gel (Mini-PROREAN® TGX Gels; Bio-Rad Pacific Ltd) and separated through gel electrophoresis. Gels were soaked in transfer buffer (30 mM glycine, 16 mM Tris-HCl, and 20% methanol) and proteins in the gels were then blotted to polyvinylidene difluoride membranes. Nonfat dry milk in PBST (137 mM NaCl, 27 mM KCl, 100 mM Na2HPO4, 20 mM KH2PO4 0.05% tween 20, pH 7.4) was used for blocking nonspecific binding sites. The polyvinylidene difluoride membranes were then incubated with anti-HRas mAb (1:200) or anti-β-actin mAb (1:10000). And then secondary antibodies (anti-mouse-1:10000, anti-rabbit–1:10000) were applied to the membranes. ECL western blotting detection kit was used for detection of protein bands in the membranes. The signals were then developed using X-ray films.
Statistical analysis
The experiment data are representative of three independent experiments with duplicate and expressed as the standard deviation. The values were compared with the control using analysis of variance followed by unpaired Student's t-test. P values of *P < .05, **P < .01, and ***P < .001 were regarded as statistically significant.
Results
Quantification and qualification of CME
HPLC-UV analysis was conducted to generate quantitative profiles of the compositions of CME. As shown in Figure 1, prominent peak of cordycepin was presented in the chromatograms of CME. Cordycepin was exclusively detected in the range of 18–20 min. The result demonstrated that CME contained ∼4.12 mg of cordycepin and 0.54 mg of adenosine per 1 g of CME (Fig. 1).

High-performance liquid ultraviolet chromatograms of CME. Cordycepin were detected ranging from 18 to 20 min. CME contained ∼4.12 mg/g of cordycepin and 0.54 mg/g of adenosine. CME, Cordyceps militaris extract.
In LC-MS spectrum, cordycepin was detected in the range of 136–137 sec. (Fig. 2). Qualitative components were presented in the CME as shown in Figure 3. Among them, cordycepin was detected in the range of 136–137 sec.
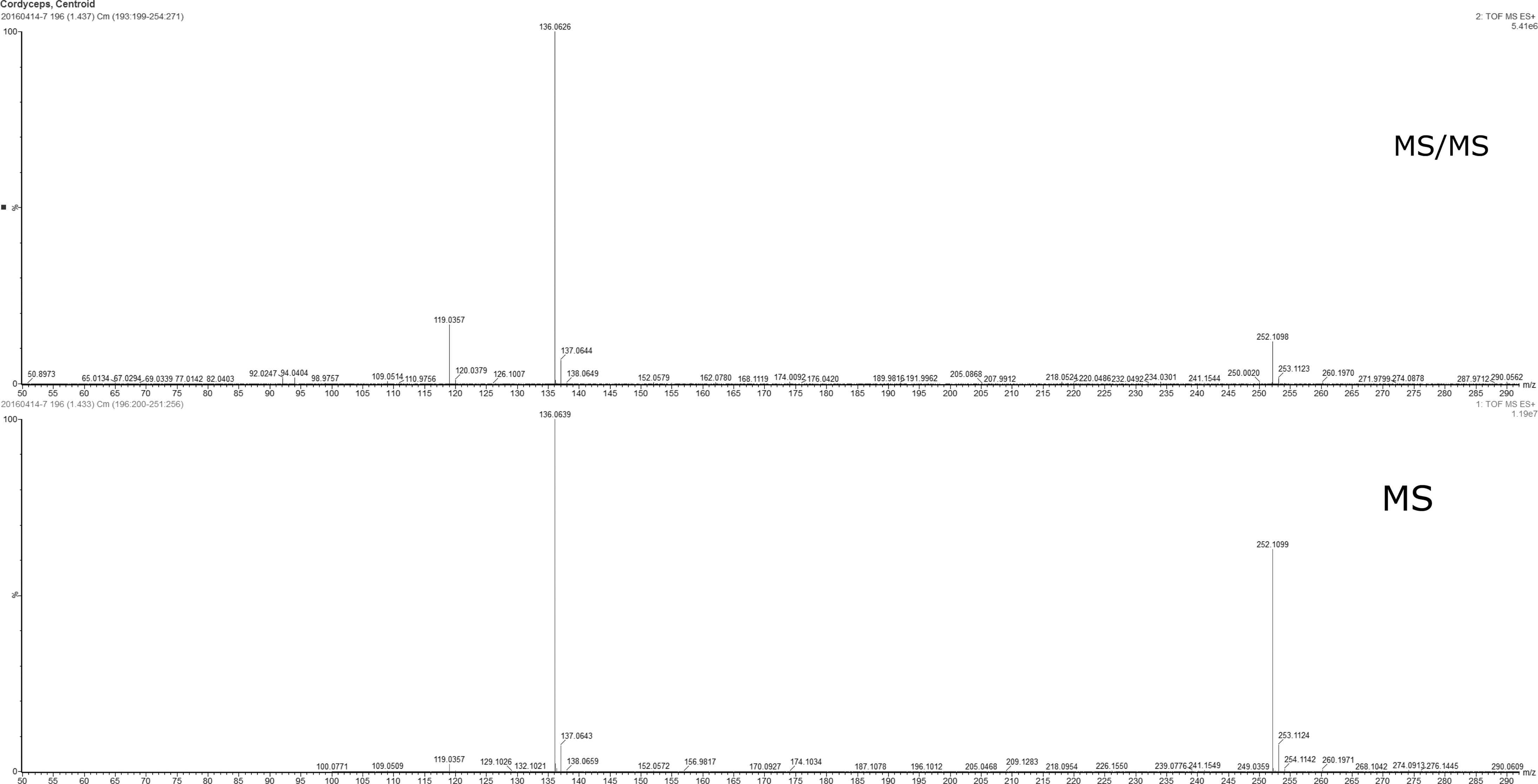
LC-MS spectrum (R.T 1.44, cordycepin). The high peak of cordycepin was present ranging from 136 to 137 sec. LC-MS, liquid chromatography-mass spectrometry. For ease of reading, the figure can be viewed and enlarged online.
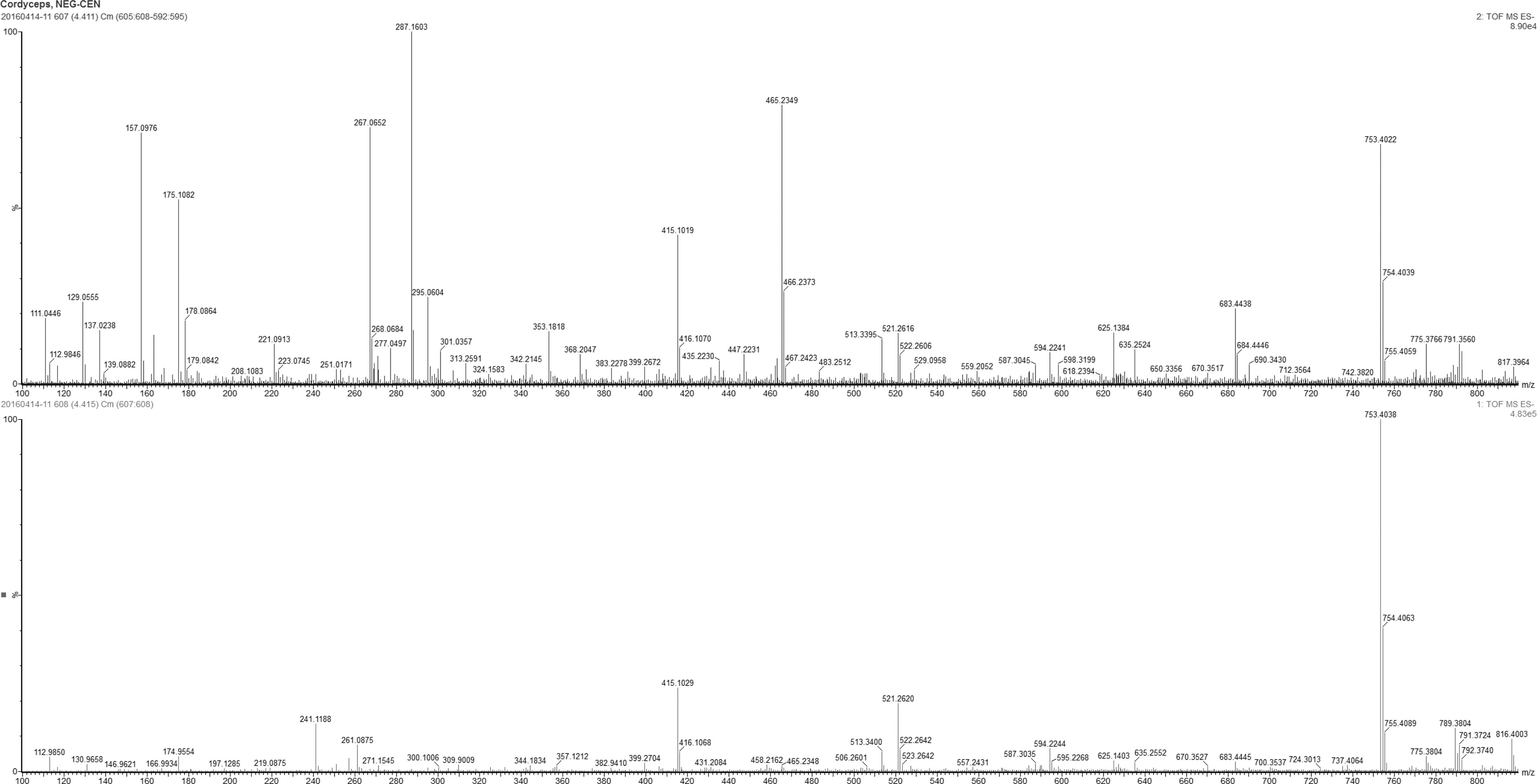
LC-MS spectrum (R.T 4.41, electrospray ionization-negative). Different qualitative components were present in the CME. Among them, cordycepin was detected ranging from 136 to 137 sec. For ease of reading, the figure can be viewed and enlarged online.
The inhibitory effect of CME on the viability of A549/CR cells
To examine the antitumor activity of CME, we carried out a cell-based viability assay using A549/CR cells. The result revealed that CME inhibited the cell viability of A549/CR cells in a dose-dependent manner. The half maximal inhibition of CME or Afatinib was observed at the concentration of 0.57 ± 0.12 mg/mL or 2.3 ± 0.23 μg/mL, respectively (Fig. 4A–C). These results suggested that the inhibition of A549/CR cell viability by CME may result in the CME-mediated cell apoptosis.
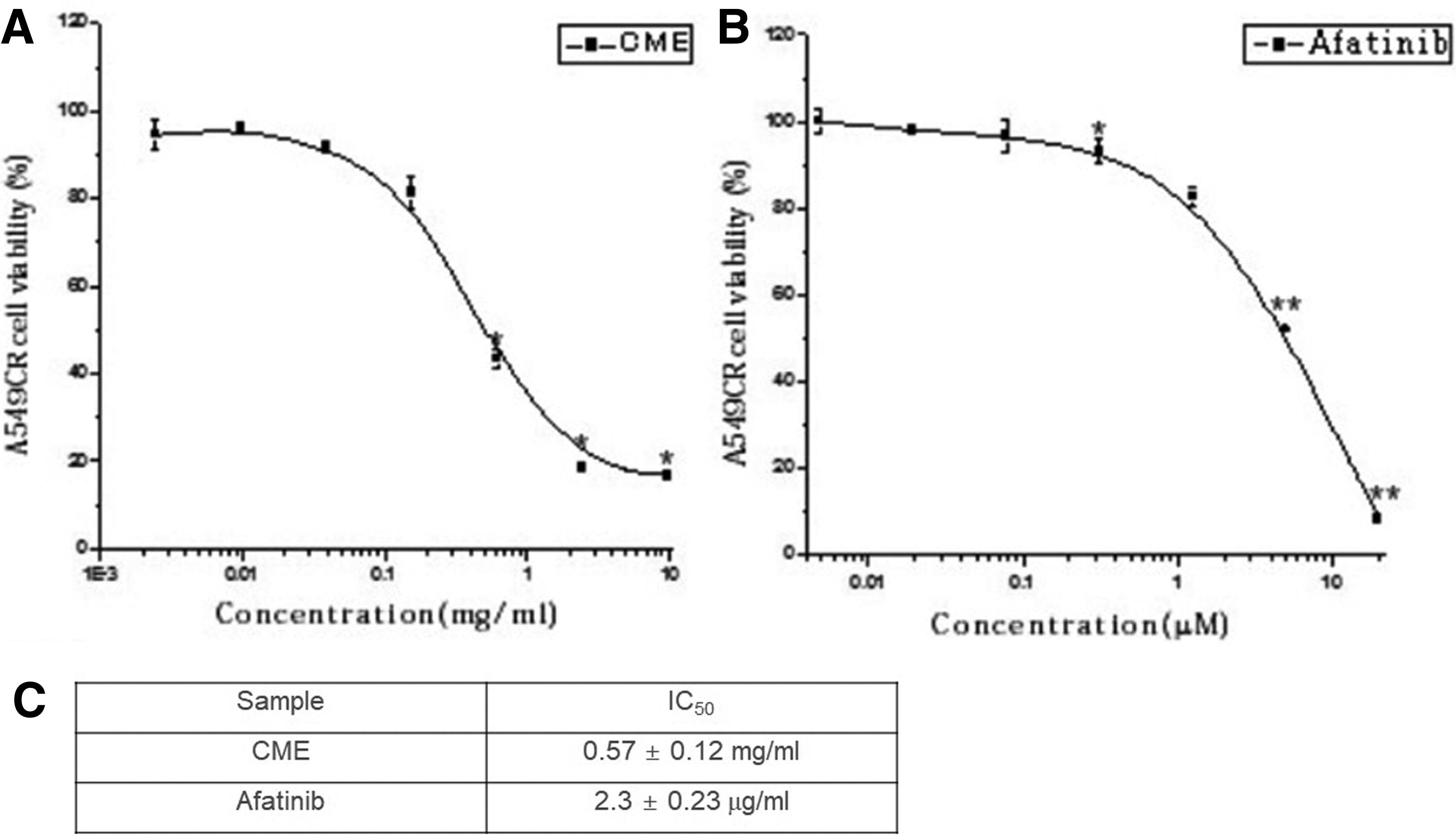
Inhibitory effect of CME on the viability of A549/CR lung cancer cells. A549/CR lung cancer cells were treated with different concentrations of CME and Afatinib. After incubation for 72 h MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide] assay was performed. IC50 value of A549/CR lung cancer cells by CME, and Afatinib observed at a 0.57 ± 0.12 mg/mL and 2.3 ± 0.23 μg/mL, respectively (*P < .05, **P < .01 versus control). CR, cisplatin-resistant.
The effect of CME on A549/CR cell apoptosis
To determine whether the antiproliferation function of CME was due to induction of apoptosis in the cells, we employed a cell apoptosis assay using Muse Annexin V and Dead Cell Kit. A549/CR cells were treated with CME or Afatinib at a concentration of 1.5 mg/mL or 2.9 μg/mL, respectively. The live cells and apoptotic cells were analyzed by Muse system when treated with CME or Afatinib. Time-dependent percentages of the live cells were decreased ranging from 78.32 to 17.63. The early apoptotic cells were significantly increased ranging from 19.18% to 42.80% in a time-dependent manner (Fig. 5). The total apoptosis were significantly higher in the both CME and Afatinib groups than the vehicle-treated control group.

Analysis of programmed cell death of A549/CR cells treated with CME.
Caspase activity of CME-treated A549/CR lung cancer cells
To confirm the induction of apoptosis in A549/CR cells treated with CME, in vitro caspase activity assay was carried out. Three types of caspases (caspase -3, -8, and -9) were activated in A549/CR cells treated with CME (1.5 mg/mL), whereas Afatinib (2.9 μg/mL)-treated group showed a minimal activation of A549/CR cells. These data suggested that CME enhanced caspase activation, resulting in stimulation of apoptosis in the cells (Fig. 6).
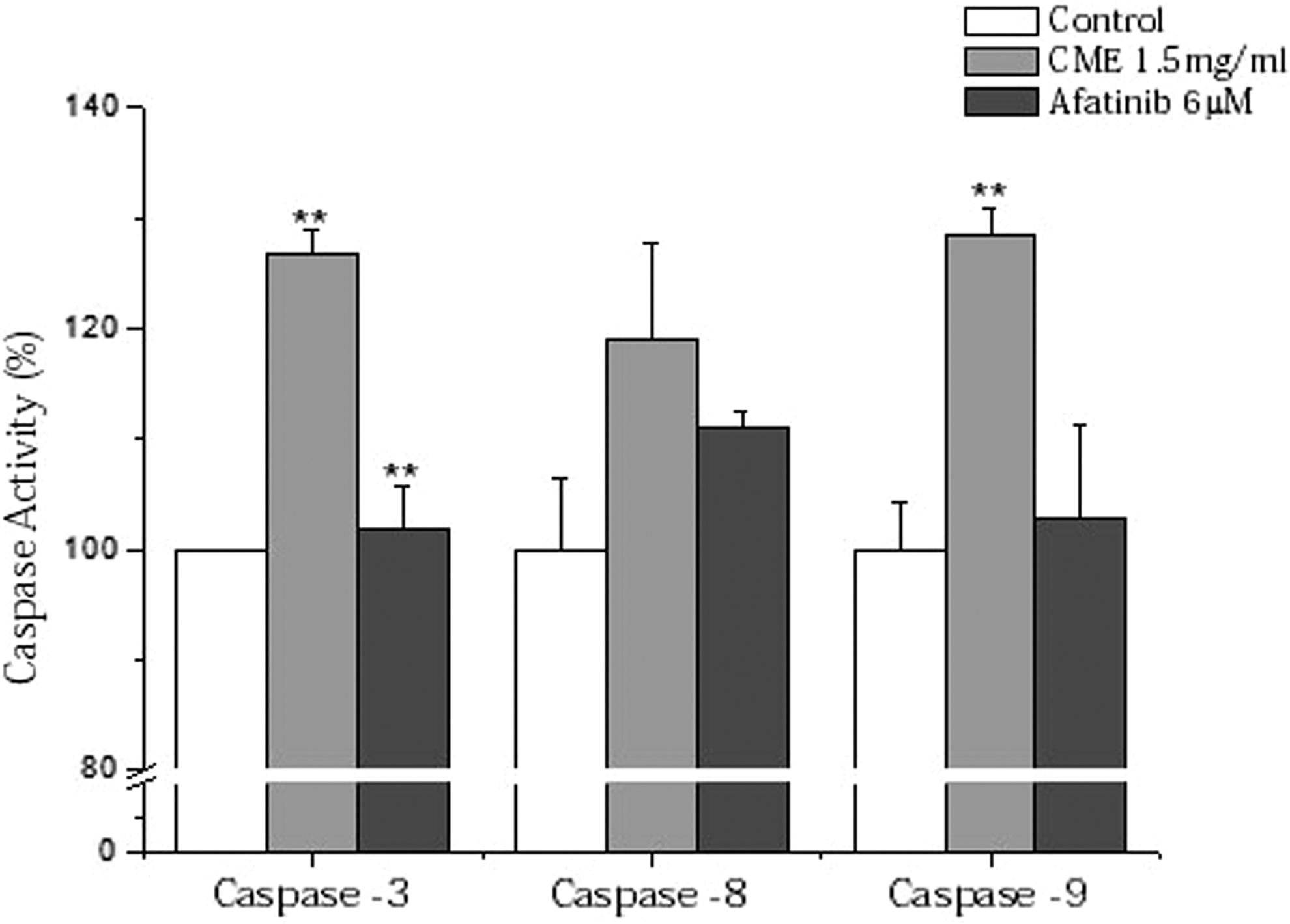
Analysis of caspases activities in A549/CR lung cancer cells. The bar graphs showed caspases activities of CME-treated A549/CR lung cancer cells compared with vehicle-treated control as percentages. Afatinib was used as a control (**P < .01 versus control).
Cell cycle analysis of CME-treated A549/CR cells
The cell cycle analysis was performed to determine which stage of cell cycle inCME-treated A549/CR cells was arrested (Fig. 7). When A549/CR lung cancer cells were treated with CME (1.5 mg/mL) or Afatinib (2.9 μg/mL), a significant time-dependent increase of S phase in both cells was observed. CME-treated A549/CR lung cancer cells showed 33.2 ± 1.2% of sub-G1 phase compared with of Afatinib-treated cells (17.6 ± 0.6%) and vehicle-treated control group (13.3 ± 1.6%) at the 48 h (Fig. 7). Consequently, these findings demonstrated that CME increased accumulation of S phase in A549/CR lung cancer cells followed by stimulation of the cell apoptosis.
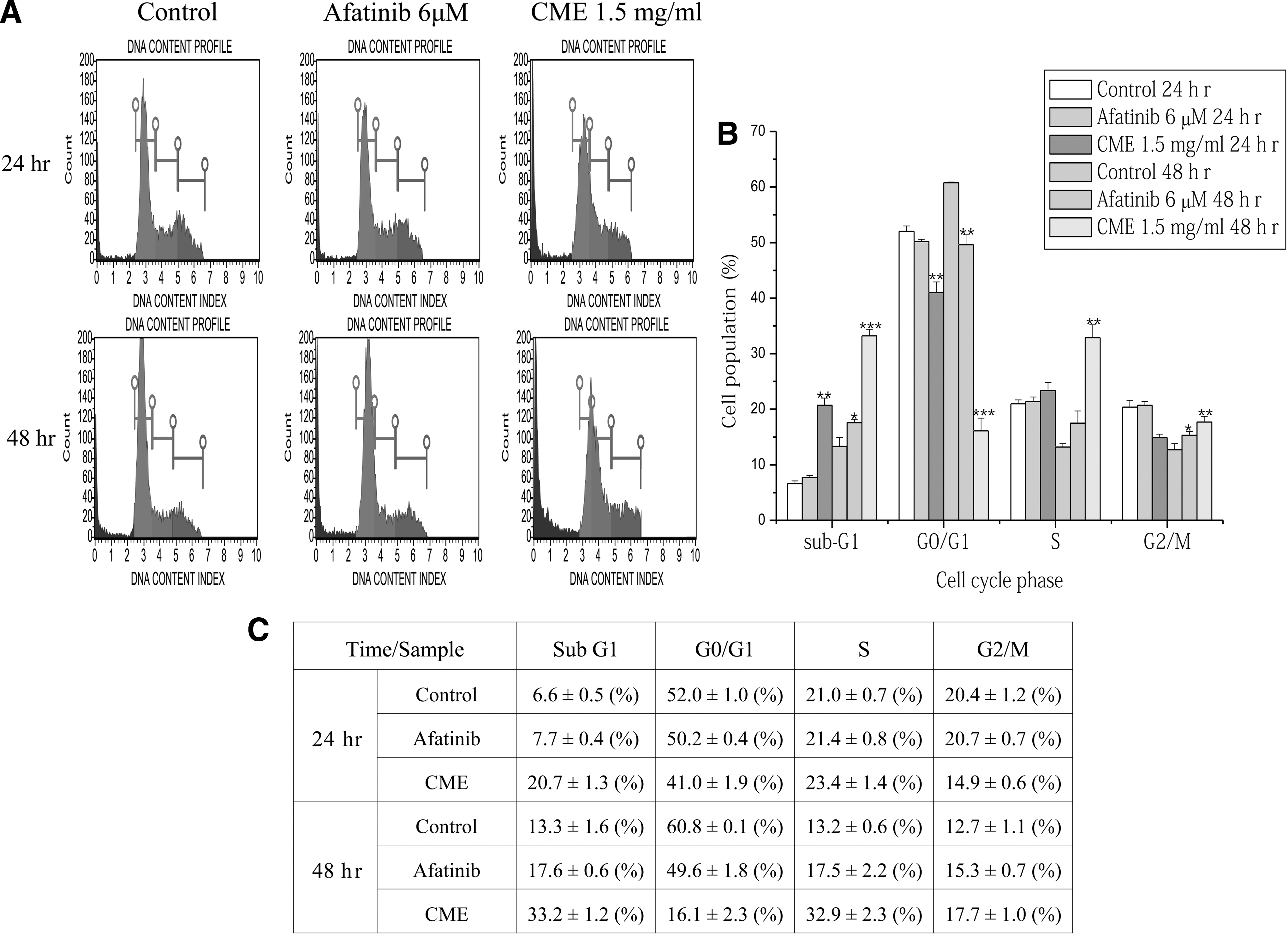
Cell cycle analysis of CME-treated A549/CR lung cancer cells. The cell cycle analysis of A549/CR lung cancer cells treated with CME were performed by using DNA content index parameter to determine G0/G1, S phase, and G2/M. The peaks of Green, blue, red, and olive colors appeared sub-G1, G0/G1, S and G2/M phases, respectively. The bar graphs showed the percentages of the cells in different cell cycle phase at different times (24 or 48 h). (*P < .05, **P < .01, ***P < .001 versus control).
Proteomic analysis of CME-treated A549/CR lung cancer cells
To investigate the molecular mechanism of A549/CR cells treated with 1.5 mg/mL of CME, proteomic profiling was carried out using a protein chip-based antibody array. Lysates from the CME-treated A549/CR lung cancer cells were labeled with a fluorescent dyes, Cy5 or Cy3 after which the cross-reactivity of both samples was investigated (Fig. 8A). Labeled protein extracts were applied on the antibody array to analyze the expression profile of cell cycle proteins in CME-treated cells. Fluorescence intensities of all spots were detected using a fluorescent scanner. Based on the data, H-Ras was significantly downregulated in CME-treated cells (Fig. 8B, C). There was no difference in expression levels of all other proteins. The expression data were confirmed by immunoblot analysis (Fig. 8E).
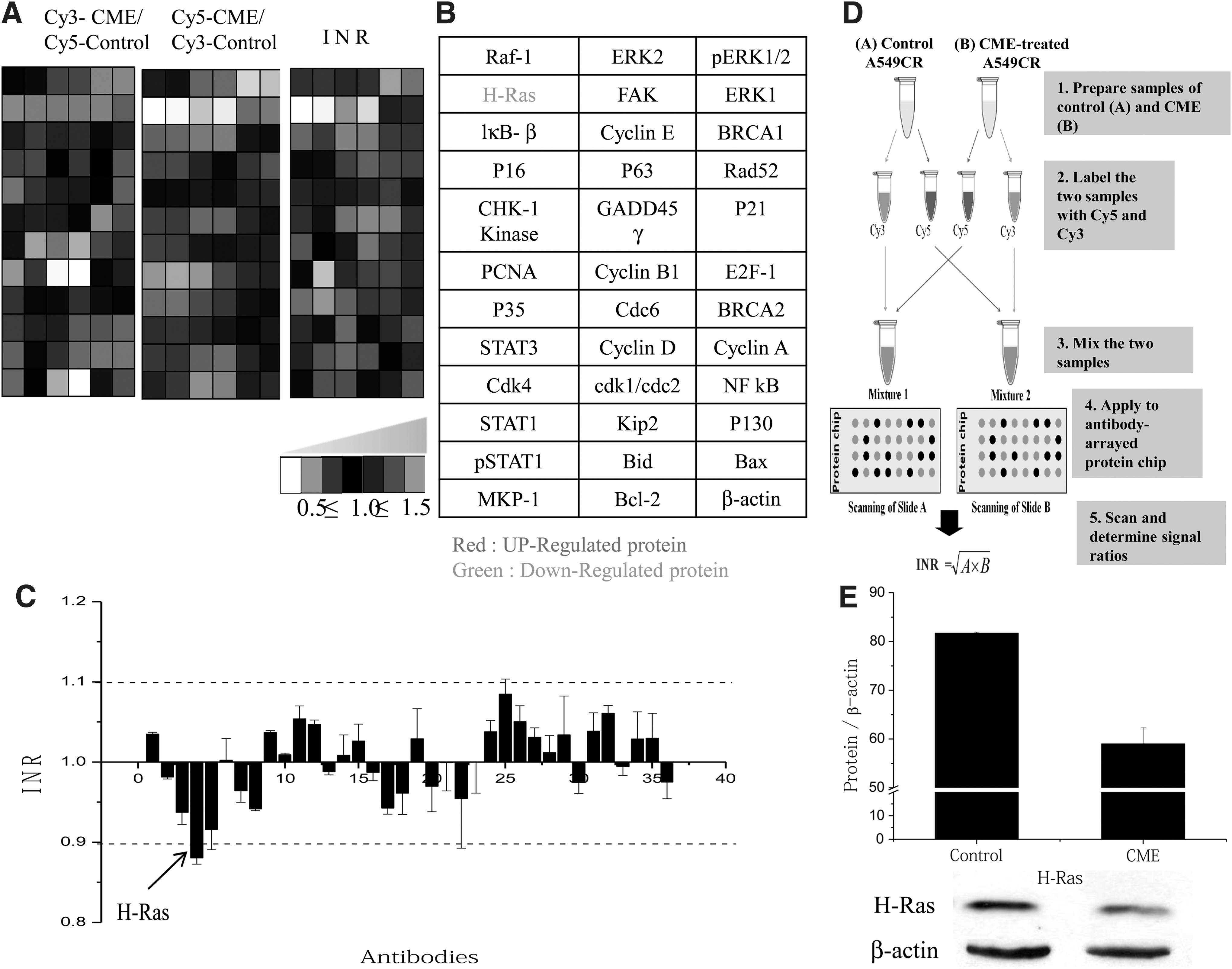
CME expression profiling using antibody microarray and western blot analysis. Graded virtual image of Cy5:Cy3 ratios from A549/CR cells treated with CME
Discussion
The human Ras protein family is composed of H-, N-, and K-Ras. And these mutations can transform normal cells into neoplasm. H-Ras is a small guanosine triphosphate-binding protein that is getting involved in transferring extracellular signals within cells. The Ras signaling pathway plays a critical role in regulation of cell growth, proliferation, survival, and apoptosis corresponding to various extracellular stimuli. Activation of H-Ras promotes cell proliferation by regulating cell cycle progression in most cancers. 25,26 Activated Ras protein stimulates downstream effecters such as the Raf/MEK/ERK (also known as the MAPK/ERK) and PI3K-Akt pathways. Ras-mediated stimulation of the MEK/MAP kinase and PI3K-Akt pathway upregulates cellular levels of cyclin D1, a critical player in cell cycle progression. Enhanced levels of cyclin D1 promote cell cycle progression through the G1 checkpoint and into S phase. 27 –29 In this study, CME induced sub-G1 arrest of A549/CR cells and regulated the expression of H-Ras, MEK-1, and MEKK-1. Also, CME significantly suppressed the cell viability of A549/CR cells. These results demonstrated that CME controlled cell cycle progression by inhibiting Ras downstream signaling, resulting in suppressing the proliferation of A549/CR cells. Recently, it was reported that Cordycepin, a major active compound of CME showed antiproliferative effect on A549/CR cells through downregulation of H-Ras. 30 These data strongly suggested that one of the major compound in CME inhibiting the proliferation of A549/CR cells is due to cordycepin.
The Ras/MAPK and PI3K/Akt are involved in activation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). NF-κB promotes the transmission of several genes and the inhibitors of apoptosis, which is involved in the perturbation of apoptosis induced by both intrinsic and extrinsic pathways through direct suppression of caspases (caspases-3, -6, -7, and -9). 31,32 Caspases act as important mediators of apoptosis and contribute to the overall apoptotic morphology by cleaving various cellular substrates. Chen et al., also reported that H-Ras is a crucial regulator of death receptor-mediated apoptosis pathways. 33 In this study, CME increased the caspases activities that lead to the apoptosis of A549/CR cells. Based on the aforementioned results, we presumed that CME promote caspase-dependent apoptosis by inactivating NF-κB, the key regulator of Ras-mediated apoptosis.
Because of the high percentage of human tumors harboring oncogenic Ras mutants, interrupting the Ras-signaling pathway has been a major focus of new drug-development efforts. The major approaches taken are as follows: (1) the inhibition of Ras protein expression; through ribozymes, antisense oligonucleotides, or RNAs; (2) the prevention of membrane localization of Ras; and (3) the inhibition of downstream effecters of Ras function. 28 To the best of our knowledge, this study is the first to report the antitumor effects of CME on A549/CR cells by downregulating of H-Ras-mediated cellular signaling pathway.
Cisplatin is the first-line regimen in advanced NSCLC. Since patients with NSCLC usually required for long-term treatment, drug resistance is a well-known limitation of cisplatin-based chemotherapy. As a common strategy to overcome drug resistance, high dose of chemotherapeutic agents are administered. However, adverse effects are usually increased as a consequence. 34 To reduce the adverse effects, in recent years, targeted therapies were developed. EGFR TKIs is one of the molecule targeting agents, which is used to the patients with EGFR mutations. 34 Wang et al., reported that targeted inhibition of Apurinic/apyrimidinic endonuclease (APE1) enhanced the activity of cisplatin in A549 cells. 35 Recent studies suggested that the microRNA regulated cisplatin resistance in lung cancer cells. Also, siRNA has been investigated to overcome cisplatin resistance. 36 –38 Recently, Wang et al., identified that Matrine (an alkaloid from Sophora flavescens) induced apoptosis in CR NSCLC cells through inhibition of β-catenin/survivin signaling. 39 In our study, CME inhibited the proliferation of A549/CR cells by downregulating H-Ras and inducing sub-G1 arrest, and caspase-dependent apoptosis. Taken together, our data suggested that CME showed the possibility of overcoming the cisplatin resistance in NSCLC.
Footnotes
Acknowledgments
This research was supported in part by grants of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare (grant no. HI14C0849).
Author Disclosure Statement
No competing financial interests exist.