
Abstract
Inflammatory bowel disease (IBD), including ulcerative colitis (UC) and Crohn's disease, is a group of chronic and relapsing inflammatory conditions within the gastrointestinal tract. An increase in intestinal epithelial cell (IEC) apoptosis is a major characteristic of UC. Tumor necrosis factor-α (TNF-α) plays an essential role in the regulation of apoptosis. Aberrant activation of the immune response to resident microflora contributes to overproduction of TNF-α in the mucosal tissue of the gastrointestinal tract; a hallmark of UC. There are no curative medications for IBD. Thus, establishment of novel strategies for the treatment of this disease is imperative. Lactic acid bacteria (LAB) have been characterized as probiotics that can alleviate imbalances in indigenous microflora in UC, exhibiting beneficial effects for the treatment and prevention of IBD. In this study, we elucidate the potential of LB-9, a novel probiotic LAB, to protect against colitis development using a dextran sodium sulfate (DSS)-induced mouse model of UC. Treatment using LB-9 reduced clinical symptoms of colitis. In addition, both colitis-induced and NF-κB-mediated IEC apoptosis was markedly reduced in mice treated with LB-9. Moreover, these results were closely associated with reduced TNF-α levels. Our study demonstrates that the LB-9 probiotic exhibits therapeutic potential for UC through suppression of TNF-α-mediated IEC apoptosis in a murine DSS-induced colitis model, with important biological implications for treatment of IBD in humans.
Introduction
Ulcerative colitis (UC) is a major form of inflammatory bowel disease (IBD) and is characterized by relapsing and chronic inflammation in the gastrointestinal tract. 1 –3 A feature commonly observed in UC pathogenesis is functional disruption of the intestinal epithelial barrier, which is linked to aberrant apoptotic death in intestinal epithelial cells (IECs) due to exacerbation of immune responses against resident microbial flora. 4,5 Therefore, persistent control of IEC apoptosis during UC is critical to maintaining homeostasis and functionality in IEC. 6
Excessive production of tumor necrosis factor-α (TNF-α), a characteristic of UC, contributes to the dysregulation of mucosal immunity to commensal microflora. 7,8 In particular, IEC apoptosis is considerably increased in UC, and TNF-α plays a central role in determining the apoptotic pathway involved. 9,10 Accordingly, inhibition of TNF-α using antibodies against the cytokine was reported to result in ulcer healing associated with mucosal regeneration. 7
Prevention and reduction of symptoms are among the most extensively evaluated factors among UC therapies. 11 Current medical treatments for UC involve pharmacological interventions, such as the administration of anti-inflammatory drugs and/or antibiotics. However, these therapies are often associated with significant adverse side effects, and thus there is a need to develop novel therapeutics with minimized undesirable effects. 12 Due to the effects of IBD on the colon and intestinal microbiota, nutritional therapy incorporating probiotics is increasingly recognized as a reliable treatment. 13,14 Therefore, study of probiotic preparations that modulate the intestinal microflora are an appropriate line of investigation.
Probiotics, live microorganisms, confer health benefits to the host when administered in adequate amounts, 12 and administration of probiotics has been shown to induce remission and alleviation of symptoms of UC in both animal and human studies. 15 –17 Recently, it was reported that lactic acid bacteria (LAB), the most common commercial probiotic bacteria, improves the functions of indigenous microflora and exerts positive effects in UC. 18 –20 However, to date, sufficient research data are not available to establish appropriate applications of LAB for ameliorating UC; consequently, the therapeutic potential of probiotics in UC remains unclear.
Therefore, to determine the therapeutic value of a putative LAB species for UC, we isolated the novel probiotic, Lactobacillus plantarum LB-9 from Kimchi and assessed its therapeutic potential in a dextran sodium sulfate (DSS)-induced colitis mouse model. We found that LB-9 ameliorates colitis by suppressing the apoptosis of IECs through downregulation of NF-κB activity. Moreover, TNF-α expression was significantly reduced in IECs of mice with DSS-induced colitis treated with LB-9. Thus, these findings suggest a mechanism of protection against UC in which LB-9 downregulates TNF-α production, followed by the downregulation of NF-κB-mediated apoptosis of crypt IECs in UC. Our study sheds light on the underlying mode of action of the probiotic LB-9 in UC, thereby, representing a novel protective approach for the treatment of IBD, particularly colitis.
Materials and Methods
Reagents
DSS was provided by MP Biomedicals (Santa Ana, USA; molecular weight 36,000–50,000, colitis grade). The 2,2,2-tribromoethanol, citric acid, and Tween 20 were purchased from Sigma-Aldrich (St. Louis, USA). Four percent paraformaldehyde was prepared with paraformaldehyde (Samchun Chemicals, Pyeongtaek, Korea). Histosec® paraffin and protease inhibitor cocktail were provided by Merck (Merck Millipore, Darmstadt, Germany). The antibodies used were anti-β-actin (Novus Biologicals, Littleton, CO, USA), anti-cleaved-caspase 8, anti-phospho-p65, anti-cleaved-PARP, anti-cleaved-caspase 3, and anti-phospho-IκB (Cell Signaling Technology, USA), anti-TNF-α, and anti-TNF receptor (Abcam, Cambridge, UK). The In Situ Cell Death Detection Kit was obtained from by Roche (Basel, Switzerland).
Animal care and experimental protocols
All animal experiments were approved by the Korea University Institutional Animal Care and Use Committee (Approval No. KUIACUC-2018-52). Experiments were performed using 8-week-old male C57BL/6J mice. The mice were kept in microisolator rodent cages at 22°C on a 12-h light–12-h dark cycle. Food and drinking water were provided ad libitum throughout the experimental period. Experimental colitis was induced by adding 3% DSS to the drinking water for 6 days. A concentration of 1.2 × 10 10 CFU of LB-9 probiotic was administered by oral gavage twice a day, starting 7 days before the DSS treatment. Colitis development was determined by change in body weight and fecal condition. The feces were examined on days 0, 3, and 6. At day 7 of the colitis induction period, the mice were sacrificed. The mice were anesthetized with 250 mg/kg body weight of 2,2,2-tribromoethanol. Colon tissues from cecum to rectum were harvested, and the colon lengths were measured. The isolated colons were immediately frozen and stored at −80°C.
Evaluation of colitis
The disease activity index (DAI) of colitis was scored by the loss of body weight, stool consistency, and presence of blood in the feces, as described previously. 21 DAI scoring was conducted by a blinded observer.
Histopathological analysis
Colon tissues were fixed in 4% paraformaldehyde and embedded in paraffin. The 3 μm sections were cut from the paraffin blocks and stained with Hematoxylin and Eosin. Scoring of histological damage was measured by two categories: inflammatory cell infiltration and tissue damage by ulceration. The infiltration of inflammatory cells was graded on a scale from 0 to 3 (0, none; 1, mild; 2, moderate; and 3, severe) in each layer of the colon, including epithelium, crypts, stroma, submucosa, and transmural layer. The severity of ulceration was graded histologically on a scale from 0 to 4 (0, none; 1, mild and focal surface; 2, mucosal layer; 3, into colon wall). 22 We modified and established a 0 to 7 scoring system by summarizing all the scores.
Immunofluorescence
The 3 μm sections were deparaffinized and rehydrated. The tissues were permeabilized with citrate buffer (10 mM citric acid; 0.05% Tween 20, pH 6) by irradiation using microwave, twice, for 2 min. The tissues were blocked with 3 g/100 mL bovine serum albumin (BSA) in phosphate-buffered saline (PBS) for 1 h at 25°C. After blocking, tissues were treated with the appropriate primary antibody overnight at 4°C. The tissue sections were rinsed with PBS and incubated with the appropriate fluorophore-conjugated secondary antibody. Fluorescent antibody-stained slides were then treated with an anti-photobleaching reagent and sealed with a cover glass; 3,3′-diaminobenzidine-stained slides were mounted on glass slides. All slides were analyzed by microscopy (Axiovert 40 CFL; Carl Zeiss AG, Germany).
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling staining
The evaluation of apoptosis was performed with colon tissue sections and terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining using the In Situ Cell Death Detection Kit, following the manufacturer's protocol. The final mounting was done after counterstain by 4′,6-diamidino-2-phenylindole (DAPI). The TUNEL-DAPI-stained slides were analyzed with Axiovert 40 CFL microscope (Carl Zeiss AG, Germany).
Immunoblot analysis
The frozen colon tissues were homogenized on ice in radioimmunoprecipitation assay buffer with a protease inhibitor cocktail and PMSF. Samples were then sonicated. Sonicated tissues were centrifuged at 15,000 g for 45 min at 4°C and then stored at −20°C until use. Bradford reagent (Bio-Rad, München, Germany) was used for measuring protein concentration against a BSA standard. Proteins were denatured with sample buffer for 5 min at 100°C, separated on a 12% polyacrylamide gel, and blotted onto 0.2-mm polyvinylidene difluoride membranes. The membranes were blocked with 3 g/100 mL BSA in 0.05% TBS-T (50 mM Tris, 150 mM NaCl, and 0.05% Tween-20) for 1 h at 25°C and then incubated overnight at 4°C with primary antibodies. After incubation with the respective primary antibodies, the membranes were washed three times for 5 min in 0.05% TBS-T and then incubated with species-specific horseradish peroxidase-labeled secondary antibody for 1 h at 25°C. After washing the membranes three times for 10 min in TBS-T, the reaction was allowed to develop using the Amersham ECL Prime Western Blotting Detection Reagent (GE Health Care Life Sciences). The signal intensities were measured on an ImageQuant LAS 4000 (GE Health Care Life Sciences).
In vivo bioluminescence imaging
The probiotic LB-9 labeled with 500 μM Rhodamine 123 in DMSO was orally administered to C57BL/6J mice. Bioluminescence imaging was performed by placing the mice on the Xenogen Lumina II system 1 h after probiotic administration. The signal was analyzed using the Living Image 4.2 software.
Statistical analyses
Statistical analyses were presented as mean ± standard deviation (SD). Analyses were performed using a two-tailed t-test. P-value ≤.05 was considered statistically significant.
Results and Discussion
LB-9 has a protective effect against DSS-induced colitis
To determine whether LB-9 administration prevented DSS-induced acute colitis, mice were subjected to oral administration of LB-9 followed by treatment with DSS. Drinking water containing 3% DSS was provided to the mice and the disease phenotype of colitis was then assessed by DAI during the last 6 days of experimentation. UC is characterized by a loss of body weight, abdominal discomfort, and bloody diarrhea. 3,12 As expected, the DAI value was remarkably increased in mice treated with DSS compared with that in untreated control mice (Fig. 1A). However, pretreatment with LB-9 resulted in significantly lower DAI value in mice subjected to DSS treatment (Fig. 1A), suggesting that LB-9 has the capability to reduce the susceptibility of the mice to DSS-induced colitis.

Protective effect of LB-9 against DSS-induced colitis.
The development of DSS-induced colitis is closely associated with pathological damage in the colon. Hence, colon length was measured in the mice after DSS administration. Following DSS exposure, a marked reduction in the colon length of DSS-treated mice was observed compared with control mice. However, LB-9 treatment significantly attenuated the decrease in colon length in the DSS-treated mice (Fig. 1B, C). In addition, DSS treatment resulted in colitis-associated histological alterations, such as structural disruption of the surface epithelium, infiltration of inflammatory cells, and disruption of cryptal glands (Fig. 1D). The pathological characteristics of colon tissues in DSS-treated mice were lessened in mice treated with LB-9 (Fig. 1D), indicating a protective effect against DSS-induced colitis.
LB-9 treatment protects against DSS-induced IEC apoptosis
Next, we aimed to determine the point at which LB-9 began to alleviate DSS-induced colitis. It is well established that the barrier function of IECs against luminal microbes is central to IBD pathogenesis. Hence, increased cell death of IECs unequivocally results in the disruption of the normal intestinal epithelium, which is frequently observed in the inflammatory sites of IBD, including UC. 4,5,23
To examine the protective effect of LB-9 on IEC apoptosis following DSS treatment, we performed TUNEL staining on colon tissue harvested from mice with DSS-induced colitis. Figure 2A shows that after DSS treatment, the apoptotic index of IECs was remarkably enhanced compared with that seen in control mice, and importantly, this increased sensitivity to apoptosis was markedly reduced in mice treated with LB-9 before DSS exposure.

Effect of LB-9 on DSS-induced IEC apoptosis.
To further evaluate the protective effect of LB-9 against apoptosis, the activity of cleaved PARP and cleaved caspase-3 was assessed in colon tissues by immunoblot analysis. Consistent with TUNEL results, apoptosis in IEC was increased following DSS treatment, as indicated by increased levels of both PARP and caspase-3. Furthermore, the increased apoptotic activities observed in IECs from the DSS-treated mice were remarkably reduced in mice that had received LB-9 treatment (Fig. 2B). These results indicate that LB-9 protects against apoptotic death of IECs, which is reflected in the reduced susceptibility of the mice to colitis.
LB-9 treatment reduces TNF-α-induced cell death in IECs
It is reported that DSS-induced colitis is closely related to activation of proinflammatory cytokines, such as TNF-α, IL-6, and IL-10. 24 In particular, TNF-α plays a central role in the elevation of intestinal mucosal inflammation in IBD. Moreover, expression of TNF-α is increased in IBD patients, which contributes to the increased apoptotic death in the IECs of colon tissues. 25,26
Thus, we investigated whether LB-9 treatment intervened with the apoptotic cell death induced by TNF-α in IECs of mice with DSS-induced colitis. To analyze the influence of LB-9 on TNF-α expression during colitis, the presence of TNF-α in colon tissue was evaluated by immunohistochemical analysis. The level of TNF-α was significantly elevated in the colon tissues of mice exposed to DSS (Fig. 3A). However, the level of TNF-α was significantly reduced in the intestines colonized with LB-9. These results suggest that the decreased levels of TNF-α following LB-9 treatment led to the inhibition of DSS-induced apoptosis of IECs.
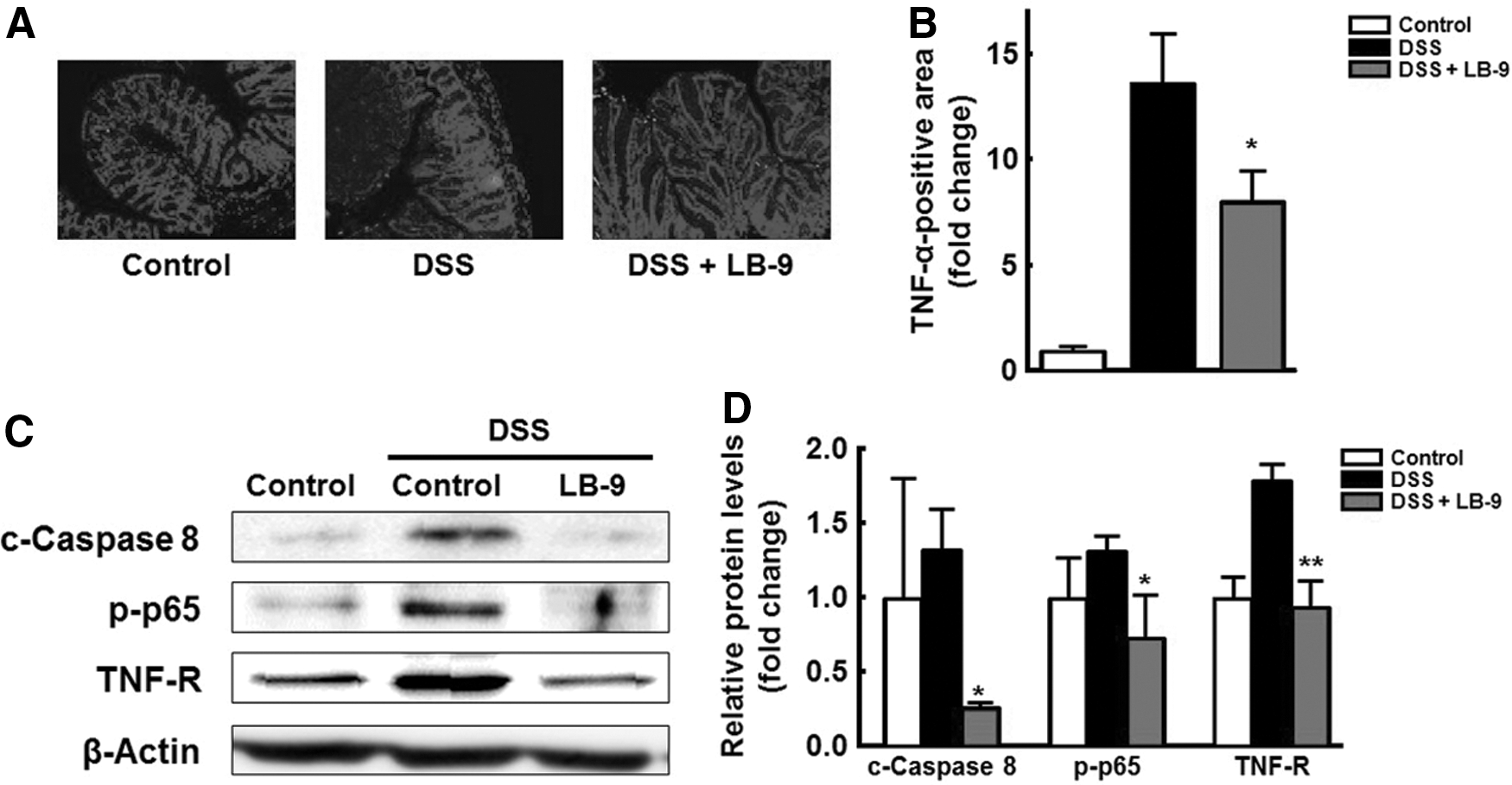
Effect of LB-9 on TNF-α-induced IEC apoptosis.
TNF-α promotes IEC apoptosis by death receptor-dependent caspase-8 activation and TNF receptor expression through activation of the NF-κB pathway in IECs. 27 Hence, we investigated the caspase-8 and NF-κB-mediated TNF receptor expression in colon tissues of mice with DSS-induced colitis, with or without LB-9 treatment. The activation of caspase-8, NF-κB, and the TNF receptor, was higher in the colon of DSS-treated mice, in comparison with the control (Fig. 3B). However, the induction of these key proteins related to TNF-α-mediated IEC apoptosis was significantly reduced compared with that observed in LB-9-treated control mice (Fig. 3B). A previous study suggested that mRNA expression and cytokine levels of TNF-α were significantly decreased after LAB treatment during DSS-induced colitis. 28 TNF-α levels were correlated to amelioration of DSS-induced colitis. 29
In conclusion, our findings indicate that treatment with LB-9 inhibits IEC apoptosis induced by DSS through downregulation of TNF-α production. We showed that LB-9 pretreatment protected against DSS-induced colitis by inhibiting the apoptotic death of IECs, presumably through the suppression of TNF-α production. Our study establishes a rationale for LB-9 administration as a therapeutic approach in the prevention of IBD. and in particular, the prevention of UC.
Footnotes
Acknowledgments
The authors acknowledge Dr. Ho Won Lee from the Biomedical Research Institute, School of Medicine, Kyungpook National University for providing technical assistance with in vivo bioluminescence imaging analysis. This research was supported by a grant from the Lotte R&D Center, Korea.
Author Disclosure Statement
No competing financial interests exist.