
Abstract
This study aimed to examine the antidiabetic effects of various concentrations of adlay bran oil (ABO) in high fat diet and streptozotocin-induced diabetic rats. Dietary supplementation with 10% ABO for 4 weeks effectively decreased the blood triacylglycerol, glucose, and total cholesterol levels in diabetic rats, although body weight remained the same. The mRNA and protein expressions of hepatic glucose transporter 2 (GLUT-2) and phosphoenolpyruvate carboxykinase (PEPCK) were increased and that of glucokinase (GCK) were decreased in diabetic rats. However, 10% ABO treatment reduced the mRNA and protein expressions of GLUT-2 and PEPCK and elevated the expression of hepatic GCK in diabetic rats. Thus, ABO enhanced hepatic glucose metabolism to decrease blood glucose in diabetic rats. In addition, 10% ABO supplementation increased the expression of phosphorylated protein kinase B (Akt) relative to the total Akt levels in the muscles of diabetic rats, indicating enhanced insulin sensitivity. The results indicate that ABO displays a potential for improving hyperlipidemia and hyperglycemia in diabetes by enhancing insulin sensitivity and hepatic glucose metabolism.
Introduction
Diabetes mellitus (DM) is an important metabolic disease, which has evoked enormous financial burden, and severely affects medical health care insurance policies in developed countries. The prevalence of DM is estimated to increase by 4.9% by 2030. 1 DM includes type 1 and type 2, both of which exhibit hyperglycemia. The prevalence of type 2 diabetes (T2D) is about 90%, of all diabetes, thereby accounting for most of the diabetic cases. 2,3 The key etiology of T2D is chronic inflammation associated with insulin resistance. 4 Due to insulin resistance, T2D patients are unable to transport glucose into the liver, muscle, and adipose cells, which in turn results in overproduction of insulin by pancreatic beta cells eventually leading to the dysfunction of the cells. 5,6 Moreover, insulin insensitivity mediates elevation of gluconeogenesis in the livers of T2D patients, 5 thus worsening the hyperglycemia caused by dysfunction in glucose uptake. Furthermore, it is known that 55% of T2D cases are associated with obesity, since insulin resistance, which hinders the uptake and use of glucose by the cells, results in increased breakdown of fats for meeting the energy requirement and the consequent hyperlipidemia. 7,8 That is why most T2D patients are also diagnosed with dyslipidemia, including reduced high density lipoprotein cholesterol levels, delayed catabolism of triacylglycerol (TG)-rich lipoprotein, increased postprandial TG level, and increased low density lipoprotein (LDL) production. A clinical trial demonstrated that diabetes patients with high TG and LDL are vulnerable to coronary heart disease. 9
Insulin regulates glucose homeostasis by regulating glucose uptake into skeletal muscles and adipose tissues. The glucose transporter (GLUT) permits the facilitated diffusion of glucose down its concentration gradient into cells upon insulin stimulation. 10 Liver also plays an important role in regulating postprandial blood glucose homeostasis or during fasting. 11 GLUT-2 can mediate bidirectional glucose transport, either in or out of hepatocytes, after insulin stimulation. 12 High plasma insulin concentration stimulates the transport of glucose into the hepatocytes after a meal. Conversely, during fasting, hepatocytes produce glucose (gluconeogenesis), which is transported into the blood from the liver. 10
Enzymes such as glucokinase (GCK) and phosphoenolpyruvate carboxykinase (PEPCK) are rate-limiting enzymes for hepatic glucose metabolism. Hepatic GCK is involved in glycolysis and glycogenesis, leading to a decrease in blood glucose levels. 13 PEPCK, on the other hand, responds to the lack of glucose in the cells, resulting in de novo glucose synthesis. 14
The Akt signaling pathway regulates the GLUT-4 mediated glucose transportation into skeletal muscles and adipocyte tissues and inhibits the activity of glucose synthase. 15 The Akt-phosphatidylinositol 3-kinase (PI3K) signaling pathway regulates blood glucose metabolism and thus contributes to insulin resistance. 16
Although several oral or injectable antidiabetic drugs are largely used to control blood glucose levels in T2D patients, they can have serious adverse side effects. 17 Therefore, finding other potential therapeutic candidates from natural sources for treating diabetes has become a main trend. Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) has been used as both food and medicine in Asian countries and exhibits anti-inflammatory and antioxidative effects. 18 A previous study has shown that adlay bran oil (ABO) could decrease oxidative stress, blood lipids, LDL-cholesterol, and plasma insulin levels in rats through the actions of its phytosterol and polyphenolic compounds. 19 –21
A combined treatment with high fat diet (HFD) and low-dose streptozotocin (STZ) in mice or rats produces the symptoms of hyperlipidemia, insulin resistance, and hyperglycemia and can be used to induce T2D in animal models. 22 –24 Until now, the effects of ABO on T2D are unknown. In this study, we investigated the effects of ABO on blood glucose, blood lipid levels, and related metabolic pathways in HFD and low-dose STZ induced T2D rats.
Materials and Methods
Preparation of adlay oil
Adlay (Coix lachryma-jobi L. var. ma-yuen Stapf, Job's tears) seeds of Taichung Shuenyu 4 were obtained from Taichung City in Taiwan. The adlay brans were separated from air-dried seeds according to the standard procedure. 25 ABO was extracted from 1 kg adlay brans for 2 h using an extruder, yielding 200 mL adlay oil, which was then stored at −20°C and kept in the dark until use.
Animals and diets
Male Sprague-Dawley rats (6-week old) were purchased from BioLASCO Taiwan Co. (Taipei, Taiwan, R.O.C.). Animal care and experimental protocols were approved by the Institutional Animal Care and Use Committee of National Taiwan University (NTU-IACUC/protocol 125/2013). HFD was composed of commercial chow diet with increased fat content (60% of total calories). One kilogram of HFD contained 390.6 g lard and 43.4 g soybean oil. The STZ solution (Sigma Chemical Co., St. Louis, MO, USA) was prepared by dissolving the powder in citrate buffer (pH 4.5). The rats were either fed chow diet in the normal control (NC) group (n = 8) or HFD (n = 24). After 2 weeks, the HFD-induced rats were randomly divided into three groups (n = 8 each) as follows: HFD + STZ group, HFD + STZ + ABO-L group, and HFD + STZ + ABO-H group. For ABO-L group and ABO-H group, HFD was supplemented with 5% and 10% (w/w) ABO, respectively. These groups continued to receive HFD or HFD containing ABO for 4 and 3 weeks after ABO treatment; all the HFD-fed rats received 35 mg STZ/kg body weight by intraperitoneal injection to induce high blood glucose. Blood was withdrawn from tail veins, and fasting blood glucose (FBG) was monitored using a blood glucose meter (OK Biotech Company, Hsinchu, Taiwan) every week along with body weight; total food and water intakes were recorded. At the end of the experiment (day 42), all the rats were fasted for 12 h and then sacrificed by cardiac puncture, and the serum, liver, and muscles were harvested for further experiments.
Biochemical analysis
Blood was withdrawn every week from the tail vein of rats after fasting for 12 h. Biochemical parameters, including the levels of total cholesterol (TC) and TG, were determined in the serum samples by biochemical analyzer (Cayman Chemical Company, Ann Arbor, MI, USA).
RNA isolation and quantitative reverse transcription–polymerase chain reaction
Total RNA was extracted from the target tissues using TriPure isolation reagent (Roche Applied Science, Penzberg, Upper Bavaria, Germany), and cDNA was prepared from RNA using a kit (Takara Bio, Inc., Otsu, Shiga, Japan) according to the manufacturer's protocol. The cDNA was used for quantitative reverse transcription–polymerase chain reaction (qRT-PCR) using KAPA SYBR® FAST qPCR Kit (Roche, Basel, Switzerland). The primers of Rat-18s (housekeeping gene), Rat-Glut-2, Rat-PEPCK, and Rat-GCK for qRT-PCR are listed in Table 1. The relative expression of genes was calculated using 2−ΔΔCt method. 26
Primers Used in Quantitative Reverse Transcription–Polymerase Chain Reaction
PEPCK, phosphoenolpyruvate carboxykinase.
Western blotting
Protein extraction was performed from the liver and muscles as published previously. 27 The lysates (15 μg total protein) were electrophoresed on 12% Tris-Glycine sodium dodecyl sulfate–polyacrylamide gel and transferred to polyvinylidene fluoride membrane (Bio-Rad Laboratories, Hercules, CA, USA). The membranes were incubated at 4°C overnight with primary antibodies against glyceraldehyde-3-phosphate dehydrogenase (Abcam, Cambridge, MA, USA), phosphorylated AKT (Cell Signaling, Danvers, MA, USA), AKT (Cell Signaling), GLUT-2 (Millipore, Burlington, MA, USA), PEPCK (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and GCK (Proteintech®, Chicago, IL, USA). The membranes were then incubated with horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse immunoglobulin G (Cell Signaling) at 25°C for 2 h. The membranes were probed with enhanced chemiluminescence detection reagent and revealed by UVP, a luminescence imaging system (Thermal Fisher Scientific, Waltham, MA, USA).
Statistical analysis
Each result was calculated as mean ± standard deviation. The GraphPad Prism 6 software (GraphPad Software, Inc., La Jolla, CA, USA) was used for statistical analyses using one-way analysis of variance and Duncan's multiple range test or Student's t test, with P < .05 considered significant.
Results
Effects of ABO on body weight, food intake, and tissue weight in rats
Throughout the experiment, all the rats were healthy and showed no pathological signs or abnormalities. Although the final body weight of rats in HFD + STZ, HFD + STZ + ABO-L, and HFD + STZ + ABO-H groups did not differ from NC group (Table 2), the daily food intake of rats in HFD + STZ group had significant reduction compared with NC group, whereas the absence of difference in food intake among HFD + STZ, ABO-L, and ABO-H groups revealed a better food efficiency among these groups (Table 2). At the end of the sixth week, rats were sacrificed, and the adipose tissue weight was measured. However, ABO supplementation had no effect on the weight of adipose tissues (data not shown).
Effect of Different Diets on Body Weight, Food Intake, and Feed Efficiency in Rats After Six Week Experimental Period
The results are expressed as mean ± SD for 8–10 rats.
Feed efficiency (%) = (body weight gain/total food intake) × 100.
The values not sharing the same superscripts are significantly different (P < .05). Significance levels were analyzed by one-way ANOVA and Duncan's multiple range test.
NC, normal control; HFD + STZ, high fat diet + streptozotocin; HFD + STZ + ABO-L, high fat diet + streptozotocin +5% adlay bran oil; HFD + STZ + ABO-H, high fat diet containing 10% adlay bran oil; SD, standard deviation; ANOVA, analysis of variance.
Effects of ABO on blood lipid and FBG in HFD and STZ-treated rats
The treatment with HFD and STZ in rats resulted in higher blood TC and TG levels compared with rats in the NC group, but the levels were restored effectively in the ABO-H group (Table 3). ABO-L rats also exhibited significantly lower TC levels, but blood TG concentrations were similar to those in HFD + STZ rats. However, 10% ABO supplementation effectively reduced both blood TC and TG levels compared with HFD and STZ treatment.
Changes in Blood Lipids and Blood Glucose of Rats Fed with Different Diets for Six Weeks
Results are expressed as mean ± SD for 8–10 Sprague-Dawley rats.
Values with different superscripts are statistically different (P < .05). Significance values were analyzed by one-way ANOVA and Duncan's multiple range test.
We also investigated the effect of ABO on HFD and STZ-induced hyperglycemia. One shot of STZ induced a 2.45-fold increase in blood glucose levels compared to rats in the NC group after the sixth week. However, the ABO-H group, but not ABO-L group, displayed significantly lower blood glucose compared to HFD + STZ rats (P < .05), indicating that supplementation of 10% ABO improves blood glucose in rats with diabetes (Table 3).
Effects of ABO on glucose metabolism in the liver
To investigate the molecular mechanism of the hypoglycemic effect of ABO, we determined the mRNA expression levels of GLUT-2, GCK, and PEPCK in the liver and normalized them to the 18S mRNA levels. The expression of hepatic Glut2 gene in rats of both ABO-L group and ABO-H group was significantly decreased compared to the rats in HFD + STZ group (P < .05; Fig. 1). Besides, the HFD + STZ group showed a decrease in the expression of the GCK gene compared with NC group that was rescued in the ABO-H group (P < .05; Fig. 1). Similar recovery was observed in PEPCK gene expression, a key enzyme for gluconeogenesis (P < .05; Fig. 1). We next examined the expression of hepatic GLUT-2, PEPCK, and GCK proteins in rats after the 4-week ABO treatment. The STZ + HFD-treated rats expressed elevated protein levels of GLUT-2 and PEPCK (P < .05; Fig. 2A, B), which were rescued by 10% ABO treatment. Similarly, the expression of GCK protein was suppressed upon exposure to HFD + STZ to nearly undetectable level compared with NC; however, ABO treatment significantly increased GCK protein expression (Fig. 2C), consistent with the pattern of corresponding mRNA expression.
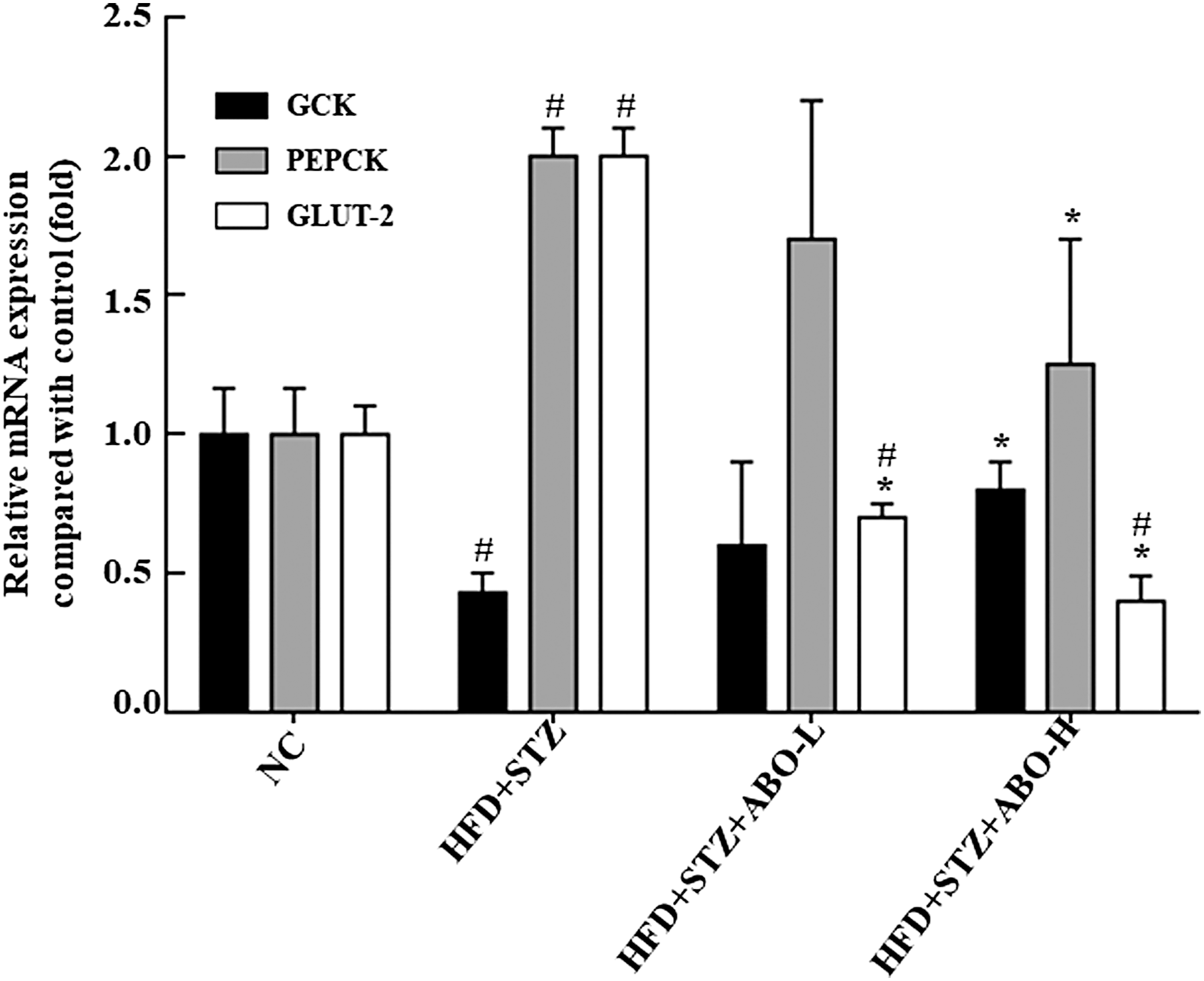
Effect of ABO on the expression of hepatic glucose transporter gene and on glucose metabolism. The gene expressions of hepatic Glut-2, GCK, and PEPCK in rats were determined by quantitative reverse transcription–polymerase chain reaction, and 18s was used as an internal control. Treatment with 5% and 10% ABO decreases the higher Glut-2 gene expression level in HFD+STZ treated rats, and 10% ABO decreases the higher PEPCK gene expression level and increases the lower GCK expression level in HFD + STZ treated rats. The results are expressed as mean ± SD for three rats in each group. Significance was analyzed using Student's t-tests. #Indicates significant difference (P < .05) when comparing each group with NC group. *Indicates significant difference (P < .05) when comparing HFD + STZ + ABO-H or HFD + STZ + ABO-L with HFD + STZ group. NC, normal control; HFD + STZ, high fat diet + streptozotocin; HFD + STZ + ABO-L, high fat diet + streptozotocin +5% adlay bran oil; HFD + STZ + ABO-H, high fat diet containing 10% adlay bran oil; GLUT-2, glucose transporter-2; GCK, glucokinase; PEPCK, phosphoenolpyruvate carboxykinase; SD, standard deviation.
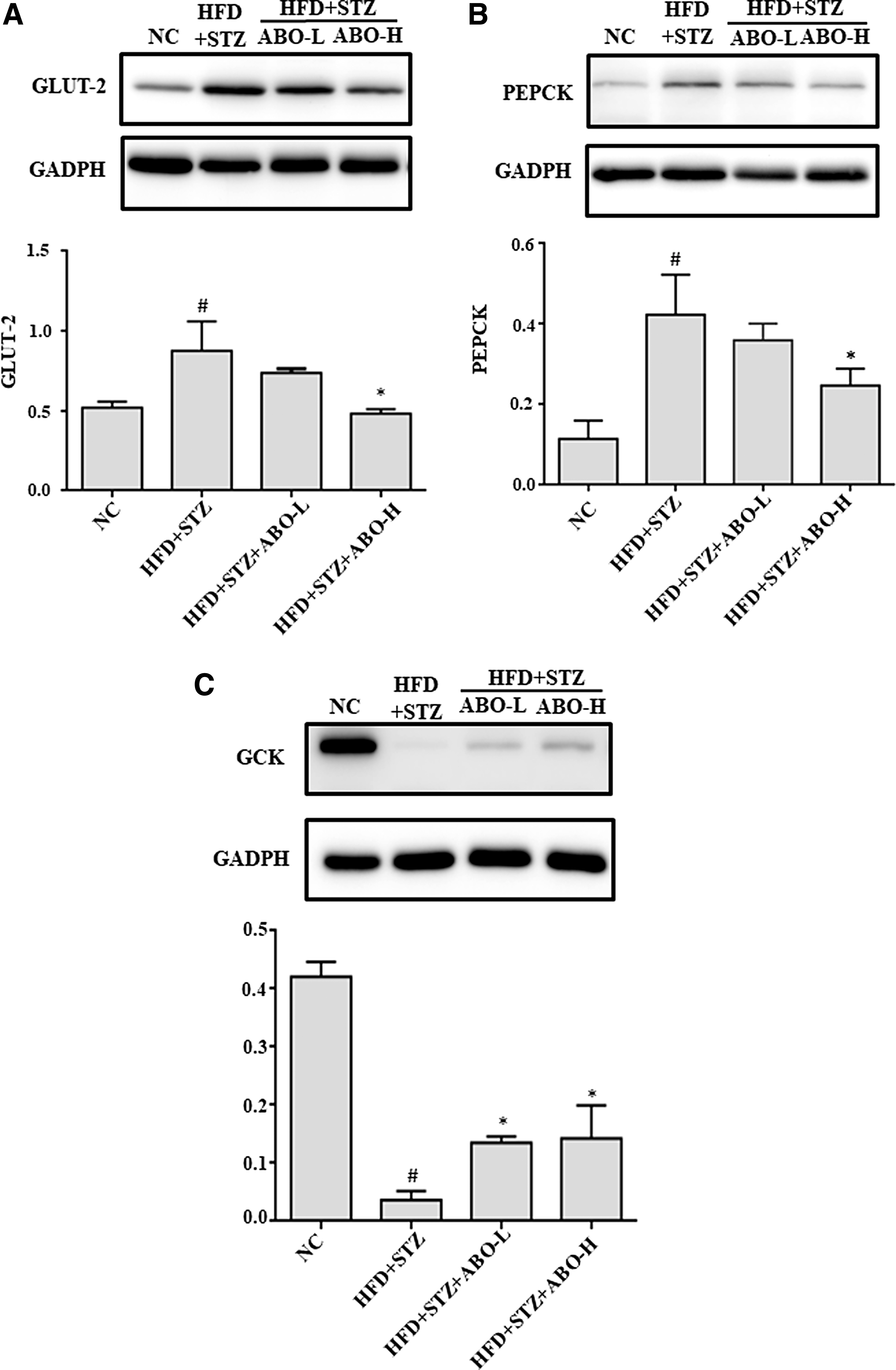
Effect of adlay bean oil on the protein expressions of glucose transport and glucose metabolism in liver. The protein expressions of hepatic
Effects of ABO on insulin-signaling transduction in muscles
We further examined the insulin-sensitizing effect of ABO by investigating the AKT pathway, which is known to be involved in insulin signaling. As shown in Figure 3, the ratio of phosphorylated Akt to total Akt levels in the muscles of ABO-H group rats was increased compared to the HFD + STZ rats, suggesting that the antihyperglycemic effects of ABO might involve reactivation of the signal transduction pathway of insulin.
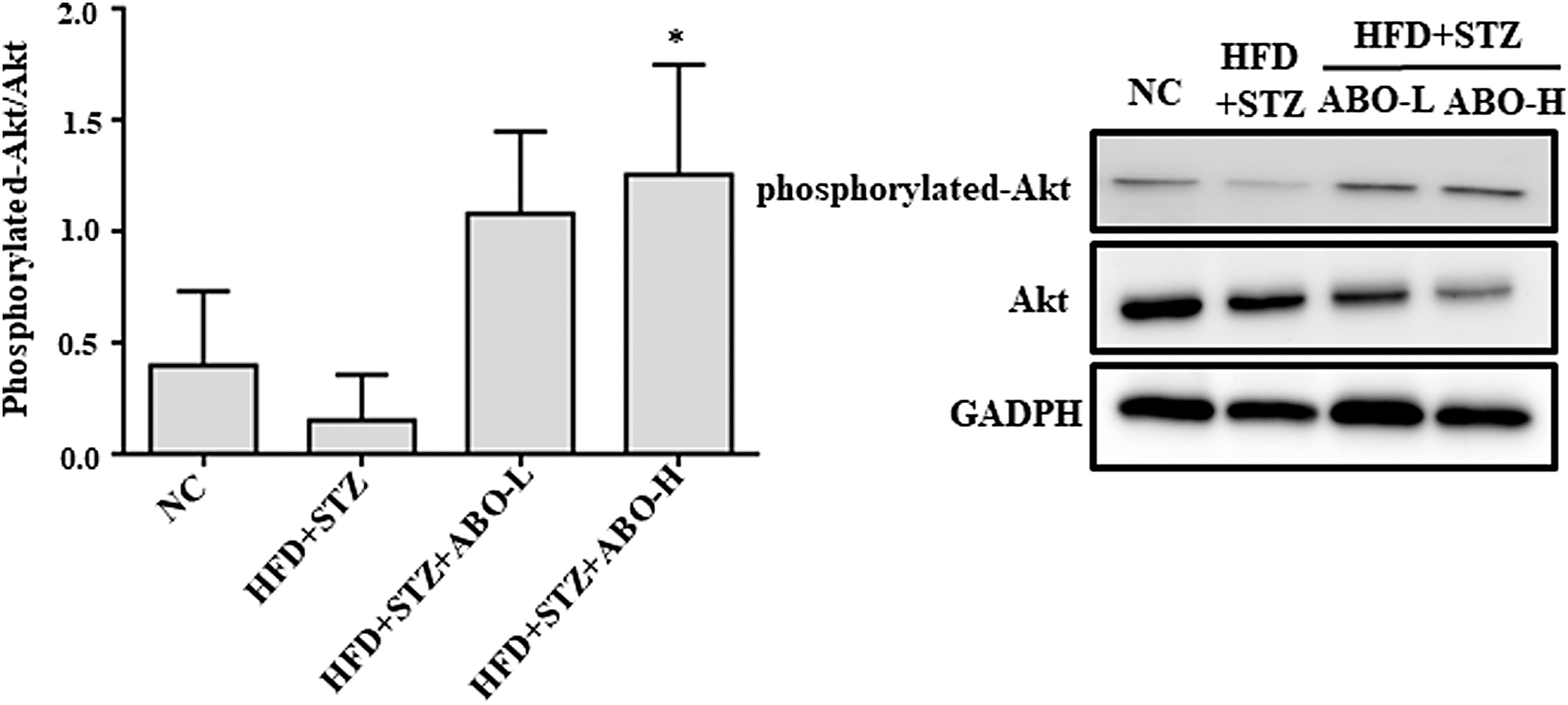
Effect of ABO on the protein expression of insulin signaling in muscles. The protein expression of phosphorylated Akt/total Akt in rats was determined by western blotting, and GAPDH was used as a loading control. Treatment with 10% ABO enhances the ratio of phosphorylated-Akt/Akt protein expression. The results are expressed as mean ± SD for three rats in each group. Significant difference was analyzed by Student's t-tests. *Indicates significant difference (P < .05) when comparing HFD + STZ + ABO-H with HFD + STZ group.
Discussion
Our results in the current study indicate that supplementation of the diet with ABO reduces blood lipids and glucose in diabetic rats. In the previous study, we compared the effect of different oil supplements on plasma lipids and concluded that the ABO supplementation resulted in much lower blood TG and LDL levels compared to the lard control and substituting half of the lard with ABO was very effective. 19
In this study, we demonstrate that ABO lowers the high blood TG and cholesterol levels induced by HFD and STZ treatment in a dose-dependent manner. Interestingly, we did not observe significant differences in the body weight nor weight of adipose tissues, indicating that supplementation of diet with ABO did not result in any extra fat accumulation. A high ratio of polyunsaturated fatty acids (PUFAs) to saturated fatty acids (SFAs) decreases the accumulation of body fat, plasma TG, and TC in HFD-fed hamsters. 28 Indeed, ABO exhibited a higher PUFA/SFA ratio (2.44%) compared to lard (0.39%). 19 Besides, the phytosterol compounds in ABO, including camperstanol and β-sistosterol, decrease serum TC levels in animals by blocking cholesterol absorption in the intestine and inhibiting cholesterol synthesis in the liver. 20,29,30
In this study, the combination of HFD and low-dose STZ treatment was used to mimic late stage of T2D. 31 Although STZ treatment has been previously associated with decreasing body weight, 32 we did not observe any significant weight loss after STZ treatment. The absence of any effect on body weight might be attributed to the low dose of STZ (35 mg/kg body weight) used in this study instead of high doses used elsewhere (>50 mg/kg body weight). We observed a significant increase in the body weight in HFD-induced rats at week 4; however, the STZ injection decreased the body weight gradually. Thus, the body weights of control and HFD + STZ groups were comparable at the end of the experiment.
STZ induction has been shown to cause a decreased level of insulin secretion due to damage to pancreatic beta cells, leading to insulin deficiency and subsequent dysfunction of glucose and lipid catabolism. 33,34 Insulin is reported to bind to insulin receptor in insulin sensitive tissues, such as muscles, and mediate tyrosine phosphorylation and recruitment of insulin receptor substrates (IRS). The activation of IRS further triggers phosphorylation of Akt by PI3K. 35 Phosphorylated Akt stimulates glucose uptake in muscles through GLUT-4 modulation. 36 In this study, we found that 10% ABO treatment increased the levels of phosphorylated Akt, suggesting that ABO improves the insulin sensitivity of muscles and, thus, helps in the regulation of blood glucose levels.
Animal models of diabetes exhibit an increased expression of PEPCK protein and a decreased level of GCK protein in the liver. 37 –39 A similar phenomenon was observed in our diabetic animal model, and diet supplementation with 10% ABO restored the expression levels of these genes at both the mRNA and protein levels, thus decreasing de novo glucose synthesis and enhancing the hepatic glycolysis and glycogenesis. An increased expression of GLUT-2 exacerbates the high blood glucose in T2D animals by catalyzing the glucose efflux. 10 ABO supplementation decreases hepatic GLUT-2 expression, which might contribute to its hypoglycemic effect. The polyphenolic substances in adlay bran, including ferulic acid, p-coumaric acid, and syringic acid, might contribute to its strong antioxidation activities. 40 Previous studies demonstrated that ferulic acid–treated diabetic rats displayed reduced hepatic GLUT-2 expression. 41 The syringic acid increases plasma insulin concentrations in diabetic rats, resulting in lowering of blood glucose. 42 Furthermore, we reported earlier that the flavonoids in adlay bran offer anti-inflammatory effects. It is now clear that both inflammation and oxidative stress contribute to the dysfunction of insulin. 43,44 Thus, compounds with the antioxidant and anti-inflammatory characteristics could be beneficial in restoring the function of insulin.
In conclusion, we show that ABO has the potential to reduce the concentration of blood lipids and glucose in diabetic rats through the modulation of glucose production and glucose output in the liver. In addition, ABO increases insulin sensitivity and the downstream signaling pathway, which may further contribute to the amelioration of T2D.
Footnotes
Acknowledgment
This study was supported by the funding from the Ministry of Science and Technology of Taiwan (MOST 104-2320-B-002-039).
Author Disclosure Statement
No competing financial interests exist.