
Abstract
Osteoporosis is a systemic skeletal disease that causes bone weakness and fragility. Consuming bone-beneficial nutrients through diet can prevent and treat osteoporosis. Acer palmatum (Japanese maple) leaves are used to make tea, but there have been few reports of their health benefits, especially regarding bone homeostasis. In this study, we evaluated the effects of A. palmatum hot water extract (APE) on osteoclastogenesis and osteoblastogenesis in cultured cells. APE suppressed the number of tartrate-resistant acid phosphatase-positive multinucleated osteoclasts in RANKL induced RAW264.7 cells. Furthermore, APE facilitated Alkaline phosphatase activity and calcium deposition during osteoblast differentiation in MC3T3–E1 cells. High-performance liquid chromatography analysis was performed to investigate the effective components of APE, and four flavonoids orientin, isoorientin, vitexin, and isovitexin were identified with the LC-MS analysis. Treatment with fractionated APE suppressed osteoclastogenesis and facilitated osteoblastogenesis in cultured cells. These findings suggest that APE contains antiosteoporotic compounds; thus, APE might have health promoting effects that help prevent osteoporosis by inhibiting osteoclastogenesis and facilitating osteoblastogenesis.
Introducton
In the human body, bones are continuously broken down and restructured. Bone metabolism is maintained by the balance of bone resorption by osteoclasts and bone formation by osteoblasts. 1,2 Osteoclasts break down bones, which are crucial for the maintenance, repair, and remodeling of bone tissue. Osteoclast precursor cells are monocytic and fuse to form multinucleated osteoclasts during differentiation. Mature osteoclasts digest proteins and minerals at the molecular level by secreting the enzyme tartrate resistant acid phosphatase (TRAP). Osteoblasts express the receptor activator of nuclear factor (NF)-κB ligand (RANKL), which activates the RANK receptor to regulate the differentiation, activation, and survival of osteoclasts and their precursor cells. 3 In contrast, osteoblasts synthesize bones during bone formation. 4,5 The osteoblast precursor migrates from blood vessels to the bone surface and is absorbed by osteoclasts. Mature osteoblasts secrete collagen to induce the deposition of calcium, phosphate, and other ions that form new bone matrix. This mineralized bone is the main support in vertebrates and provides important mineral storage for physiological homeostasis. The mineralization of osteoblasts using cultured cells is used to evaluate the effects of drug treatment on bone formation. 6,7
Decreased bone mass resulting from an imbalance of osteoclast and osteoblast activities can lead to osteoporosis. 8 Osteoporosis is a severe disease that causes bones to become weak and brittle, increasing the risk of broken bones. 9 In Japan, 13 million people are estimated to be suffering from osteoporosis, with only 20% of sufferers receiving treatment. 10 Pharmaceutical drugs have been developed and widely used to cure osteoporosis in a robust manner. However, some antiosteoporosis drugs were reported to have serious side effects, such as osteonecrosis of the jaw and abnormal fracture of the femur. 11 Because osteoporosis is highly undertreated, reducing its onset would be preferable to treatment. Improved dietary habits and moderate exercise are effective for preventing osteoporosis. Although a nutritionally balanced diet is necessary, some bone-beneficial compounds are recommended for consumption to aid bone metabolism. These compounds, including calcium, vitamin D, and vitamin K, activate osteoblastogenesis or inhibit osteoclastogenesis. Therefore, the prevention of osteoporosis through diet management has been widely promoted. 12 Various health-promoting foods have attracted attention for preventing osteoporosis, and the development of antiosteoporotic foods is desirable. Many functional foods have been developed for their health-promoting effects. For example, green tea extract was reported to protect against bone loss by inhibiting osteoclast differentiation. 13 The main active components in tea extract are catechins, particularly epigallocatechin gallate, and the antiosteoporotic effects of tea extract have being revealed. 14,15
To confirm the potential of broad-leaf tea, we focused on the health-promoting effects of the leaf extract of Acer palmatum (Japanese maple). This broad-leaf tree is among the most popular maple trees, called irohamomiji in Japan, and its leaves are used to prepare tea, although few reports have focused on its cellular effects. Previously, A. palmatum extract has been reported to contain different flavones such as vitexin, which have various positive effects. 16 Despite its potential as a health-promoting food, studies are lacking on the cellular effects of A. palmatum.
In this study, we investigated the effects of hot water extract from A. palmatum (APE) on bone metabolism in cultured cells. As increased osteoclastogenesis and decreased osteoblastogenesis are major indicators of bone loss, we evaluated the cellular effects of APE at different doses on osteoclast differentiation in RAW264.7 cells and osteoblast differentiation in MC3T3-E1 cells. Furthermore, four flavonoids were identified in APE by LC-MS analysis.
Materials and Methods
Preparation of tea leaves and A. palmatum extract
A. palmatum leaves were used to make tea leaves and to prepare APE according to the following procedure (shown in Fig. 1A, B). A. palmatum autumn leaves, cultivated in Mie Prefecture, Japan, were washed and dried. The leaves were hand rolled at 15–20°C for 1 h and underwent endogenous enzymatic fermentation at 15–20°C for 12 h. Then, the leaves were air-dried for 12 h. The process was based on tea manufacture. The dried leaves (29.3 g) were then finely ground by a jet mill. The ground leaves (18.0 g) were extracted with hot water at 80°C for 10 min. The mixture was centrifuged (13,000 g, 15 min), and the supernatant was collected. After removing the water by evaporation, the remaining dry matter (3.8 g) was used as the APE in this study. A stock solution was prepared by dissolving APE in ultrapure water at 5 mg/mL.
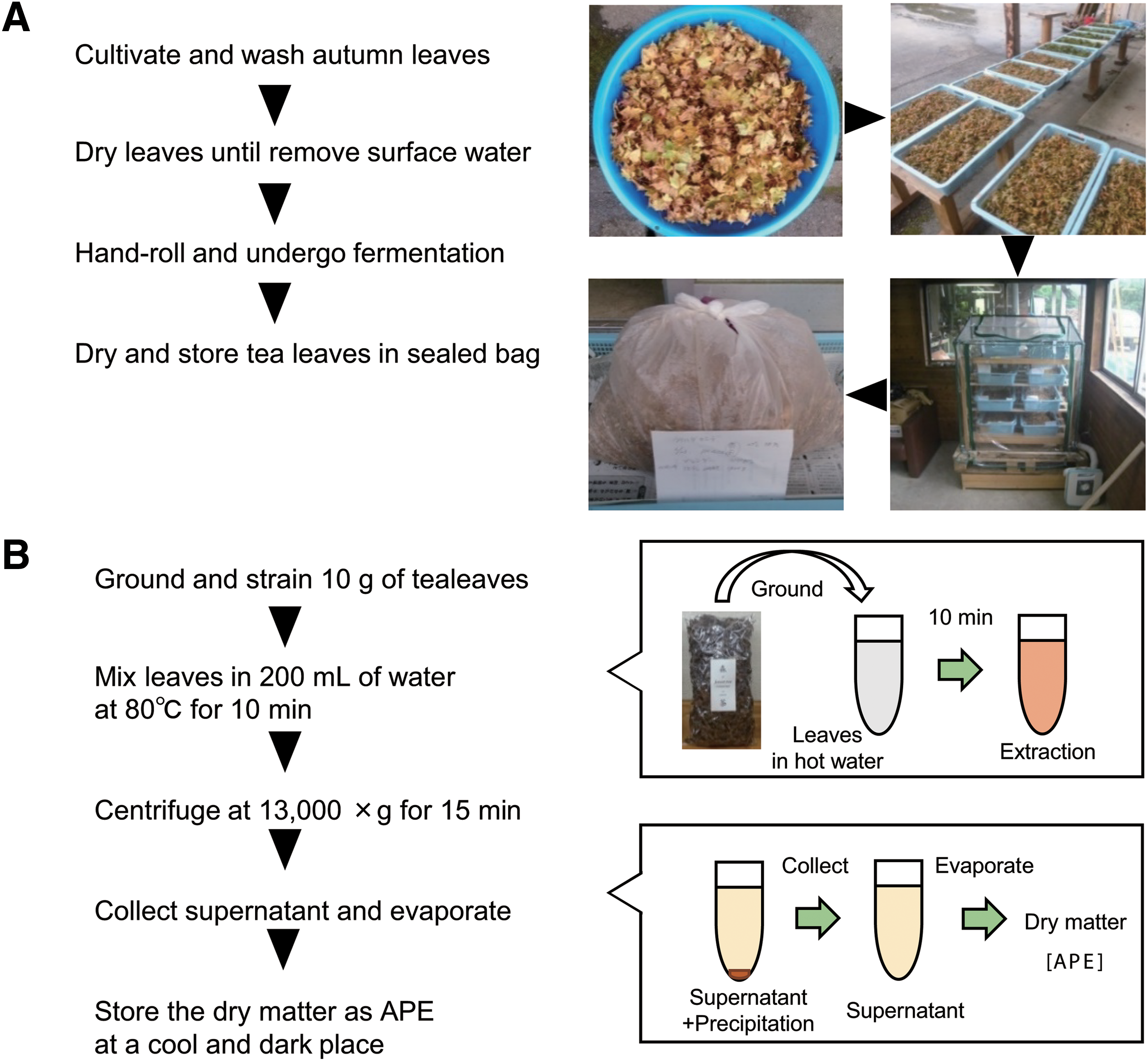
Preparation and extraction of APE from A. palmatum leaves.
High-performance liquid chromatography analysis, fractionation, and LC-MS analysis
APE was analyzed and fractionated by high-performance liquid chromatography (HPLC; JASCO Corporation, Tokyo, Japan) equipped with a C18 column (5C18-AR-II, 20ID × 250 mm; Nacalai Tesque, Kyoto, Japan), a UV detector at 220 nm (Jasco UV-1570; JASCO Corporation), and using a mobile phase of water and methanol (0–20 min; water, 20–60 min; water to methanol, 60–70 min; methanol) at a flow rate of 5 mL/min. For fractionation, 1 mL of the 5 mg/mL APE stock solution was injected. LC-MS analysis (LCMS-2010; Shimadzu, Tokyo, Japan) was carried out in the positive and negative ion mode using electrospray ionization.
Cell culture and reagents
RAW264.7 cells (American Type Culture Collection, Manassas, VA, USA, passages 6–8) were grown in alpha-modified minimum essential medium (αMEM; Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Biowest, Nuaillé, France) at 37°C under 5% CO2. A cell scraper was used to harvest the RAW264.7 cells. MC3T3E-1 cells (RIKEN Cell Bank, Tsukuba, Japan, passages 6–8) were grown in αMEM and supplemented with 10% FBS at 37°C under 5% CO2. 0.05% Trypsin/EDTA (Nacalai Tesque) was used to harvest the MC3T3-E1 cells. Cells were cultured until they reached 80% confluence before use in each assay. The reagents orientin (Sigma-Aldrich, St. Louis, MO, USA), isoorientin (Abcam, Cambridge, MA, USA), vitexin (Sigma-Aldrich), and isovitexin (Wako, Osaka, Japan) were used in the experiments.
Cell viability assay
RAW264.7 cells (4.0 × 103 cells/well; 96-well plate; individual well area, 0.34 cm2) and MC3T3-E1 cells (2.0 × 103 cells/well; 96-well plate) were cultured for 24 h. Cells in the exponential growth phase were treated with APE at concentrations of 0.01–10 μg/mL for 24 h. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT; Nacalai Tesque, Kyoto, Japan) solution (5 mg/mL in ultrapure water) was then added to each well, and the plate was incubated for a further 4 h at 37°C. Formazan crystals were dissolved in 100 μL of 0.01 M HCl/10% SDS. The absorbance was quantitated as the percentage compared with the untreated control using a Microplate Reader (CHROMATE4300; practical Japan, Chiba, Japan) at 600 nm.
Osteoclast differentiation
RAW264.7 cells (2.0 × 103 cells/well; 96-well plate) were cultured for 24 h. During osteoclast differentiation, the cells were treated with receptor activator of nuclear factor κ-B ligand (RANKL) (Oriental Yeast, Tokyo, Japan) and macrophage colony-stimulating factor (M-CSF; Affymetrix Japan, Tokyo, Japan) at final concentrations of 50 and 10 ng/mL, respectively. The cells were simultaneously treated with APE at final concentrations of 0.01–10 μg/mL or 5 μM of each flavonoid. After 3 days, the medium was replaced with fresh medium containing the same reagent, and the plate was incubated until day 5.
TRAP staining
RAW264.7 cells were washed with PBS and fixed with 10% formalin for 10 min. Washed cells were stained with TRAP staining solution 17,18 for 60 min at 37°C. TRAP-positive osteoclasts with three or more nuclei in the well were considered mature osteoclasts.
Osteoblast differentiation
MC3T3-E1 cells (2.0 × 10 4 cells/well; 96-well plate for Alkaline phosphatase (ALP) assay, or 1.0 × 10 5 cells/well; 24-well plate; individual well area, 1.86 cm2) were cultured for 48 h. During osteoblast differentiation, the cells were treated with 5 mM β-glycerophosphate and 50 μg/mL ascorbic acid. The cells were simultaneously treated with APE at final concentrations of 0.01–10 μg/mL or 5 μM of each flavonoid. Every 3 days the medium was replaced with fresh medium containing the same reagent until the assay was finished.
ALP activity assay
The ALP activity assay was performed using day 3 or day 7 MC3T3-E1 cells. ALP activity was measured with an ALP Assay Kit (Takara, Shiga, Japan) according to the manufacturer's protocol. The absorbance was quantified using a Microplate Reader (CHROMATE4300) at 405 nm, as the percentage compared with the vehicle treated control.
ALP staining
The ALP staining was performed using day 7 MC3T3-E1 cells with a TRAP/ALP Staining Kit (Wako) according to the manufacturer's protocol. In brief, cells were fixed with 4% paraformaldehyde for 10 min and permeabilized with acetone/ethanol (50/50) for 1 min. After washing with PBS, cells were stained with ALP staining solution for 30 min at 37°C. The solution was replaced with ultrapure water (UPW) before imaging.
Alizarin red staining
Alizarin red staining was performed at day 28. MC3T-E1 cells were washed with PBS and fixed with 10% formalin for 10 min. Washed cells were stained with 1% alizarin red S (pH 6.37; Waldeck Gmbh & Co. Kg., Münster, Germany) for 5 min at room temperature. The staining solution was then removed, and the degree of calcium deposition was quantitated using the captured images. The integrated density was calculated for each sample using ImageJ software (
Microscopy
An inverted microscope (IX51; Olympus, Tokyo, Japan) equipped with a digital camera (DP26; Olympus) was used to image the TRAP-stained osteoclasts, ALP-stained osteoblasts, and alizarin red-stained osteoblasts.
Statistical analysis
All data are expressed as the mean ± SD of the results from three different cultures. The results were analyzed using the Student's t-test. Values of P less than .05 and .01 were considered statistically significant. At least two independent experiments of triplicate samples were performed to verify results.
Results
Cell viability assay of APE-treated cells
Hot water extract of A. palmatum was prepared according to the indicated procedure (Fig. 1A, B, see Materials and Methods section). We examined the cell toxicity of APE at final concentrations of 0.001–10 μg/mL. Then, an MTT assay was used to measure cell viability from the mitochondrial activity of the two cell lines, RAW264.7 and MC3T3-E1. Compared with the untreated control, both cell lines showed that APE was not toxic within this concentration range (Fig. 2A, B). Therefore, APE was added to cells at this concentration range in the following experiments.
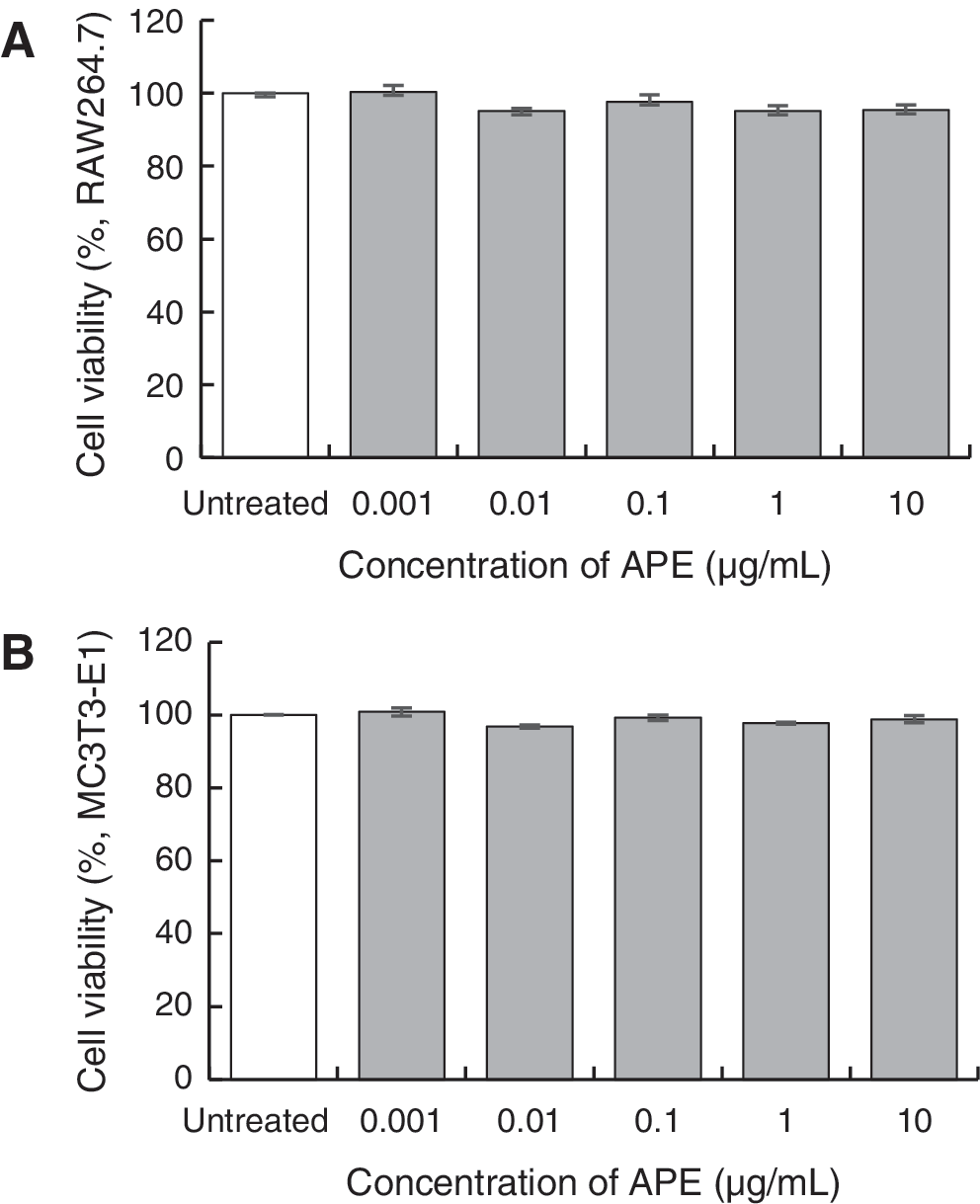
Effect of APE on cell survival. RAW264.7 cells
Effect of APE on osteoclast differentiation
Next, we evaluated whether APE affected RANKL-induced osteoclastogenesis using RAW264.7 cells. Under RANKL-treated conditions, the RAW264.7 cells repeated cell fusion to differentiate into multinucleated cells. Concurrent with RANKL and M-CSF treatment, the cells were cultured with APE at concentrations of 0.01, 0.1, 1, and 10 μg/mL for 5 days (Fig. 3A). TRAP-positive multinucleated cells with more than three nuclei were counted as mature osteoclasts (Fig. 3B). Therefore, TRAP-positive multinucleated cells were significantly decreased by treatment with 10 μg/mL of APE.

Effect of APE on osteoclast differentiation.
Effect of APE on osteoblast differentiation
We evaluated the cellular effects of APE on osteoblastogenesis using MC3T3-E1 cells. Concurrent with β-glycerophosphate and ascorbic acid treatment, cells were cultured with APE at concentrations of 0.01, 0.1, 1, and 10 μg/mL for 28 days to investigate osteoblastogenesis. Calcium deposition was quantified from the degree of osteoblastogenesis after alizarin red staining (Fig. 4B, C). Calcium deposition was significantly higher in cells treated with 1–10 μg/mL of APE compared with vehicle treated control. Next, cells were cultured with APE at concentrations of 0.01, 0.1, 1, and 10 μg/mL for 3 or 7 days to investigate ALP activity (Fig. 4D). The relative absorbance was significantly higher in cells treated with 10 μg/mL of APE. ALP staining was performed to confirm the effect of APE treatment for 7 days, and an obvious increase of ALP-stained area was observed (Fig. 4E).
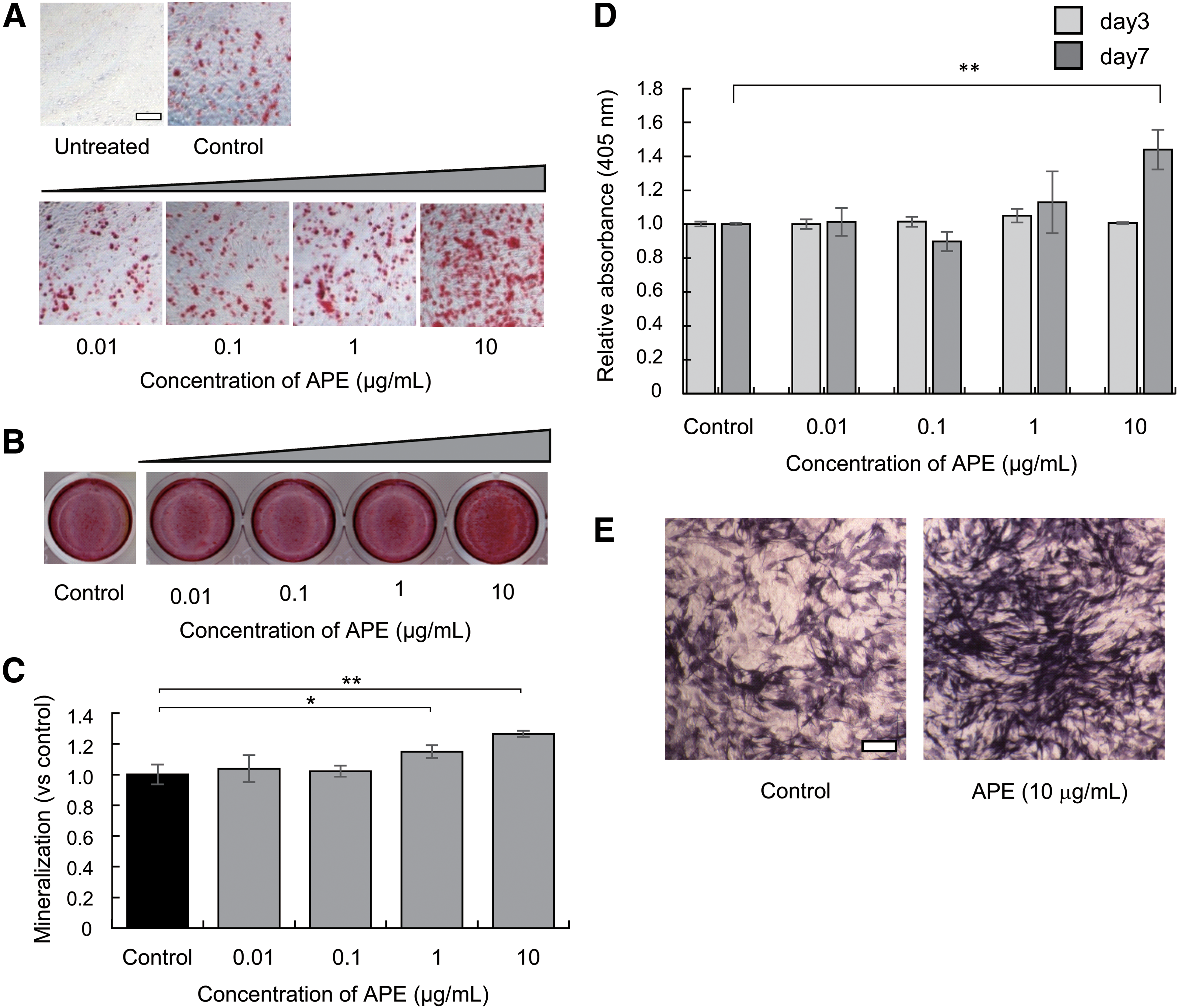
Effect of APE on osteoblast differentiation.
HPLC analysis and fractionation of APE
To identify the active component in APE, we analyzed APE by HPLC with UV detection at 220 nm (Fig. 5A). APE showed four major peaks (compounds i, ii, iii, and iv). LC-MS analysis was performed to determine the molecular weight (MW) of the compounds in APE (Fig. 5B).The four peaks showed the ion peaks at [M+H]+ = 449, 449, 433, and 433 (positive ion mode) and at [M-H]− = 447, 447, 431, and 431 (negative ion mode), respectively, by mass spectra (Supplementary Figures S1–S4). Thus, MW of compounds i, ii, iii, and iv was 448, 448, 432, and 432, respectively. We identified these four peaks as i = orientin, ii = isoorientin, iii = vitexin, and iv = isovitexin using commercial products as standards.
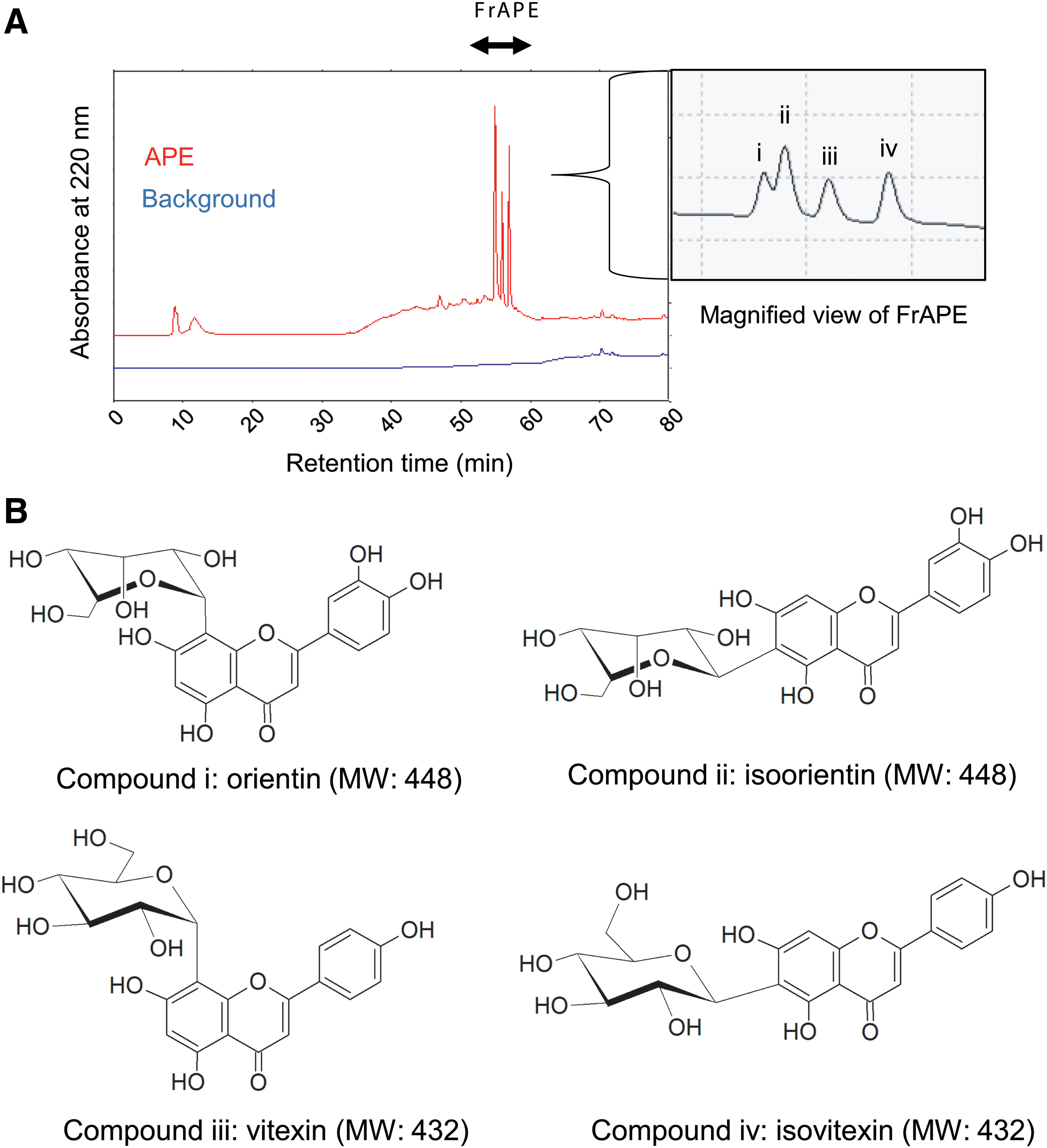
HPLC chromatogram of APE diluted in ultrapure water.
To investigate the effect of the fraction (FrAPE), APE was fractionated by HPLC, as shown in Figure 5A. The dry weight of the fraction collected from 5 mg of APE was 0.483 mg (9.7%). FrAPE and a mixture of commercial products of flavonoids were used for further experiments.
Effect of fractionated APE on osteoclast and osteoblast differentiation
FrAPE was added to RAW264.7 cells with a differentiation-inducing reagent, as described in the experiments above. Counting the TRAP-positive multinucleated cells after TRAP staining indicated that FrAPE suppressed their differentiation into multinucleated osteoclasts (Fig. 6A, B). RAW264.7 cells treated with 10 μg/mL of FrAPE showed significantly decreased osteoclastogenesis. A mixture of flavonoids was added to RAW264.7 cells at a concentration of 5 μM for each flavonoid. TRAP-positive multinucleated cells were counted, after TRAP staining, and indicated that the mixture of flavonoids decreased osteoclastogenesis (Fig. 6C).

Cellular effect of fractionated APE and mixture of flavonoids.
Finally, the activity of FrAPE and the mixture to facilitate osteoblastogenesis was measured as for the experiments above. FrAPE was added to MC3T3-E1 cells with differentiation-inducing reagent. The quantification of calcium deposition after alizarin red staining indicated that 1–10 μg/mL of FrAPE facilitated osteoblast differentiation (Fig. 6D, E). Then, MC3T3-E1 cells were cultured with 1–10 μg/mL of FrAPE for 7 days to investigate ALP activity. Treatment with FrAPE tended to increase ALP activity compared with vehicle control (Fig. 6F). In addition, a mixture of flavonoids was added to cells at concentration of 5 μM for each flavonoid for ALP activity assay and ALP staining. Treatment with the mixture tended to increase ALP activity in MC3T3-E1 cells (Fig. 6G), and similar results were obtained by ALP staining (Fig. 6H).
Discussion
Osteoporosis is a systemic skeletal disorder with various symptoms caused by bone loss. 8,9 Previous reports have shown that foods containing antiosteoporotic compounds can prevent symptoms from progressing in osteoporotic animal models. 19 –21 Recently, antiosteoporotic diets have attracted increased attention suggesting that they are desirable for preventing osteoporosis. 22,23 In this study, we prepared tea leaves and its extract APE from A. palmatum leaves. We found that APE suppressed osteoclastogenesis and facilitated the osteoblastogenesis of cultured cells. The components of APE, which had antiosteoporotic effects on RAW264.7 and MC3T3-E1 cells, were analyzed and separated by HPLC. These processes were similar to those of previous reports of green tea extract-treated cells. 13,24
A previous report identified vitexin (MW = 432), isovitexin (MW = 432), orientin (MW = 448), and isoorientin (MW = 448) in A. palmatum leaves. 16 The four compounds i-iv were detected in APE, and we determined that compounds i-iv corresponded to orientin, isoorientin, vitexin, and isovitexin, respectively. Among these flavonoids, orientin was reported to facilitate osteoblastogenesis through the Wnt pathway. 25 Furthermore, orientin has various medicinal properties, including antioxidant, 26,27 antiaging, 27 anti-inflammatory, 28 antiadipogenesis, 29 and neuroprotective effects. 30 Previously, vitexin was isolated from leaves, and its antioxidant effects were investigated. 31 Similar to orientin, vitexin showed multiple health-beneficial activities. 32 These flavones have received increasing attention because of their wide range of pharmacological effects and might be potential substitute medicines for disease therapy. Therefore, A. palmatum hot water extract has potential health-promoting effects, and we confirmed the effects of APE on osteoclastogenesis and osteoblastogenesis.
Cultivated leaves were processed to afford APE according to the protocol shown in Figure 1. APE-treated RAW264.7 cells showed decreased TRAP-positive multinucleated cells during osteoclast differentiation, while APE-treated MC3T3-E1 cells showed increased calcium deposition and ALP activity during osteoblast differentiation. Candidate effective compounds were determined by HPLC, as indicated in Figure 5A, followed by fractionation. FrAPE and the mixture of four flavonoids decreased osteoclastogenesis in RAW264.7 cells and increased calcium deposition in MC3T3-E1 cells. Although APE and FrAPE significantly suppressed osteoclastogenesis, the effect of individual flavonoids was small. Two compounds with similar structures, vitexin and isovitexin, showed relatively large effects on osteoclastogenesis at 5 μM. The mixture of four flavonoids decreased osteoclastogenesis, and we expect that vitexin and isovitexin are the main active components. In MC3T3-E1 cells, although individual flavonoids showed no notable effect on ALP activity (data not shown), the mixture slightly increased activity. It is possible that multiple compounds can enhance their activities against osteoporosis in a cooperative manner in APE. From HPLC analysis, there were no significant peaks in APE except for the four flavonoids, and no other effective compounds were detected. Overall, these results show that APE suppressed osteoclastogenesis and facilitated osteoblastogenesis, and the active compounds were identified.
In summary, we prepared tea leaves from A. palmatum and investigated the effect and bioactive components of the extract. Treatment with a hot water extract from A. palmatum tea leaves decreased TRAP-positive multinucleated osteoclasts in RAW264.7 cells and increased calcium deposition in MC3T3-E1 cells. HPLC and LC-MS analysis showed the presence of antiosteoporotic components in APE, and the four major peaks were identified as orientin, isoorientin, vitexin, and isovitexin, respectively. This is the first report of an extract from A. palmatum inhibiting osteoclastogenesis and facilitating osteoblastogenesis. In conclusion, our results support the advantage of drinking tea made from A. palmatum leaves to promote bone formation for maintaining bone health. Further studies are required to identify the pathway effected by the components in APE and evaluate the antiosteoporotic effects of APE in animal models.
Footnotes
Acknowledgments
This work was supported partly by a Local Creation Accelerated Grant from the cabinet office, government of Japan (to M. N.). The authors thank Mr. Hibino and Ms. Umetani, Ph. (Mie prefecture Industrial Research Institute) for grinding A. palmatum tea leaves by a jet mill.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.