
Abstract
Nonalcoholic fatty liver disease is a progressive disease involving the accumulation of lipid droplets in the liver. In this study, we investigated the anti-hepatosteatosis effects of fermented Cordyceps militaris extract (CME) in AML-12 hepatocytes. Although the levels of adenosine and cordycepin were reduced in the extracts of CM grown on germinated soybean (GSCE) and fermented CM grown on germinated soybean (GSC) by Pediococcus pentosaceus ON188 (ON188E), the expression of fatty acid oxidation (FAO) genes were upregulated only by GSC-ON188E treatment in a dose-dependent manner. In contrast, a lipogenic gene, stearoyl Coenzyme A desaturase 1, was downregulated by ON188E. Formation of intracellular lipid droplets by the addition of oleic acid was reduced by ON188E to levels observed in WY14643-treated cells. When cells were treated with ON188E, sphingosine kinase 2 mainly responsible for hepatic sphingosine 1-phosphate (S1P) synthesis was upregulated and S1P was elevated. Collectively, the fermented GSC extract activates FAO through elevation of S1P synthesis and has potential as a therapeutic for hepatosteatosis.
Introduction
Nonalcoholic fatty liver disease (NAFLD) represents a group of liver diseases that begin with the simple accumulation of hepatic triglycerides (TG), called hepatosteatosis. NAFLD has a large disease spectrum from simple steatosis to hepatocellular carcinoma, through to nonalcoholic steatohepatitis (NASH), fibrosis, and cirrhosis. 1 The first stage is hepatosteatosis caused by accelerated de novo lipogenesis, inhibited triacylglycerol (TG) release, and increased fatty acid (FA) influx from adipose tissue by circulation. 2 Impaired fatty acid oxidation (FAO) and abnormal FA uptake, together with increased de novo lipogenesis in the liver, contribute to the development of hepatosteatosis, followed by an inflammatory stage called NASH. 3,4 Characterized by the presence of hepatocyte injury, an inflammatory infiltrate, and collagen deposition, NASH can lead to the development of cirrhosis and hepatocellular carcinoma. 5
FAO is a process catabolizing FA into acetyl-Coenzyme A (CoA) in the mitochondria and peroxisomes, by which citric acid cycle and electron transport chain were initiated to maintain energy homeostasis. Certain molecular markers have been studied for their indispensible roles in FAO regulation such as peroxisome proliferator-activated receptor alpha (PPARα), carnitine palmitoyltransferase 1α (CPT1α), and peroxisomal acyl-coenzyme A oxidase 1 (ACOX1). As a primary rate-controlling enzyme, CPT1α responded for the translocation of long-chain fatty acids (LCFAs) into mitochondria in FAO. 6,7 Similarly, the oxidation of LCFAs is initiated by ACOX1 in peroxisomal FAO system. 8 In addition, although PPARα and its target genes are well-known as the FA metabolic regulators, 9,10 stearoyl CoA desaturase 1 (SCD1) catalyses the synthesis of monounsaturated FAs that can later form TG for energy storage. 11,12 Because impairment of FAO and FA uptake results in development of hepatosteatosis, the research into the development of therapeutic interventions against hepatosteatosis has been actively on-going by modulation of hepatic FAO.
The therapeutic effects of traditional medicines and natural products have been drawing attention for several decades. Among them, Cordyceps militaris (CM) (Family: Clavicipitaceae), a traditional medicinal mushroom, has been used extensively as a crude drug and functional food in East Asia. 13 CM is not only known for its antioxidant, anti-allergy, anti-asthma, and anticancer activities, 14,15 recent studies have provided evidence that CM extracts (CMEs) also have strong anti-inflammatory and anticancer activities. 16,17 The protective effects of CM on mitochondria, testosterone stimulation, and aging have also been demonstrated, suggesting that CM is an efficacious medicine for eternal youth. 18,19
Furthermore, CM is known to contain numerous components that possess anti-inflammatory and immune-modulating activities. 20 A major component of CM, adenosine, and its receptors has been demonstrated to have important roles in regulating hepatosteatosis and atherosclerosis. 21,22 Among adenosine derivatives, cordycepin is the most widely studied bioactive compound that regulates lipid metabolism through AMP-activated protein kinase in HepG2 cells, 23 inflammatory response through the nuclear factor NF-κB pathway in macrophages, 24 and altering carcinoma progression of colonic cells. 25 These two compounds were investigated together as phenotypic switches of macrophages in inflammatory diseases. 26
Moreover, the beneficial effects of CM supplementation on the development of fatty liver, oxidative stress, and inflammatory cytokine levels have been demonstrated previously in ob/ob mice. 27 However, the preventive effects of CM were not profound in terms of fatty liver. In addition, the mechanisms of CM supplementation and its therapeutic targets have not been elucidated. A recent report showed that adenoviral overexpression of hepatic sphingosine kinase 2 (SPHK2), a major sphingosine kinase isoform in the liver, activates FAO and improves hepatosteatosis conditions in a diet-induced obese mouse model. 4 Therefore, the involvement of sphingosine 1-phosphate (S1P), the product of SPHKs, may be important in the alleviation of hepatosteatosis.
In this study, we investigated the efficacy of various CM products obtained using different growth conditions and fermentation procedures on FAO in hepatocytes, and the involvement of sphingolipid metabolism in the prevention of hepatosteatosis.
Materials and Methods
Preparation of CME and CM grown on germinated soybean extract
An authenticated voucher specimen of CM (Kucari 0906) was obtained from the Herbarium at the College of Bioscience and Biotechnology, Konkuk University (Seoul, Korea). The mycelium of CM was inoculated onto germinated soybeans (Rhynchosia volubilis Lour) and was cultured at 20–25°C for 4 weeks (GSC; Kucari 0903). Dried CME and the extract of CM grown on germinated soybean (GSCE) were ground to a powder and extracted with boiling water for 2 hours. For fermentation, Pediococcus pentosaceus ON188 was inoculated in the GSC water extract and incubated at 37°C for 48 hours. The supernatant extracts were filtered again and dried using a rotary evaporator under vacuum at 40°C then freeze-dried. The powder (ON188 extract, ON188E) was stored at −20°C until the experiment.
Chemical analysis of CME and GSCE using gas chromatography time of flight mass spectrophotometry
Adenosine and cordycepin were purchased from Sigma-Aldrich (St. Louis, MO, USA). Both compounds were dissolved in distilled water to a concentration 10 μg/mL. We then aliquoted 50 μL of each compound into 1.5 mL microtubes and concentrated them to complete dryness in a speed vacuum concentrator (SCANVAC, Korea). The freeze-dried samples (CM, GSC, and ON188) were reconstituted with 1 mL of the solvents initially used, diluted to a concentration of 500 μg/mL for cell treatment.
Three types of samples were prepared for the standard compounds as follows: reagent blank, standard compounds, and standard compounds spiked with the extracts. We added 5 μL of 40 mg/mL methoxyamine hydrochloride (Sigma-Aldrich) in pyridine (Thermo, USA) to the dried samples and the standards were incubated for 90 minutes at 30°C. We then added 2 μL of the mixture, consisting of 13 FA methyl esters, as an internal retention index marker, and 45 μL of N-methyl-N-trimethylsilyltrifluoroacetamide (MSTFA +1% TMCS; Thermo) was added for trimethylsilylation (1 hour at 350 g and 37°C). 28
A 0.5 μL aliquot of derivatized mixture was injected using an Agilent 7693 ALS (Agilent Technologies, Wilmington, DE, USA) in splitless mode, into an Agilent 7890B gas chromatograph (Agilent Technologies) for chromatographic separation (30 m long, 0.25 mm id. Rtx-5Sil MS column with 0.25 m 95% dimethyl 5% diphenyl polysiloxane film), using an additional 10 m integrated guard column (Restek, Bellefonte, PA, USA). The column temperature was held constant at 50°C for 1 minute, then increased at 20°C/min to 330°C, and maintained for 5 minutes. 29
Mass spectrometric analysis was conducted on a LECO Pegasus HT time-of-flight (TOF) mass spectrometer, controlled by the LECO ChromaTOF software 4.50 version (LECO, St. Joseph, MI, USA). Transfer line and ion source temperature were set to 280°C and 250°C, respectively. Mass spectra were collected from 85 to 500 m/z at 17 spectra per second of acquisition rate and 1800 V detector voltage. Data preprocessing was conducted using ChromaTOF software and upon data acquisition, the apex mass values, the entire spectrum, retention time, peak purity, and signal-to-noise ratio were acquired. 30
AML-12 cell culture
Murine hepatocytes (AML-12) were obtained from the American Type Culture Collection (Manassas, VA, USA) and cultured in Dulbecco's modified Eagle's medium (DMEM)/F12 (Welgene, Inc., Korea) supplemented with 10% heat-inactivated (56°C, 30 minutes) fetal bovine serum (Welgene), 10 U/mL penicillin, and 10 μg/mL streptomycin (Gibco, Life Technology, CA, USA), and 40 nM dexamethasone (Sigma-Aldrich). Cells were maintained at 37°C in a 5% CO2-supplied incubator. For treatment of natural extracts, AML-12 cells were seeded at 5 × 10 5 cells per well in six-well culture plates. When the cells reached 70–80% confluence, they were incubated in serum-free DMEM/F12 for 4 hours and then treated with natural extracts for 24 hours.
RNA preparation and real-time quantitative polymerase chain reaction
Total RNA was extracted from AML-12 cells using an easy-spin Total mRNA Extraction Kit (Intron Biotechnology, Inc., Korea) according to the manufacturer's instructions. Complementary DNA was synthesized using an iScript cDNA synthesis Kit (Bio-Rad, Hercules, CA, USA) in a polymerase chain reaction (PCR) thermal cycler (TaKaRa, Japan). Real-time PCR analysis was performed using a SYBR Green Premix (TaKaRa) in a Step-One Plus real-time PCR system (Applied Biosystems, Carlsbad, CA, USA). The subsequent mRNA level was normalized to mouse GAPDH. Our study used the following primer sequences: GAPDH, forward-CAA GGT CAT CCA TGA CAA CTT TG, reverse-GGC CAT CCA CAG TCT GG; SPHK2, forward-AGA CGG GCT TTA CGA G, reverse-CCT GCT CAA ACC CGC CAT; FAS, forward-GGT CGT TTC TCC ATT AAA TTC TCA T, reverse-CTA GAA ACT TTC CCA GAA ATG TTC C; SCD1, forward-TTC TTG CGA TAC ACT CTG GTG C, reverse-CGG GAT TGA ATG TTC TTG TCG T; CPT1α, forward-CTT CCA AGG CAG AAG AGT GGG, reverse-GAA CCT TGG CTG CGG TAA GAC; PPARα, forward-ATC CAC GAA GCC TAC C, reverse-CAC ACC GTA CTT TAG CAA G; ACOX1, forward-ACG CCA CTT CCT TGC TCT TC, reverse-AGA TTG GTA GAA ATT GCT GCA AA. Statistical significance was calculated using a Student's t-test with Prism software (GraphPad, CA, USA).
Protein assay and western blot analysis
AML-12 cells were grown at 1 × 10 6 cell per plate (60 mm) for 24 hours and incubated with various doses of CME and GSCE after 4 hours incubation in no-serum media. Cells were harvested and homogenized in 200 μL lysis buffer (10 mM Tris pH 7.8, 1 mM ethylenediaminetetraacetic acid, 150 mM NaCl, phosphatase inhibitor, and protease inhibitor) by vortexing. Lysate (supernatant) was collected for protein analysis using a Protein Assay Kit (Bio-Rad). SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) was performed with 20 μg of total protein. The proteins were later transferred onto polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA). Rabbit primary antibodies against CPT1α (Cell Signaling Technology, MA, USA), PPARα, ACOX1, and Sphk2 (Abcam, Cambridge, MA, USA) were used to detect target proteins. After incubation with a secondary horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G antibody (Millipore), the blots were developed with an enhanced chemiluminescent substrate (Millipore) and the bands were detected using chemiluminescence imaging equipment (Viber Lourmat, France). Mouse primary antibody against β-actin (Millipore, Cambridge, MA, USA) was used to normalize the samples.
Cell viability
Cells were grown in 2 × 100 mm plates until 90% confluence and viability was examined using the Trypan Blue staining method. Viable cells were seeded in 96-well plates at a density of 2 × 10 4 cells, in 200 μL complete DMEM/F12 medium. After 24 hours of cell attachment, the cells were exposed to various doses of GSCEs and incubated for 24 or 48 hours at 37°C. At the end of the incubation period, the mitochondrial function was assessed by cell proliferation assay kit (Welgene) using 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT) for 4 hours at 37°C and quantified at 540 nm in a microplate spectrophotometer (Epoch 2; Biotek, USA).
FAO measurement
AML-12 cells were grown and WY14643 and ON188E were treated at various concentrations for 24 hours. Cellular FAO was measured by FAO assay kit (Abcam) according to the manufacturer's instructions. Fluorescence signal was measured by Enspire plate reader (PerkinElmer, Waltham, MA, USA).
Sphingolipids analysis
For analyses of sphingolipids, AML-12 cells were harvested and lysed in phosphate-buffered saline. A known amount of C17:0–ceramide (internal standard) was added to the cellular extracts containing 1 mg of protein. Sphingolipids were extracted using chloroform/methanol (2:1, v/v) containing 0.01% butylated hydroxytoluene. KOH was added to the mixture to saponify the sphingolipids at 37°C for 2 hours. The lipid extracts were neutralized by adding acetic acid, and the organic phase was separated and dried under nitrogen gas. Sphinganine, sphinganine 1-phosphate (Sa1P), sphingosine, and S1P were separated by high-performance liquid chromatography using a C18 column (XTerra C18, 3.5 μm, 2.1 × 50 mm) and ionized in positive electrospray ionization mode, as described by Yoo et al. 31 [M+]/product ions from corresponding sphingolipid metabolites were applied for multiple reaction monitoring quantification by a bench-top tandem mass spectrometer, API 4000 Q-trap (Applied Biosystems, Framingham, MA, USA), interfaced with an electrospray ionization source.
Oil Red O staining
After treatment with GSCEs for 24 hours, cells were washed and incubated with 10% formalin for 1 hour at room temperature. Cells were then washed with 60% isopropanol and dried completely, followed by incubation with Oil Red O working solution for 10 minutes. Samples were then washed thoroughly several times with distilled water and examined by microscopy (Leica Microsystems, Korea). Lipid contents were quantified by isopropanol extraction of Oil Red O-stained cells and measurement of optical density using Epoch reader (Biotek, USA).
Statistical analyses
Results are expressed as mean ± standard error of the mean. Three independent experiments were performed in one group (n = 3). Student's t-test was used for assessing statistical significance between control and sample-treated groups, represented by P-values. Results were considered to be statistically significant when P < .05.
Results
Extracts of CM, GSC, and GSC-ON188 contain cordycepin and adenosine
As cordycepin and adenosine have been studied for their anti-inflammatory, anticancer, and anti-hepatosteatosis activities, 21,24 –26 we examined the levels of these compounds in CME, GSCE, and GSC-ON188E. To identify the bioactive components in CME and GSCE, we analyzed the extracts using gas chromatography time-of-flight mass spectrometry (Supplementary Fig. S1). Total ion chromatograms of GSCE and ON188E are given in Figure 1A and B. The two major peaks of cordycepin and adenosine of the standard samples in the ion chromatogram (positive ion mode 236 m/z) were eluted at retention times of 832.6 and 851.75 seconds, respectively (data not given). Structure identities of adenosine and cordycepin were observed compared with a library of MS spectra (Supplementary Fig. S1C, D).

ON188E, but not CME or GSCE, upregulates FAO genes. After 4-hour serum starvation, AML-12 cells were co-treated with 200 μM oleic acid, 10 μM WY14643 (WY), and 50 μg/mL of CME, GSCE, and ON188E for 24 hours. The mRNA expression levels of FAO genes including CPT1α
The amounts of the two representative active components in the CME were significantly higher than those in GSCE and ON188E (Table 1). The value of adenosine in CME was 143.5 ± 1.64 μg/g, whereas in GSCE it was 58.2 ± 1.27 μg/g. The content of adenosine in ON188E was 68.9 ± 0.57 μg/g, which was higher than that of GSCE. Similarly, the cordycepin levels contained in CME (2.64 ± 0.02 μg/mL) were higher than those of GSCE (1.26 ± 0.01 μg/g) and ON188E (1.36 ± 0.01 μg/g). These results suggest that the growth conditions and fermentation process affect the contents of representative molecules in CM.
Measurement of Adenosine and Cordycepin in Cordyceps militaris Extract, GSCE, and ON188E by Gas Chromatography/Mass Spectrometry
CME, natural C. militaris extract; GSCE, the extract of Cordyceps militaris grown on germinated soybean; ON188E, GSCE fermented by Pentosaus.
ON188E upregulates expression of FAO genes
To examine the effects of CME, GSCE, and ON188E on hepatic FAO, AML-12 cells were treated with each of these extracts. As oleic acid is suggested to be a hepatic steatosis inducer, 32 cells were pretreated with 200 μM oleic acid for 24 hours, followed by treatment of 50 μg/mL CME, GSCE, or ON188E. Expression of FAO genes such as PPARα, CPT1α, and ACOX1 were determined by mRNA measurements. We observed an induction of PPARα and CPT1α mRNA in cells incubated with ON188E, whereas there was no change in the CME- or GSCE-treated cells in comparison with the control (Fig. 1A, B). In contrast, cells exposed to WY14643, a PPARα agonist, showed a modest increase in CPT1α and PPARα expression (Fig. 1A, B). Besides, ON188E slightly elevated ACOX1, whereas CME that downregulated ACOX1 and GSCE did not cause any changes in the expression of ACOX1 or SPHK2 (Fig. 1C, D). These results suggest that ON188E induces the expression of FAO genes and activates FAO in hepatocytes.
ON188E has no effect on AML-12 cell viability yet inhibits lipogenesis
Based on the results of ON188E-mediated induction of FA oxidative gene expression, we decided to characterize the effects of ON188E on hepatic FAO. Cell viability was not altered when AML-12 cells were treated with 50 μg/mL of ON188E for 24 or 48 hours (Fig. 2A, B). To investigate whether ON188E can alter lipogenesis, the expression of fatty acid synthase (FAS) and SCD1 mRNA was measured. Although FAS expression was not altered, SCD1 was downregulated by 25% after ON188E treatment (Fig. 2C, D). In contrast, WY14643 treatment led to upregulation of FAS by 48% and SCD1 by 31%. These results suggest that FA synthesis is inhibited by ON188E at the SCD1 step.
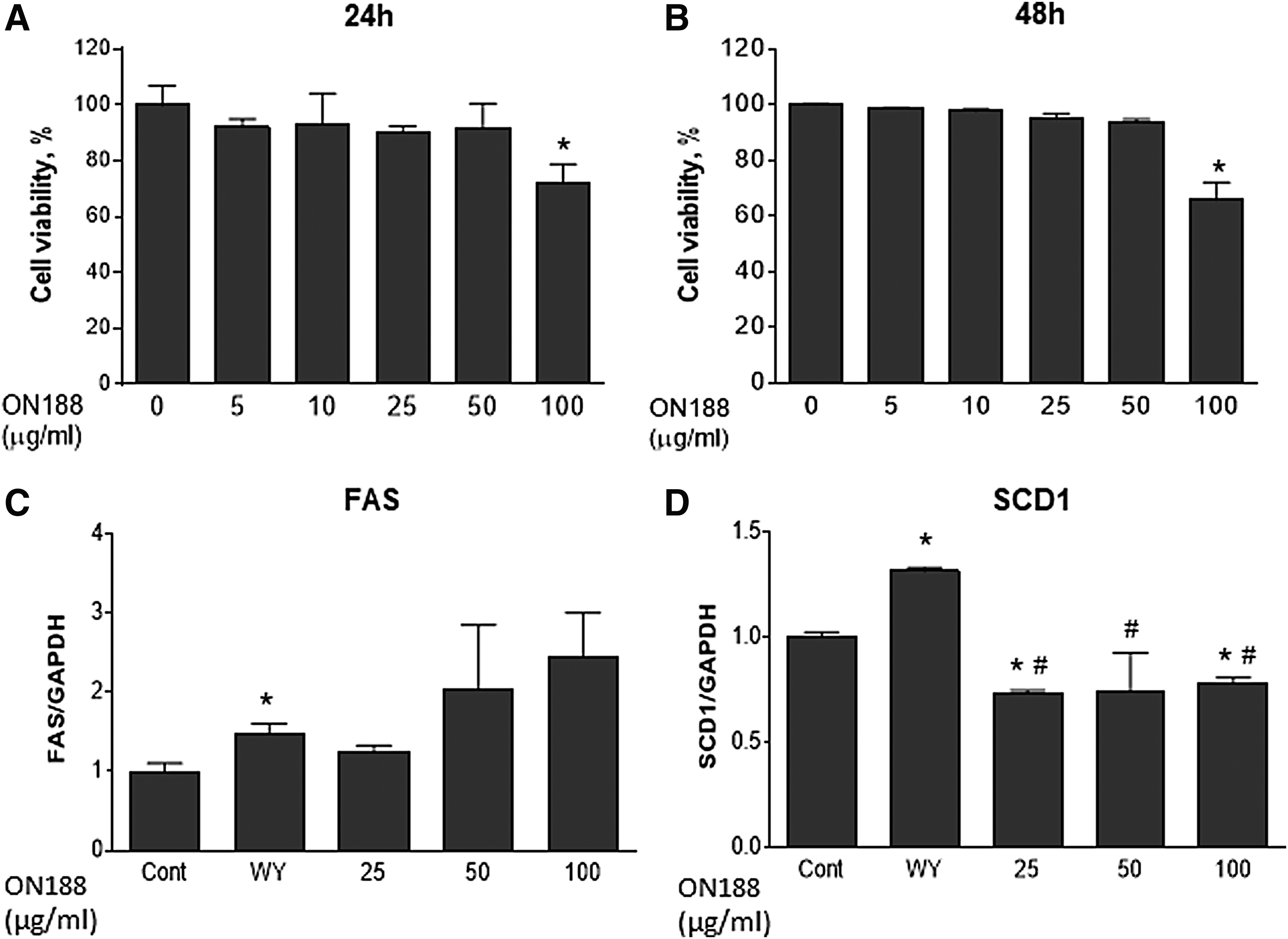
Cytotoxicity of ON188E in AML-12 cells and the expression of TG biosynthetic genes. AML-12 cells were treated with various concentrations of ON188E and incubated for 24 hours
ON188E induces the expression of FAO genes and reduces lipid droplets in cells
To examine the effects of ON188E on FAO under no steatosis and normal conditions, AML-12 cells were treated with ON188E without oleic acid for 24 hours. The expression of FAO genes was then analyzed to determine mRNA and protein levels at various concentrations of ON188E. ON188E treatment upregulated CPT1α and PPARα expression in a dose-dependent manner by 3.9- and 10.4-fold, respectively, compared with those of untreated controls (Fig. 3A, B), whereas modest downregulation was observed for ACOX1 expression at 25 μg/mL of GSC-ON188 but no change at higher concentration (Fig. 4C). In contrast, WY14643 upregulated CPT1α, PPARα, and ACOX1, although not to the levels of the ON188E-treated cells (Fig. 3A–C). Consistent with the mRNA expression, the protein levels of CPT1α, PPARα, and ACOX1 were all elevated by ON188E (Fig. 3D). To confirm whether altered expression is reflected in FAO activation, we measured the oxygen consumption representing FAO. WY14643-treated cells showed increased oxygen respiration compared with those of the no-treatment control and ON188E-treated cells. ON188E treatment activated oxygen respiration in a dose-dependent manner but not to the levels of WY14643-treated cells (Fig. 3E).
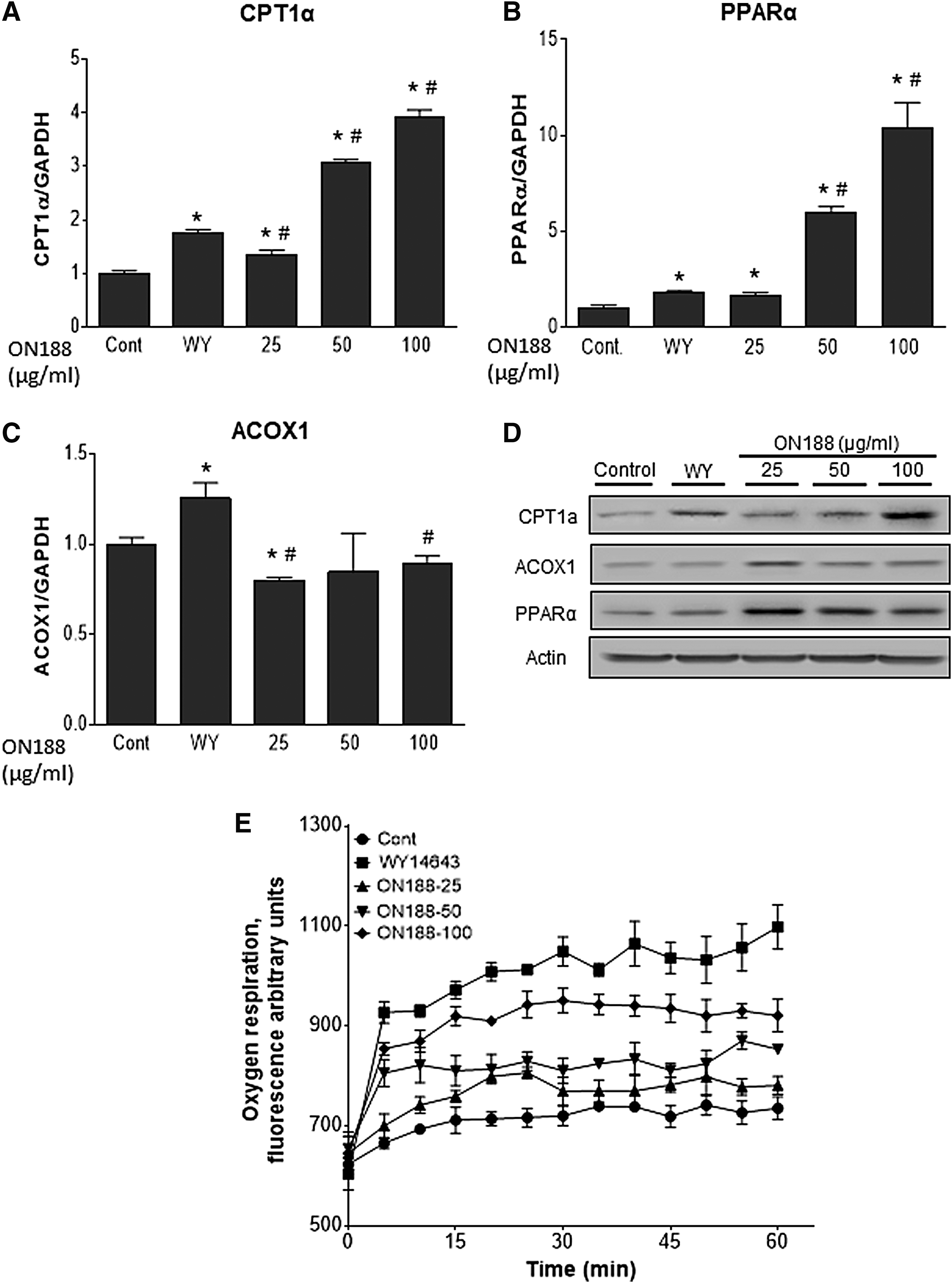
ON188E upregulates FAO genes. AML-12 cells were treated with 10 μM WY14643 or various concentrations of ON188E for 24 hours. Expression of CPT1α
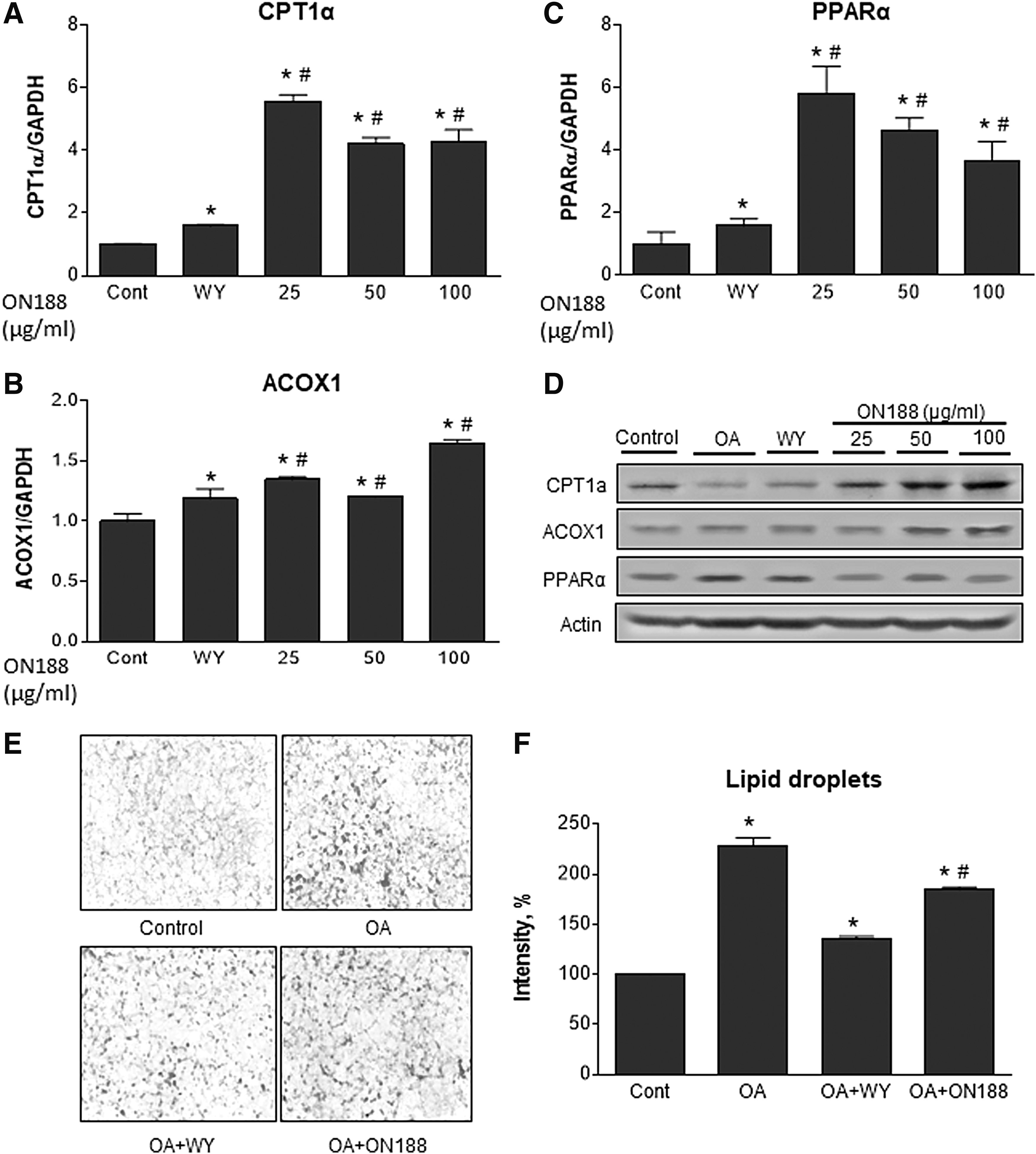
ON188E upregulates FAO genes and reduces cellular lipid droplets in the presence of oleic acid. AML-12 cells were treated with 10 μM WY14643 or various concentrations of ON188E in the presence of 200 μM oleic acid for 24 hours. Expression of CPT1α
In addition, AML-12 cells were pretreated with 200 μM oleic acid for 24 hours to mimic hepatosteatosis conditions and then incubated with ON188E for an additional 24 hours. Expression of CPT1α, PPARα, and ACOX1 mRNA were all upregulated when cells were treated with ON188E (Fig. 4A–C). Again, WY14643 slightly upregulated these genes. Immunoblot analyses demonstrated that CPT1α and ACOX1 proteins were elevated, but PPARα was not altered by ON188E (Fig. 4D). To determine whether ON188E reduces lipid accumulation in liver cells, Oil Red O staining was performed on the cells after preincubation of oleic acid followed by WY14643 or ON188E treatment. Compared with the oleic acid-treated cells, the degree of Oil Red O staining was significantly reduced by ON188E, which did not reach the levels of the WY14643-treated cells (Fig. 4E, F). These results suggest that ON188E upregulates FAO genes and activates FAO under normal and hepatosteatosis conditions.
ON188E upregulates SPHK2 and elevates sphingoid bases
Our previous study demonstrated that the adenoviral overexpression of SPHK2 activates FAO by upregulation of FA oxidative genes in the liver. 4 To examine the involvement of SPHK2 and S1P in activation of FAO, we investigated whether ON188E regulates expression of SPHK2 and cellular S1P. In the absence of oleic acid, ON188E upregulated SPHK2 in a dose-dependent manner at a maximum of 5.1-fold, whereas a slight increase was observed after WY14643 treatment (Fig. 5A, C). In contrast, the presence of oleic acid upregulated SPHK2 mRNA, but the degree of SPHK2 upregulation was less than that in the no-oleic acid treatment group. However, SPHK2 upregulation was clearly observed in the protein levels (Fig. 5B, D). To examine whether SPHK2 upregulation by ON188 leads to the elevation of cellular S1P or Sa1P, we measured the cellular levels of sphingoid bases by LC-MS/MS. Sphingosine and sphinganine, the precursors for S1P or Sa1P, were elevated twofold (Fig. 5E, F). S1P and Sa1P, the SPHK2 enzyme products, were also elevated twofold (Fig. 5G, H). These results suggest that the activation of FAO is partly mediated by an SPHK2-mediated increase in S1P and Sa1P.
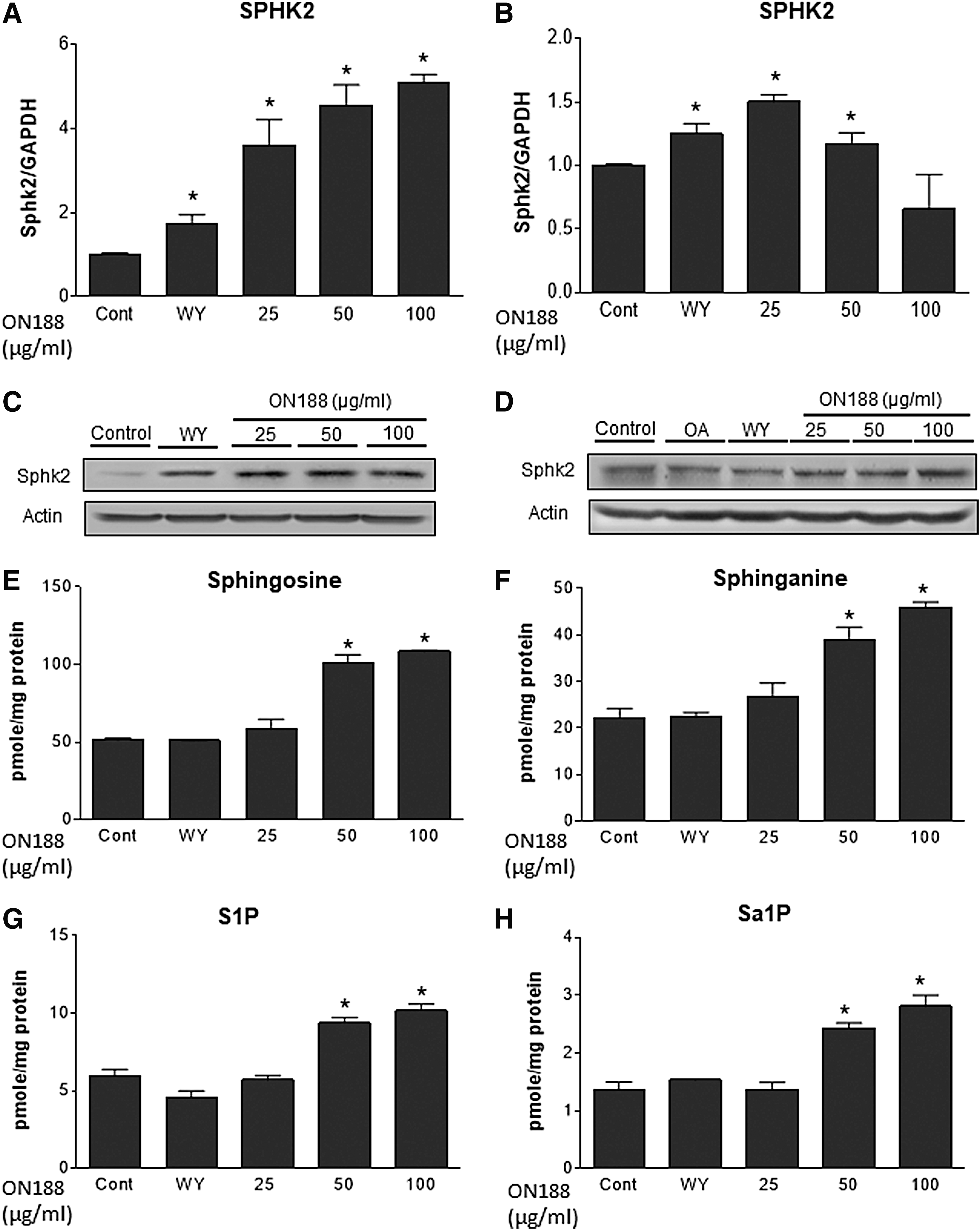
SPHK2 was upregulated by ON188E. AML-12 cells were treated with 10 μM WY14643 or various concentrations of ON188E in the absence
Discussion
Hepatosteatosis is the early stage of NAFLD, characterized by the accumulation of ectopic fat in the liver, although it presents no clinical symptoms. Ectopic fat accumulation in the liver has a positive correlation with oxidative stress, mitochondrial dysfunction, production of proinflammatory cytokines, and apoptosis. 33 As a result, NASH can progress to an advanced stage of NAFLD, which can progress to cirrhosis and liver cancer. 34 In this study, we found the following: (1) ON188E but not CME or GSCE upregulates the expression of FA oxidative genes under normal and hypertriglycemic conditions, (2) ON188E suppresses the expression of lipogenic genes and inhibits TG biosynthesis, (3) ON188E upregulates SPHK2 and elevates S1P and Sa1P, which activate FAO in hepatocytes.
CM has been commonly used as a traditional medicine in East Asia for treating inflammation, anemia, and cancer. 13 One of the major components in CM is adenosine, which possesses anti-hepatosteatosis activities through adenosine receptor-mediated signaling. 35 However, the mechanism of this hepatic TG-lowering mechanism remains unknown. As CM is limited in nature, we developed a method to cultivate CM in germinated soybeans (R. volubilis Lour) (GSC) and then compared it with natural CM and GSC fermented by P. pentosaceus ON188. We found that the expression of FAO genes in hepatocytes was upregulated only by ON188E. Of interest, upregulation of FAO genes was found regardless of the addition of oleic acid to mimic hyperlipidemic conditions. Supplied FA could activate the extracellular FA uptake and transportation system that activates intracellular TG biosynthesis leading to steatosis. This is a similar disease status to hepatosteatosis in vivo. As obesity-induced spillage of free FA from adipocytes is one of the major cause of hepatosteatosis, 36 the use of ON188E can be a therapeutic intervention method for prevention of further development of chronic liver diseases.
The fact that GSCE and ON188E had lower levels of adenosine and cordycepin, the representative bioactive compounds in CM, suggests that the anti-hepatosteatosis effect of ON188E is not because of adenosine or cordycepin. As ON188E is produced from CM cultured in germinated soybean and then fermented by P. pentosaceus ON188, ON188E contains the bioactive secondary metabolites from soybean and from the probiotic bacteria. Various reports have demonstrated that soybean sprouts are rich sources of vitamins and isoflavones for antioxidant, antidiabetes, and anti-inflammatory activities. 37,38 Besides, P. pentosaceus is a lactic acid bacteria strain that was found abundant in breast milk and having high antifungal activities. 39 This lactic acid bacteria strain is known to contain γ-aminobutyric acid, an immunologically active compound and neurotransmitter. 40 Moreover, the degree of FAO upregulation by ON188E was comparable with those observed with treatment with WY14643, a PPARα agonist. However, it is unclear which of the bioactive secondary metabolites possibly from GSC or ON188 lactic acid bacteria activate FAO at this moment and further studies will be required.
Hepatic lipids are maintained by the balance between FA/TG biosynthesis and FAO. We found that SCD1 was downregulated by ON188E, whereas no change was found in the expression of FAS and diglyceride acyltransferase 2 (data not given). As SCD1 is an important step in TG biosynthesis and catalyses the formation of stearoyl CoA for TG biosynthesis, it is regarded as a therapeutic target for hypertriglyceridemia and metabolic disease. Previous reports have revealed that mice deficient in SCD1 showed increased FAO 41 and were protected from high-carbohydrate diet-induced obesity and hepatic steatosis. 42 This result suggests that hepatocytes exposed to ON188E not only have higher FAO, but also inhibit FA/TG biosynthesis. In contrast, treatment with WY14643 compound upregulated SCD1 mRNA expression as observed previously. 43 –45 PPARα agonists such as WY14643 and fenofibrate activate not only FAO but FA/TG biosynthesis by upregulation of FA/TG biosynthetic genes including SCD1, acetyl-CoA carboxylase (ACC), fatty acid synthase (FAS), and glycerol phosphate acyltransferase. 45 Although PPARα agonists are prescribed as a substitute medicine for hepatosteatosis, a balance between beneficial activation of FAO and aggravating FA/TG synthesis should be considered carefully for intervention.
PPARα is a ligand-activated transcription factor regulating energy homeostasis. 46 As PPARα agonists have been proved to lower plasma TG and raise plasma high-density lipoprotein levels, 47,48 we used WY14643, as a positive control. It is interesting to note that the upregulation of FAO genes caused by ON188E was even higher than those of WY14643. These results imply that ON188E could have positive effects for the prevention and treatment for hepatosteatosis. In addition, our cell studies require further confirmation in animal and clinical models.
Sphingolipid biosynthesis has been regarded as an important regulator of metabolic and cardiovascular diseases. 49,50 The roles of bioactive sphingolipid metabolites including ceramide, sphingosine, and S1P have been suggested to be essential signaling messengers of metabolic regulation. 51 –53 Among them, S1P is a bioactive lipid mediator associated with injury response, migration, cell proliferation, and interaction. 54,55 Previously, we found that adenoviral overexpression of SPHK2 in the liver led to upregulation of PPARα, CPT1α, and ACOX1, and activated hepatic FAO through S1P elevation. 4 In this study, we found that SPHK2 was upregulated by ON188E treatment. This effect was more evident when cells were treated with ON188E in the absence of oleic acid. Consistent with the expression results, intracellular S1P and Sa1P were also elevated. As S1P is a ligand for PPAR, 56 we expected that the lower degree of SPHK2 upregulation by oleic acid would result in suppressed expression of FAO genes. However, we found that the degree of upregulation of FA oxidative genes in the presence of oleic acid was largely unaltered compared with that of cells not treated with oleic acid (Figs. 4 and 5). These results suggest that S1P-dependent activation of PPAR partly contribute to FAO. S1P-independent activation of FAO and the active components for these effects deserve further studies.
In conclusion, fermented ON188E activates FAO and suppresses TG biosynthesis through S1P-dependent PPARα activation and an S1P-independent pathway (Fig. 6). In addition, S1P is elevated by ON188E-mediated SPHK2 induction in hepatocytes. The fact that ON188E upregulates FAO genes regardless of oleic acid addition, suggests its potential as a pharmacological agent for the prevention and treatment of hepatosteatosis. Therefore, the anti-hepatosteatosis effects of ON188E can be applied to the development of health-promoting beverages or functional foods.
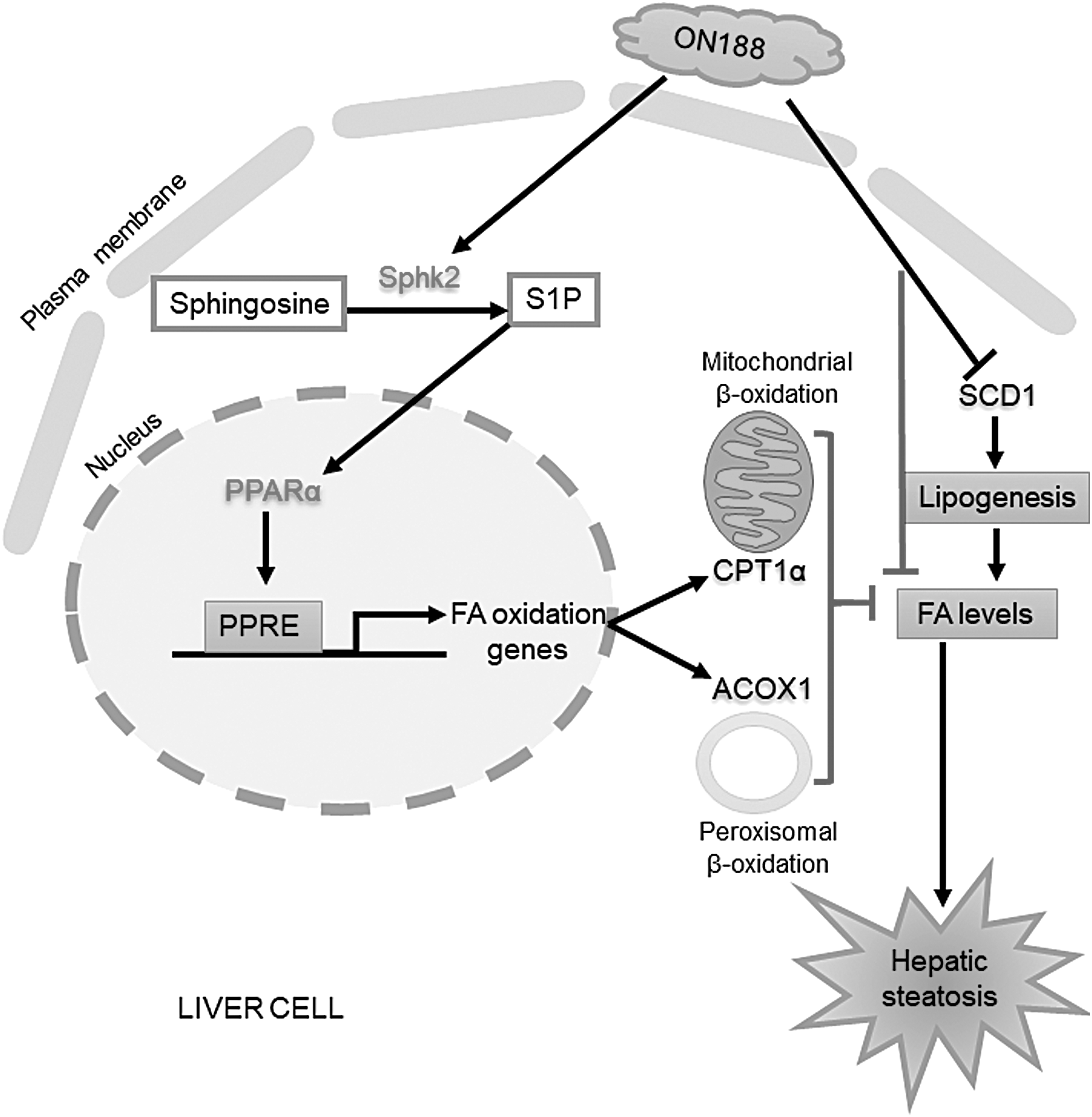
Schematic diagram for the mechanism of ON188E in the activation of FAO and the reduction of lipid droplets.
Footnotes
Acknowledgments
This study was funded by the Agricultural Microbiomes R&D program in Agriculture and Food, Ministry of Agriculture, Food and Rural Affairs, Republic of Korea [as part of the (multiministerial) Genome Technology to Business Translation Program] (916008-02-1-SB010 to T.-S.P.) and the Bio & Medical Technology Development Program through the National Research Foundation of Korea (NRF) (NRF-2014M3A9B6069338 to T.-S.P.).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.